缺磷对黑木相思生长、营养和叶绿素荧光的影响
2021-11-06裘珍飞曾炳山范春节
裘珍飞,曾炳山,范春节
(中国林业科学研究院 热带林业研究所,广东 广州 510520)
黑木相思Acacia melanoxylon原产于澳大利亚南部,是相思类高大乔木,具有固氮、速生、耐干旱、耐瘠薄等特点。20世纪90年代引进我国后,经过30 多年的培育和优良无性系的选育,黑木相思其优势在我国华南地区逐渐显现,近年来,黑木相思无性系造林推广面积日益扩大,以超过666.7 hm2/a 的速度发展,已成为速生丰产,珍贵用材和生态公益等多用途树种[1]。由于我国南方土壤普遍呈酸性,有效磷含量低,经营人工林时因缺磷影响生长量[2]。在林业生产上通过加施磷肥提高植物可利用磷水平,但盲目、过量施肥,不仅影响植株生长,同时会造成肥料浪费、环境污染和土壤养分失衡等后果。因此了解黑木相思磷胁迫下养分调节机制对栽培中磷养分的调控具有重要的科学意义。
磷是植物需要的主要营养元素,它直接参与光合作用各个环节,影响氮、钾等营养元素的吸收和利用,最终决定了植物生长发育和产量形成。低磷胁迫影响橡胶树Heveabrasiliensis对于氮、钾的吸收和积累[3]。但低磷环境增加了苹果砧木磷素和氮素的转运和利用效率[4]。磷胁迫导致叶绿素荧光参数发生显著变化,叶片光能利用和光化学效率下降[5-6]。相思树A.rostellifera是耐低磷树种,在氮和磷极度匮乏的立地上,仍能保持较大的生长量,这与叶片中保持一定的氮、磷含量有关[7]。国内开展过黑木相思苗期磷肥研究,但尚未涉及磷素缺乏对氮、钾吸收利用和叶片荧光叶绿素特征影响研究[8]。本研究以黑木相思优良无性系为材料,研究了缺磷下苗木的营养积累、分配和叶片叶绿素荧光参数,以期更加清楚地了解黑木相思在缺磷条件下自身的营养特性和光反应系统的效率,为后续的养分调控研究提供依据。
1 材料与方法
1.1 试验材料
以黑木相思组培苗为材料,选取生长统一的苗木,培养在海绵支撑的水培育苗盘中,育苗盘规格20 cm×30 cm,盘内浇灌营养液300 mL,每盘种植20 株。
1.2 试验方法
设置全P(对照)和缺P 两个处理:全P处理浇灌Hoagland’s 全营养液,缺P 处理浇灌Hoagland’s 营养液中不添加KH2PO4(136 mg/L),但加入KCl(74.5 mg/L)用于平衡K 离子,其他营养液元素组成不变。整个试验在热带林业研究所炼苗温室中进行,温室温度为23~27℃,光照8 000~10 000 Lx。每处理3 盘,共6 盘,培养3个月后测定各项指标。
1.3 指标测定及方法
1.3.1 生长及养分的测定
每盘随机抽取苗木10 株,每处理共30 株,小心清除根部海绵支撑物,测量苗高和茎粗。把植株分成根、茎、叶3 部分,称取鲜质量,收集上述根、茎、叶,在105℃恒温烘干箱中杀青30 min,后在70℃下烘干至质量恒定,分别称量根、茎、叶的干质量。利用烘干的样品,测定根茎叶的全N、全P 和全K 含量,全N 采用凯氏滴定法测定,全P 采用钼锑抗比色法测定,全K 采用火焰原子吸收分光光度法测定。
1.3.2 叶绿素荧光的测定
黑木相思苗期叶片存在2 种形态,即幼叶(羽状复叶)和成年叶(叶状柄),每处理选取平均长势的苗木,采用LI-COR 公司生产的LI-6400 型便携式光合测定仪分别测定4 片幼叶和4 片成年叶。测定前用铝箔纸包裹叶片暗适应,测定初始荧光(F0)、最大荧光(Fm),打开活化光,检测光光强设定为400 μmol·m-2s-1,测定光适应下初始荧光(F0′)、最大荧光(Fm′)、稳态荧光(Fs)。获得暗适应下最大光化学效率(Fv/Fm)和潜在活性(Fv/F0),光适应下实际光化学量子效率(ΦPSII)、激发能捕获效率(Fv′/Fm′)、光化学淬灭系数(qP)、非光化学猝灭系数(NPQ)。
1.4 统计及计算
用Excel 2016 软件进行数据整理,用Graph Pad Prism Version 7.0 (Graph Pad software,La Jolla,CA,USA)对测定数据作图,并进行方差分析、多重比较和相关性分析。
根冠比为地下鲜物质量/地上鲜物质量;N、P、K 积累量为根、茎、叶、全株的干物质量×对应的营养元素含量;N、P、K 分配率为各营养元素在根、茎、叶中的积累量/全株积累量×100。
D/%=(1-Fv′/Fm′)×100;P/%=qPFv′/Fm′×100;E/%=Fv′(1-qP)/Fm′×100[9]。其中D为天线热耗散,P为光化学耗散;E为非光化学能量耗散。
2 结果与分析
2.1 缺磷对黑木相思苗木生长的影响
表1表明:缺P 处理下,黑木相思苗木的生长受到显著影响,与全P 处理相比,苗高和地上部鲜质量显著下降,茎粗极显著下降,而地下部鲜质量在缺P 培养下略高于全P 培养,根冠比显著升高,说明黑木相思苗木通过增加根的生物量,减少地上部生物量,进而增加根冠比的适应性调整来应对缺P 胁迫。

表1 缺磷对黑木相思苗木生长的影响†Table 1 Effect of phosphorus deficiency on growth of A.menlanoxylon seedlings
2.2 缺磷对植株及器官营养元素积累和分配的影响
2.2.1 植株及器官营养元素积累
图1表明,缺P 处理苗木根、茎、叶和全株的N、P 和K 的积累量比全P 处理有不同程度的下降,除根中N 积累量没有达到显著水平外,根中K 积累量达到显著水平,其它都达到了极显著水平,下降幅度以P 积累量最大,根、茎、叶和全株的积累量分别下降了87.0%、89.6%、65.3%和76.4%,同时K 积累量在根、茎、叶和全株的降幅为23.5%、43.7%、35.7%和36.3%,而N 积累量的降幅分别为18.6%、32.7%、30.8%和30.1%。3 种营养元素的积累量最大降幅都出现在茎部,最小降幅P 元素出现在叶部,而K 和N 元素都出现在根部。

图1 缺磷对黑木相思营养元素积累的影响Fig.1 Effect of phosphorus deficiency on the accumulation of nutrients in Acacia melanoxylon
2.2.2 营养元素在根、茎、叶的分配
图2表明,缺P 培养下,N、P 和K 营养元素在根、茎和叶间的分配出现了不一致的规律性,N 和K 营养元素在根、茎和叶间的分配比例与全P 培养无显著差异,而P 营养元素在根和茎中的分配比例显著下降,在叶中的分配比例显著升高。说明在总吸收量下降的前提下,N 和K 元素仍保持体内稳定的分配比例,而P 元素随着吸收总量的大幅度下降,首先减少了根和茎中的分配,增加了在叶中的分配,这可能与叶是光合器官有关。

图2 缺磷对黑木相思营养元素分配的影响Fig.2 Effect of phosphorus deficiency on nutrient distribution of Acacia melanoxylon
2.2.3 根、茎、叶中营养元素的比值
图3表明,缺P 处理后苗木根、茎和叶中的N/K 比值略有上升,但未达到显著差异,而根、茎、叶中N/P 和K/P 比值极显著上升。说明缺P 培养3 个月时,对苗木体内N 和K 营养的平衡未产生显著影响,而N 和P、K 和P 的平衡遭到破坏,主要是P 元素的含量大幅下降引起。
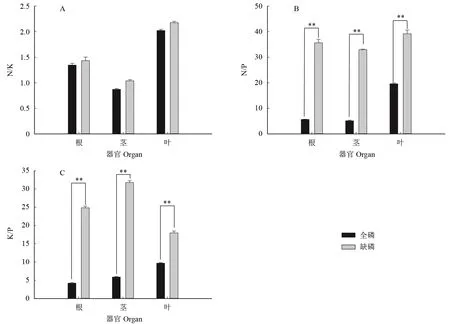
图3 缺磷对黑木相思营养元素比值的影响Fig.3 Effect of phosphorus deficiency on nutrient element ratio of Acacia melanoxylon
2.3 磷胁迫对叶片叶绿素荧光特性的影响
2.3.1 暗适应叶绿素荧光参数
表2表明,缺P 处理导致2 种形态叶的叶绿素荧光参数不同程度的下降,其中成年叶的F0、Fm和Fv/F0值下降显著和极显著,幼叶的F0、Fm值下降极显著。成年叶各参数的下降幅度大于幼叶,如幼叶的Fm值,全P 比缺P 处理下降了29.6%,而成年叶的Fm值下降了36.7%。全P 处理下,成年叶的Fv/Fm和Fv/F0大于幼叶,而缺P下,成年叶Fv/Fm和Fv/F0小于幼叶,说明成年叶PS Ⅱ反应中心的光化学效率和潜在活性受P 胁迫影响较大,而对幼叶影响较小,幼叶在P 胁迫下仍保持一定的光化学效率和潜在活性。

表2 缺磷对黑木相思2 种形态叶的叶绿素荧光参数影响†Table 2 Effect of phosphorus deficiency on chlorophyll fluorescence parameters of two morphological leaves of Acacia melanoxylon
2.3.2 光适应叶绿素荧光参数
图4表明,缺P 处理下,2 种形态叶的Fv′/Fm′、ΦPSII和qP比全P 处理有不同程度的下降,而NPQ 值上升,幼叶处理前后Fv′/Fm′、ΦPSII和qP值下降和NPQ 值上升均没有达到显著程度,成年叶Fv′/Fm′、ΦPSII和qP值下降和NPQ 值上升均达到显著程度。可见P 胁迫对成年叶PSII 反应中心的激发能捕获效率、光量子效率和电子传递活性影响都大于幼叶,对成年叶的热能耗散影响也大于幼叶。

图4 缺磷对黑木相思光适应叶绿素荧光的影响Fig.4 Effect of phosphorus deficiency on photosynthetic chlorophyll fluorescence of Acacia melanoxylon
2.3.3 光能吸收的分配
叶片光反应中心对光能的分配分为天线热耗散、光化学反应耗散和非光化学反应耗散3 种。表3表明,缺P 处理后,成年叶的天线热耗散值上升和光化学反应耗散值下降达到显著水平,幼叶的天线热耗散值和光化学反应耗散值也呈上升和下降趋势,但未达到显著水平,2 种形态叶的非光化学反应耗散值变化不显著。全P 和缺P 环境下,2 种形态叶的3 种光能分配百分率具有一致性。
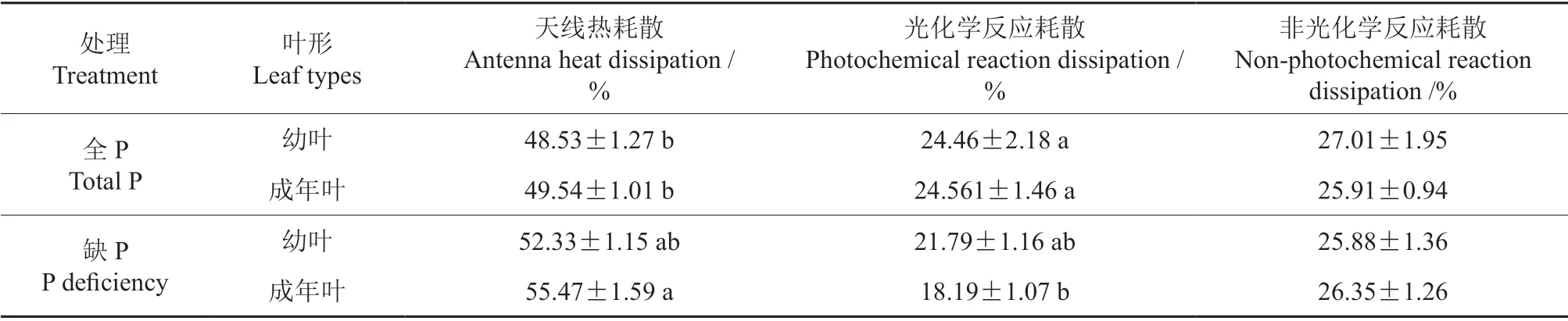
表3 缺磷处理下黑木相思2 种形态叶吸收光能的分配†Table 3 Distribution of light energy absorbed by two morphological leaves of Acacia melanoxylon under phosphorus deficiency treatment
2.4 营养元素含量与叶绿素荧光参数、苗木生长性状的相关性
2.4.1 营养元素含量与叶绿素荧光参数的相关性
Fv/Fm、ΦPSII、qP、NPQ 等荧光参数对植物非生物胁迫响应具有较好的指示性[10]。表4表明,叶N、P、K 含量与成年叶的荧光叶绿素参数Fv/Fm、ΦPSII呈显著和极显著的正相关性,叶P 含量与NPQ呈显著负相关,与qP相关性不显著;而叶N、P、K 含量与幼叶的荧光叶绿素参数无显著相关性。从相关性分析说明:叶片营养元素的变化主要影响了成年叶的叶绿素荧光性能,而对幼年叶的叶绿素荧光性能影响不大。
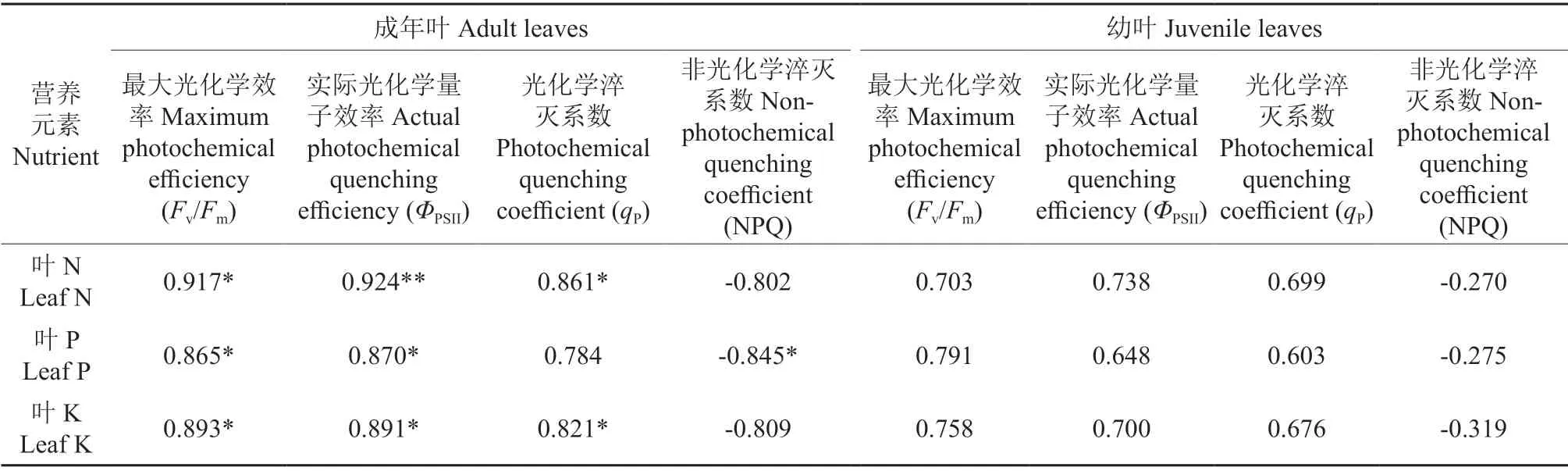
表4 黑木相思叶片营养与2 种形态叶的荧光叶绿素参数相关性†Table 4 Correlation between leaf nutrition and chlorophyll fluorescence of two morphological leaves of Acacia melanoxylon
2.4.2 营养元素含量与苗木生长性状的相关性
表5表明,N 素营养中除叶N 含量与株高具有显著正相关外,与其他各部位干重无显著相关性,K 素营养中除叶K、茎K 含量与株高具有显著正相关外,与其他各部位干质量也无显著相关性,而P 素营养中根P、茎P、叶P 含量与株高和叶干质量、地上干质量及全株干质量都具有显著的相关性,说明在缺P 培养下,N 和K 供应充足的时,只有P 素是苗木生长的限制因素。
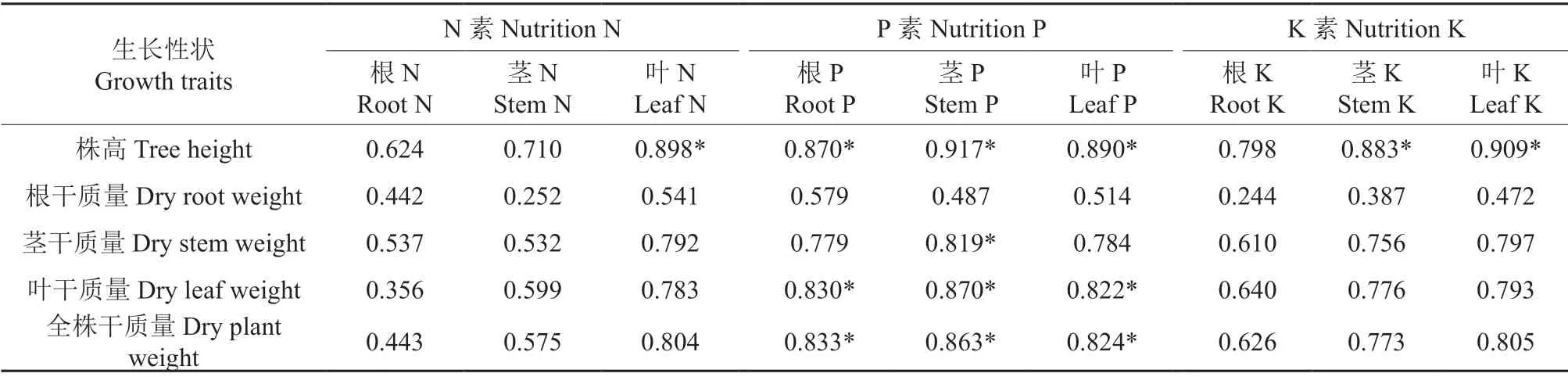
表5 黑木相思苗木各部位营养与生长性状的相关性†Table 5 Correlation between nutrition and growth in different part of A.melanoxylon seedlings
3 结论和讨论
3.1 磷胁迫对NPK 积累和分配的影响
在N 和K 供应充足的条件下,缺P 显著降低了黑木相思苗木对N、K 的吸收和积累,K 元素的积累量降低幅度大于N。这与杉木Cunninghamia lanceolata无性系在缺P 下对N、K、Ca、Mg 元素吸收量明显减少相似[11]。马尾松Pinus massoniana在极低P 胁迫下显著影响N、K 吸收量,且对N的抑制作用大于K[12],这可能是不同树种对N 和K 的需求和平衡不同。苹果砧木不同品种的P 利用效率也是影响N 吸收的原因之一,P 利用效率低的品种,对P 响应不敏感,P 胁迫后对N 吸收能力影响不大[4]。
为适应生存和生长的需要,植物对吸收的养分做出合理的分配。黑木相思在缺P 胁迫时,P 含量在根、茎中的分配比例大幅度下降,而保证了在叶中的分配,这与旱冬瓜Alnus nepalensis不同,P 水平供应对根、茎中P 含量敏感,而对叶中P 含量变化不明显相同[13]。P 元素在植物体内容易转运和重新分配,在营养生长阶段,叶片是P 素的积累中心,P 胁迫时,根和茎中的P 素主要转运到叶片中积累,这可能为了保证叶片进行正常的光合作用需要,进入生殖生长阶段,P 素主要转运到果实中积累,如大豆营养器官中积累的P 几乎全部被转运到荚果中,以致难以维持自身的发育[14]。一般N 素分布也集中在生命活动最活跃的部位,毛白杨Populus tomentosa施肥后随着植株生长,叶片中N 元素分配量增加[15]。橡胶树较轻度的低P 胁迫对N 素、K 素的分配无明显影响,无P 时,茎中的分配显著减少,而根的供给增加,且改变了N、K 养分的平衡[3]。P 胁迫下,黑木相思吸收N 和K 的总量虽然下降,但仍保持叶>茎>根的分配模式,且N/K 保持平稳,说明缺P 胁迫还没有对其生长产生致命的影响,黑木相思对P 缺乏具有一定的耐受度。
3.2 磷胁迫对荧光叶绿素性能影响
叶绿素荧光作为光合探针,能快速反应叶片PS Ⅱ反应中心的变化。P 胁迫后,黑木相思2 种形态叶的暗适应叶绿素荧光参数F0、Fm、Fv/Fm和Fv/F0不同程度下降,表示叶片PS Ⅱ原初光能转化效率及潜在活性减弱。有研究表明,缺P 对叶片N 代谢影响显著[16],而N 素影响叶绿素的合成,与光合色素含量和光合参数变化具有一致性[5]。黑木相思成年叶的暗适应叶绿素荧光参数大于幼叶,表明成年叶的PS Ⅱ原初光化学效率和能量传递效率高于幼叶,潜在活性较强,这与马尾松初生叶和次生叶叶绿素荧光性能相反[17]。可能是不同叶型光合机构有差异,黑木相思成年叶具有较强的光合能力,因此受P 胁迫影响也较大。缺P 导致光适应下Fv′/Fm′、ΦPSII和qP值下降和NPQ 值上升,Fv′/Fm′、ΦPSII和qP的下降,表明用于光化学反应的激发能减少,电子传递受阻,PS Ⅱ反应中心开放程度降低,而NPQ 增大表明将过剩的激发能以热能形式耗散,这是胁迫后启动的防御机制。黑木相思P 胁迫后叶片的叶绿素荧光参数的响应与N、Fe、Mg 养分胁迫[6,18-19]、盐胁迫[9,20]结果相似,因此叶绿素荧光技术可以实现对植株P 胁迫的无损检测。
3.3 养分含量与叶绿素荧光和生长的相关性
考来木Correa carmen叶中N、P 质量分数直接影响叶绿素水平和光合能力[6],水稻盐胁迫下叶片P、K 的积累量与叶绿素荧光有显著正相关[20]。黑木相思叶片N、P、K 含量与成年叶的叶绿素荧光参数相关性强,与幼叶的叶绿素荧光相关性弱,可见黑木相思苗期2 种形态叶的共存有利于提高苗木的抗逆性。
林木施肥研究表明,树体营养元素浓度与林木生长量、产量密切相关[21]。通过叶片营养元素临界值法可判断植物营养元素的亏缺[22],叶中N∶P化学计量比的阀值用来判别植物生长的主要限制因子[23]。P 胁迫后黑木相思叶片的N∶P 显著升高到39.1,P 含量成为严重的限制因子,黑木相思叶片P 含量与茎、叶及全株干质量的显著相关性进一步说明,在N 和K 供应充足而P 缺乏时,只有P 素是苗木生长的主要限制因素。
本研究只设计了全P 和缺P 两个处理试验,并在培养3 个月时获得数据,只反映了缺P 胁迫一段时期后黑木相思营养和叶绿素荧光参数的变化及其对生长的影响,苗木水培3 个月时尚未出现叶脉紫红色、老叶坏死、斑块易脱落[24]等缺P 病症。以后还需开展不同P 浓度和缺P 不同胁迫时间的试验,并结合代谢和光谱反射[25]等研究,建立P 养分内外诊断联动机制,为高效培育提供参考。
