基于SSR标记的山西玉米自交系的核心种质构建
2021-11-05尚春树常利芳白建荣
李 锐, 尚 霄, 尚春树, 常利芳, 闫 蕾, 白建荣
(1.山西农业大学农学院, 太原 030031; 2.山西强盛种业有限公司, 太原 030031)
玉米大约于16世纪从美洲引入我国[1],遗传基础相对狭窄一直是制约我国玉米育种的瓶颈。因此,种质的扩增、改良和创新是玉米育种的核心问题[2-3]。目前玉米种质资源与日俱增,其数目的庞大给种质保存、评价、研究和利用增加了工作量及难度。因此,为提高种质资源的管理质量和效率,构建玉米核心种质十分必要和迫切。
澳大利亚学者Frankel在1984年第一次提出了核心种质的想法, 即采取一定的取样方法,用尽可能少的种质材料最大可能代表整个资源的遗传信息和多样性[4]。早期对于核心种质的构建主要基于表型数据,虽易操作,但信息量少、易受环境影响而误差大。分子标记技术信息量大,不受环境影响,已经成为构建核心种质的主要方法[5]。目前,分子标记手段已经成功应用到玉米[6-9]、水稻[10]、籽用西瓜[11]、大白菜[12]、建兰[13]、野生山胡椒[14]等多种作物核心种质的构建中。
由于复杂的地理环境与生产条件,山西省形成了5个玉米生态种植区域[15],几乎包括了全国乃至全世界的玉米生态类型,每年审定的品种在100个以上。多年来,玉米育种团队在收集了大量国内外玉米种质的基础上,改良、培育了许多符合不同生态条件的优良自交系,这些自交系在很大程度上代表了山西玉米种质的特点。因此,构建山西玉米自交系的核心种质库具有实践指导意义,也是亟待解决的问题。本研究在用SSR分子标记对这些优良自交系进行遗传多样性分析的基础上,构建其核心种质,将为这些种质的保存、重点研究和有效利用提供可靠的依据。
1 材料和方法
1.1 材 料
1.1.1试验材料
224份玉米自交系由山西强盛种业有限公司提供, 其中包括目前生产上主要推广品种先玉335、先玉508、先玉696和郑单958的亲本自交系PH 6 WC(先玉335,先玉508,先玉696的母本)、PH 4 CV(先玉335父本)、PHB 1 M(先玉696的父本)、PH 5 AD(先玉508的父本)、郑58(郑单958母本)和昌7-2(郑单958父本)。
1.1.2SSR引物
采用中华人民共和国农业行业标准(NY/T 1432-2014《玉米品种鉴定DNA指纹方法》)的20 对核心引物和20 对辅助核心引物[16]。
1.2 实验方法
1.2.1SSR分析及聚类分析
DNA 提取、SSR分析及聚类分析方法参见李锐等[17]的方法。
1.2.2取样方法
本试验采用多次聚类结合优先取样法。首先根据所有种质遗传相似度,将样品进行聚类形成树状图。在聚类树状图中,按照一定的相似度进行分类。每一类中首先全部入选标准测验种和具有特异等位变异材料,然后从类内随机抽取一个样品保留进入下一轮聚类分析,若类内只有一个样品则该样品直接保留进入下一轮聚类分析。将所有保留的样品再次聚类,按之前同样的方法取样;再进入下一轮聚类、取样,循环,直至所取样品量达到理想取样。
1.2.3核心种质代表性检验和评价
用等位变异数、多态位点数、基因多样性、多态信息含量等指标评价原始种质和核心种质的遗传多样性。
2 结果与分析
2.1 核心种质库的构建及数据分析
利用40个SSR标记的171个等位变异,对224个自交系进行遗传距离聚类。在相似系数大约为0.71处可分为五大聚类群, 依据各聚类支中含有的不同种质类群的代表性自交系,确定类群的名称(图1)。从图中看出,瑞德群、兰卡斯特群是两个最大的群,二者数量之和超过了材料的90%;有10份材料属于唐四平头群;另有10个自交系的类群归属不明确,定为其他群。兰卡斯特群可明显看出,有335父本群、696与508父本群和其他兰卡斯特群,即有亚类分化趋势。在此基础上,再用最小距离逐步抽样法进行构建核心种质,即遗传距离小于一定数值的种质将首先被淘汰,但每次抽样都保留标准测验种和具有特异等位变异的自交系。共抽样8次,每次抽样构建自交系核心种质的遗传多样性如表1。由表1可以看出,随着抽样种质数目的减少,PIC和遗传多样性参数呈增加趋势;等位基因数与多态性位点减少极少。分析每次抽样后聚类图的类群结构发现,抽样6的聚类图仍然保持原始样本的类群划分(图2 ),但抽样7和抽样8的聚类图则不能保持原始样本的类群结构。因此,抽样6为最合适的核心种质群体。
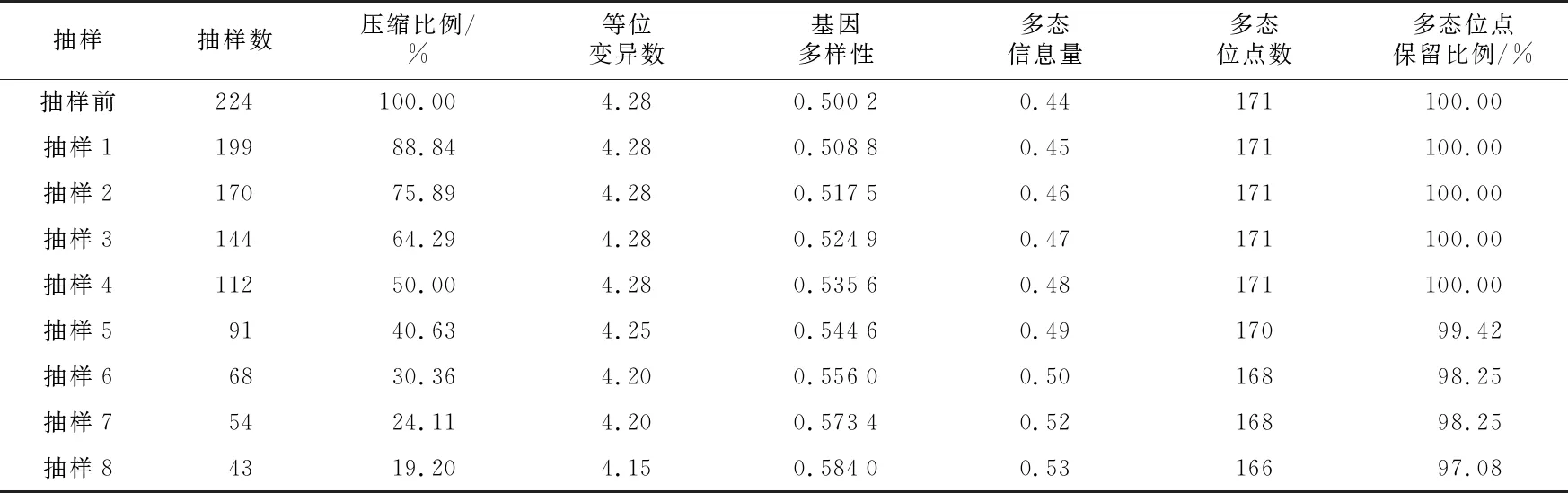
表1 聚类优先取样下各备选核心种质遗传多样性指数
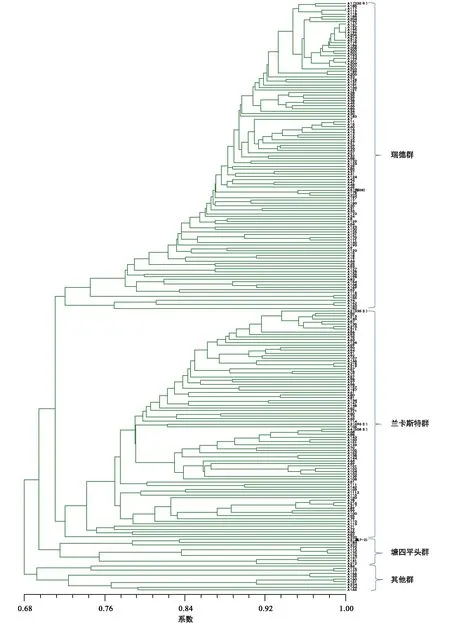
图1 基于SSR的224份原始玉米自交系聚类分析
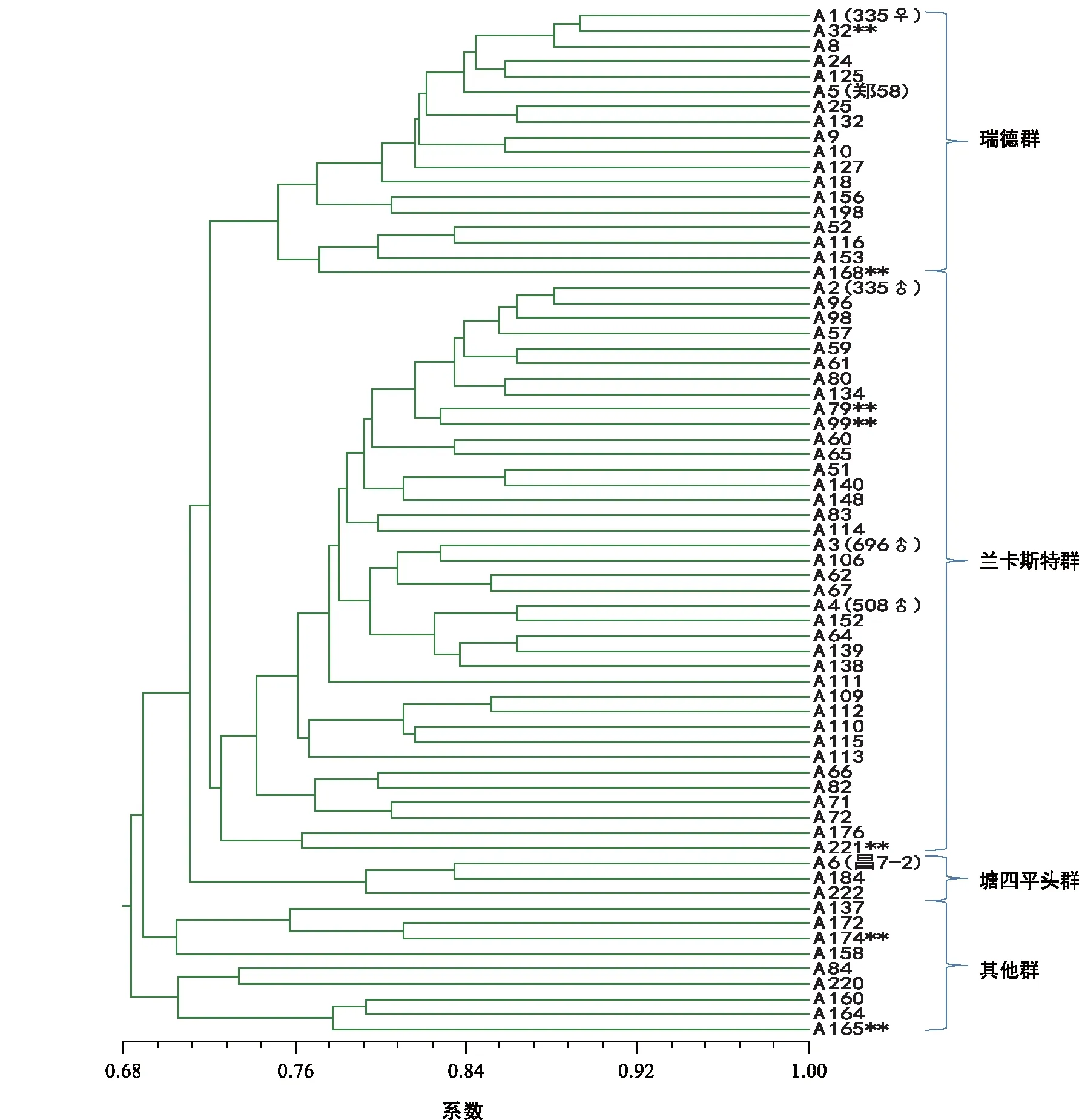
图2 基于SSR 标记的68份玉米核心自交系聚类分析
2.2 核心种质的确认与评价
原始种质224个自交系,依据30.36% 取样比例构建的核心种质包含 68个自交系,剩下的 156 个自交系为保留种质。对3组种质的等位变异数、基因多样性、多态信息量、多态位点数这 4 个遗传参数进行比较,结果如表2所示,核心种质的多态信息含量(0.50)和基因多样性(0.556 0)高于原始种质(0.44;0.500 2),等位变异数(4.20)和多态位点数(168)略小于原始种质(4.28;171)。同时,核心种质的这4个遗传参数均大于保留种质。3 组种质的4个遗传参数t检验结果显示,核心种质与原始种质、保留种质和原始种质均无明显差异(p>0.05)(表3)。以30.36% 的取样比例构建的包含 68个玉米自交系的核心种质能够充分代表原始种质的遗传多样性。
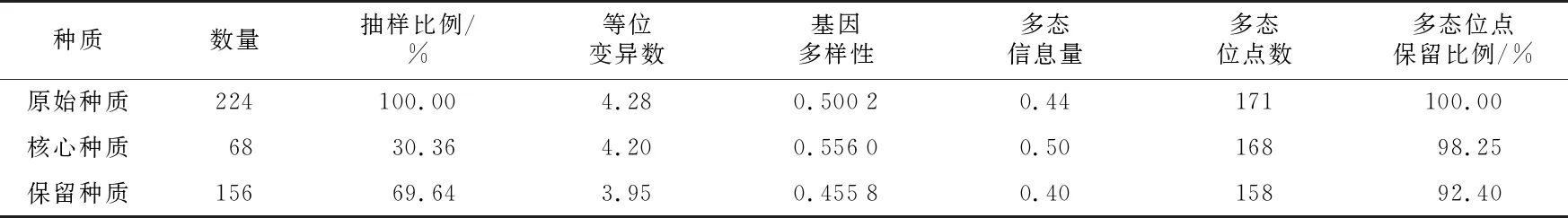
表2 核心种质、保留种质与原始种质遗传参数的比较

表3 核心种质、保留种质与原始种质遗传参数的t检验
3 讨 论
本研究根据30.36% 取样比例构建了224份玉米自交系的核心种质。3 组种质的4个遗传参数t检验结果表明,核心种质与原始种质、保留种质和原始种质之间均无明显差异,充分反映了该核心种质代表了原始种质的遗传多样性,保留种质也保留了原始种质的遗传多样性。
核心种质构建是种质资源研究的热点,构建核心种质的目的是以最小的样本数最大限度地代表原始样本群体的遗传多样性,构建的核心种质要有代表性、异质性和多样性、类型齐全。但是核心种质遗传代表性和数量之间存在非线性关系[17],因此,取样策略和取样比例是构建核心种质的关键。本研究的原始样本划分为4类杂种优势群,而玉米自交系的杂种优势类群的划分是玉米种质分析的主要内容和品种亲本组配的重要依据,因而构建的核心种质也要保留原杂种优势类群的特点,核心种质构建要考虑实际育种的需求和应用,构建好的核心种质要有利于种质的分析、保存和利用,不能盲目照搬已有的方法。因此,本研究采用多次聚类结合优先取样法,即在构建玉米自交系种质的核心种质时,虽然采用最小遗传距离逐步抽样法进行,但是,首先抽选标准测验种将它们作为类群划分的标准,使每次的抽样能反映其代表性;然后在其他抽样时先入选有特异等位变异的材料,因为它们代表了原始样本的异质性和多样性。每次循环聚类都按照这个原则进行。当进行到第7次抽样聚类图显示和原始样本聚类图的类群划分不一样时,就认为第6次抽样是合适的。研究结果显示,原始样本瑞德群的数量最多,但是核心种质中瑞德群的数量减少了很多,说明原始瑞德群中重复、冗余的样本较多;核心种质中其他群的数量相比原始群中其他群的数量相差不多。另外,本研究没有预先划定取样比例,完全根据逐步取样后能否完全反映原始样本的遗传多样性为前提,这和李自超等[18]认为各物种适宜的核心种质规模应按照具体物种的遗传多样性来定的思路是一致的。综上表明,本研究采用的策略具有合理性和实用性。构建的核心种质将有利于育种人员对育种材料的评价、保存、研究和在育种中的有效利用。
