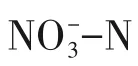硫自养/异养协同反硝化深度脱氮处理三氯蔗糖生产废水
2021-11-03耿雅雯刘锋冯震陈俊张雪智
耿雅雯,刘锋,2,冯震,陈俊,张雪智
(1 苏州科技大学环境科学与工程学院,江苏 苏州 215009;2 城市生活污水资源化利用技术国家地方联合工程实验室,江苏 苏州 215009)
三氯蔗糖主要通过单酯法合成,是一种应用广泛的食品甜味剂[1-2]。其生产过程中产生的废水(粗品母液、萃取水层和洗涤废水等)具有高化学需氧量(COD)、高氨氮(NH+4-N)的特点,可生化性差且处理难度较大[3]。此类人工甜味剂废水常用处理工艺有“混凝+铁碳微电解+Fenton+EGSB(膨胀颗粒污泥床)+两级A2O”[4]和“微电解+Fenton+混凝+ABR(折流式厌氧反应器)+A/O”[5-6]及“MVR(机械式蒸汽再压缩技术)+A2O”等,主要针对去除COD、氨氮等污染物,但对总氮的去除效果不佳,普遍存在二沉池出水硝态氮较高的现象。随着国家对工业废水总氮排放要求的日益严格,现有工艺普遍不能满足排放标准,故需要增设深度脱氮单元。
生物反硝化技术一直被认为是去除废水中硝态氮最经济可行的技术,分为自养反硝化和异养反硝化两种[7]。传统的异养反硝化需外加碳源作为电子供体,易造成二次污染,且污泥产量大,运行费用高[8]。近年来,硫自养反硝化因其处理效率高、污泥产量少和能耗低等优势在深度脱氮领域受到广泛关注[9]。但其反应过程产生的H+会导致pH 降低,从而影响脱氮速率,而研究中常用的石灰石等碱性物质又会导致出水硬度增加[10]。因此,为解决单一电子供体的反硝化体系中存在的问题,有研究者采用混合电子供体进行协同反硝化。李祥等[11]通过异养反硝化与硫自养反硝化联合,发现协同反硝化脱氮不仅能够实现反应系统的酸碱平衡,同时能够提高自养反硝化的速率,减少污泥产量。而目前此类的研究大多停留在人工配水阶段,其能否直接用于化工企业产生的高浓度硝态氮废水的处理还需要进一步研究。
S0、Na2S和Na2S2O3是硫自养反硝化中常用的电子供体。S0溶解度较低,在水中的传质速率较慢,影响反应器的启动及脱氮速率。Na2S不稳定,且其碱度较强,易对系统的酸碱环境造成冲击[12]。故本研究选用溶解度高、pH 稳定的Na2S2O3和葡萄糖作为电子供体,考察硫自养/异养协同反硝化系统对三氯蔗糖废水二级处理出水的可行性及连续运行的效果,并通过高通量测序分析协同反硝化污泥的微生物群落结构及多样性,以期为实际工艺运行提供理论依据和技术参数。
1 材料与方法
1.1 实验装置及污水来源
实验装置如图1所示,采用上流式高效流化床反硝化反应器,其主体由亚克力柱组成,反应柱高60cm,内径为8cm,有效体积为3L。采用水浴加热的方式将反应温度控制在(35±1)℃,通过蠕动泵将进水送入反应器。
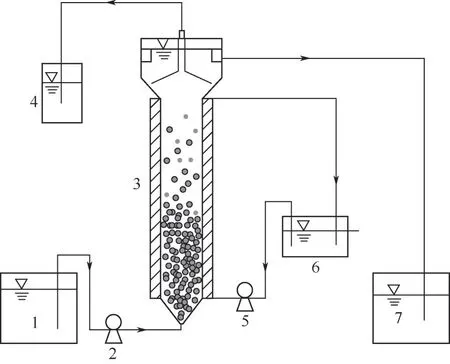
图1 实验装置
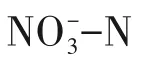
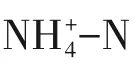
1.2 实验内容
1.2.1 协同反硝化反应器的启动(实验阶段Ⅰ)
1.2.2 协同反硝化反应器的运行(实验阶段Ⅱ)
反应器各阶段运行参数如表1所示。
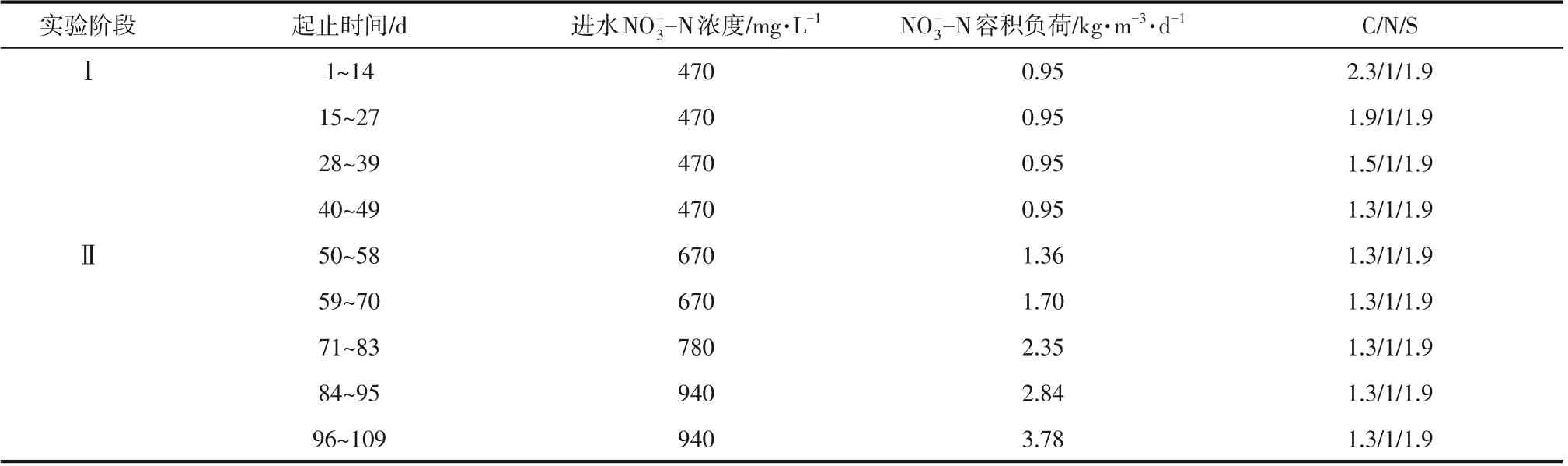
表1 反应器各阶段运行参数
1.3 分析方法
微生物菌落测定:采用MiSeq型高通量测序仪(美国Illumina 公司)对接种污泥及协同反硝化污泥进行微生物多样性测定及分析。
2 结果与讨论
2.1 协同反硝化反应器的启动
研究发现,反硝化菌的最适生长pH 为6.8~8.2[13],故在不投加pH 缓冲剂的条件下,向异养反硝化反应器中投加Na2S2O3,使得异养反硝化产生的碱度能够满足硫自养反硝化菌生长的需要,并以此来启动硫自养/异养协同反硝化系统。由图2 可知,从投加Na2S2O3的第3 天开始,反应器出水pH由7.8降低至7.5且基本保持稳定,证明在C/N/S 为2.3/1/1.9的条件下,硫自养反硝化菌得到最大限度的生长,其反应产生的H+导致pH降低。国内外对硫自养反硝化的研究大多集中在以单质硫为电子供体,但是单质硫不溶于水,其传质效率较低,故反应器启动较慢。马航等[14]发现,在以单质硫为电子供体的膜生物反应器中,需要6~10 天的时间系统才能稳定运行。而液态硫源Na2S2O3的传质速率更快,在反应器内与污泥充分接触,从而刺激硫自养反硝化细菌的生长,有效提高反应器的启动速率。
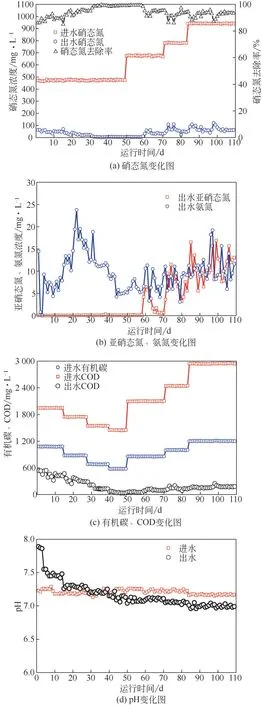
图2 反应器运行过程

2.2 协同反硝化反应器的运行
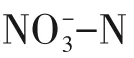
表2 国内外硫自养反硝化去除高浓度NON研究成果
表2 国内外硫自养反硝化去除高浓度NON研究成果
反应器类型填充床[25]CHSAD[26]批次实验[27]缺氧反应器[28]本研究电子供体类型单质硫单质硫+甲醇硫代硫酸钠硫代硫酸钠硫代硫酸钠+葡萄糖废水来源人工配水人工配水人工配水废碱液三氯蔗糖生产废水二级处理出水是否需pH缓冲NO-3-N去除负荷/kg·m-3·d-1是否是否否1.47 2.16 0.20 0.25 3.52
而本研究构建的硫自养/异养反硝化具有较高的脱氮水平和脱氮优势,分析其原因如下:①Na2S2O3易溶于水,其将电子供体向污泥的传质速率更高,满足硫自养反硝化菌工作的需要,研究表明,相较于单质硫和硫化物,以Na2S2O3为电子供体的硫自养反硝化脱氮效果最好[23];②硫自养反硝化与异养反硝化结合不仅可以实现酸碱互补,避免了pH 缓冲剂的投加,减少了药剂成本,碳源的投加还对硫自养反硝化有一定的促进作用,使运行更加稳定;③采用上流式流化床高效反硝化反应器,可以达到泥水的完全混合,同时反应产生的气体对泥水具有一定的搅动作用,促进反硝化的进行;④根据国家标准方法[24]测定,1g/L的葡萄糖可提供约1000mg/L 的COD,而1g/L 的Na2S2O3仅提供约250mg/L的COD,故该协同反硝化系统可有效减缓异养反硝化易造成出水有机物二次污染的问题。
2.3 微生物群落分析
2.3.1 微生物群落的丰富性及多样性分析
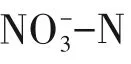
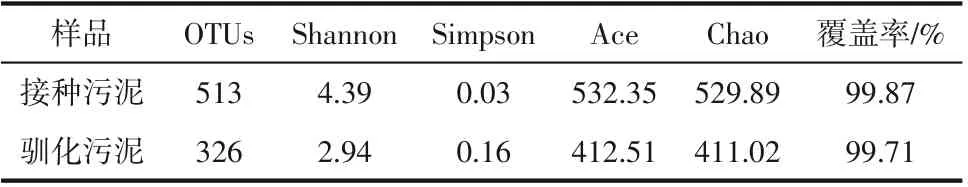
表3 污泥样品中的生物多样性指数
2.3.2 微生物在不同物种水平上的分析
为了证明微生物菌群的变化,对微生物在不同物种分类学水平上进行分析。图3(a)反应了驯化前后污泥中微生物菌群在门水平上的变化,接种污泥的微生物门种类多而分散,其中Proteobacteria、Spirochaetes、Chloroflexi、Zixibacteria 为 接 种 污 泥中丰度较高的门,其占比分别为27.44%、14.40%、13.70%和9.74%。而驯化后的污泥以Proteobacteria为主,其占比为89.65%,其余菌门单独所占的比例均不超过2%。虽然接种污泥和驯化后的污泥中的优势菌门均为Proteobacteria,但其相对丰度存在明显的差异,证明在该协同反硝化系统中,Proteobacteria 中含有大量与反硝化相关的微生物,其丰度大幅度增加增强了脱氮的效果。且在Proteobacteria下属的纲级别微生物菌群仍为样品中的优势菌群,如图3(b)所示,Gammaproteobacteria为主要的变形菌纲,其占比为81.60%,是协同反硝化污泥的优势菌纲。Han等[30]在混养反硝化中发现,Gammaproteobacteria 为优势菌纲,其中大多为反硝化菌种。

图3 微生物菌落在门、纲水平分布
为了更深入地了解硫自养/异养协同反硝化污泥的功能菌群和优势菌种,对污泥中微生物的属分类进行分析,结果如图4所示。接种污泥和驯化后的污泥在属水平上具有明显的差异,其共有的菌属较少。驯化后的污泥以反硝化菌为主,其中自养反硝化菌和兼养反硝化菌占比较高,分别为45.69%和25.38%,异养反硝化菌占比较少,为8.16%。这也说明在该协同反硝化系统中,硫自养反硝化占据更大的比例,投加的碳源只有部分用于异养反硝化。推测在C/N/S 为1.3/1/1.9 的混养条件下,硫源的投加对异养反硝化菌有一定的抑制作用,而在适量碳源存在的条件下,硫自养反硝化菌仍可以选择性富集,使反应系统保持较好的脱氮效果,故认为协同反硝化可以作为有效的生物脱氮途径来减缓异养反硝化过程中需要大量投加碳源和pH缓冲剂的问题。
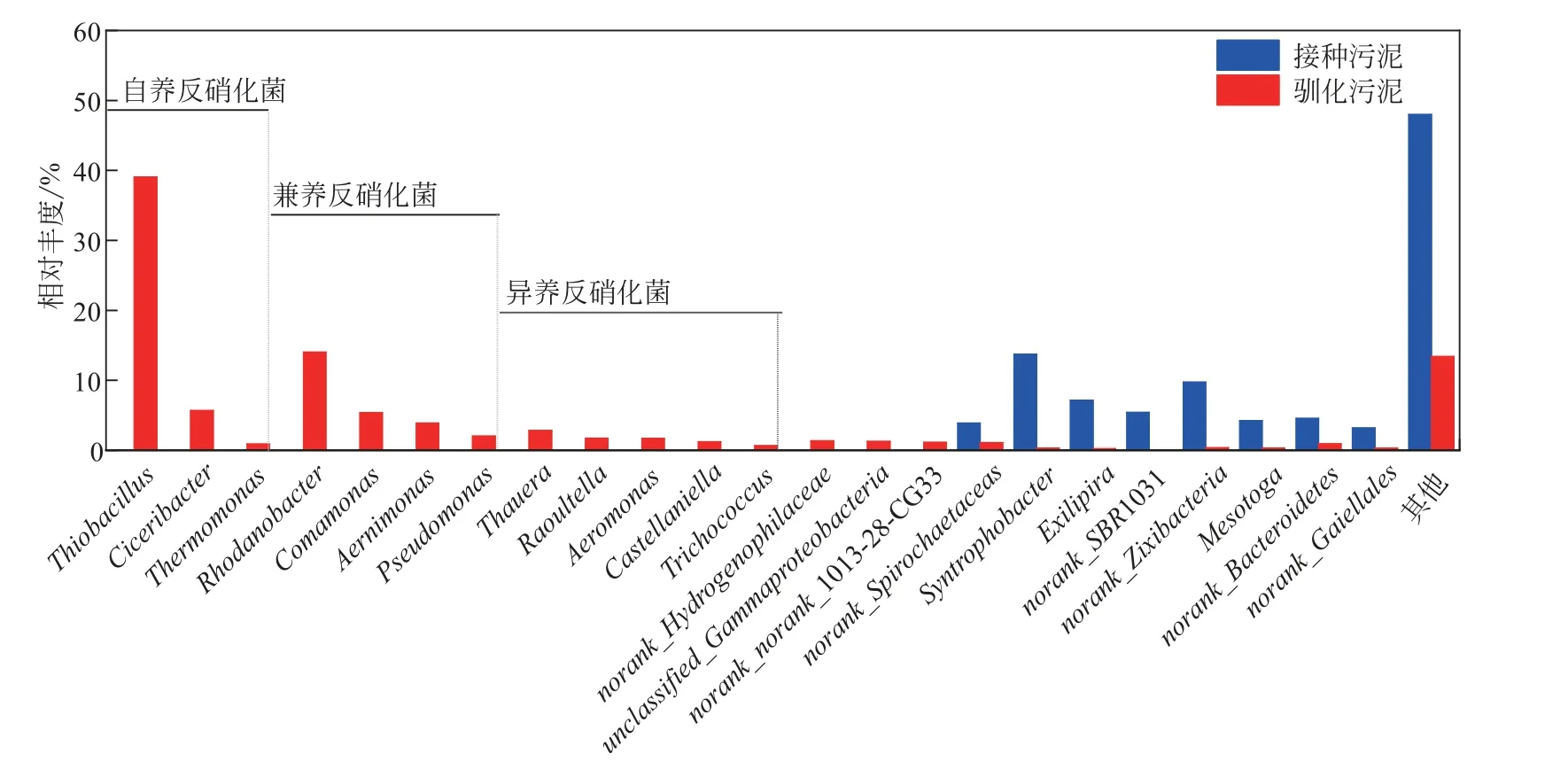
图4 微生物菌落在属水平分布
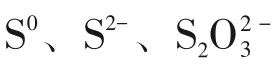
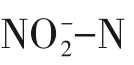
3 结论
(1)在硫自养/异养协同反硝化反应器中,将C/N/S 控制在1.3/1/1.9,在无需pH 缓冲剂的条件下,自养反硝化和异养反硝化能够实现酸碱互补,且达到电子供体的全部利用,出水无有机物的二次污染。