生物炭-泥炭复合基质对马缨杜鹃生长和生理的影响
2021-11-02卜晓莉姬慧娟马青林马文宝张俊浩俞睿思薛建辉
卜晓莉, 姬慧娟, 马青林, 马文宝,①, 张俊浩, 俞睿思, 薛建辉,3, 陈 娟
〔1. 南京林业大学 南方现代林业协同创新中心, 江苏 南京 210037;2. 四川省林业科学研究院 森林和湿地生态恢复与保育四川重点实验室, 四川 成都 610081;3. 江苏省中国科学院植物研究所(南京中山植物园), 江苏 南京 210014; 4. 西南民族大学, 四川 成都 610041〕
泥炭是园艺生产中常用的传统栽培基质之一,但泥炭的过度开采和使用会增加温室气体释放,破坏湿地生态系统的生态平衡[1-2]。为合理利用泥炭资源,园艺工作者尝试使用各类生物基质替代泥炭。生物炭是将含碳量丰富的生物质(如木材、秸秆、稻壳、玉米芯和家禽粪便等)在350 ℃~900 ℃低氧条件下热解而得到的一种低容重、高孔隙度、高阳离子交换量的碳质材料[3-5]。将富含碳的农林废物转化为生物炭,不仅可以实现农业废弃物的回收再利用,为园艺生产创造可再生的新材料,也可以为温室气体的减排做出贡献[6]。
使用生物炭作为栽培基质,不仅可以促进植物生长,还能提高植物对多种病害的抗性[7]。Nemati等[8]的研究结果显示:木炭可以作为K、Fe、Mn和Zn等营养元素的来源,具有完全取代珍珠岩和部分取代泥炭的应用潜力;史华等[9]的研究结果表明:含质量分数10%秸秆生物炭的栽培基质最适合于火鹤花(AnthuriumscherzerianumSchott)的种植,但秸秆生物炭添加量过高则会抑制其生长;陈慧等[10]的研究结果表明:在栽培基质中添加不同比例的木炭、竹炭和稻壳炭,对桑树(MorusalbaLinn.)叶片的品质有不同程度的影响;吴志庄等[11]认为,适量使用生物炭可有效改善三叶青(TetrastigmahemsleyanumDiels et Gilg)的光合特性,但生物炭使用量太高则不利于植株的生长和光合效率的提高;朱奕豪等[12]对百合(Liliumbrowniivar.viridulumBaker)种植的研究结果表明:质量分数10%花生壳生物炭与草炭复合基质中的百合长势最佳、抗性最强,但当花生壳生物炭添加量增加到质量分数20%时,对百合生长则产生抑制作用。上述研究结果表明:不同种类植物对于生物炭种类以及使用量和配比的需求有一定差异,且生物炭过量会抑制植物生长。因此,在生产和应用过程中,需根据不同植物种类的生长需求,选择合适的生物炭种类以及使用量和配比。
杜鹃花属(RhododendronLinn.)植物具有种子细小、幼苗娇弱且生长缓慢等特点,在播种育苗时需要使用大量泥炭,因此,在保证观赏价值和生产效率的前提下,寻找合适的替代品以减少栽培基质中泥炭的用量,对于实现可持续的杜鹃花苗圃生产具有十分重要的意义。本项目组前期的研究结果[13]表明:不同类型生物炭的浸提液对4种杜鹃花属植物种子萌发的影响效应各异,其中,玉米芯炭浸提液对部分种类表现出低浓度促进、高浓度抑制的“毒物兴奋效应”,而稻壳炭浸提液可促进这些种类的种子萌发,玉米芯炭和稻壳炭浸提液可抑制大白杜鹃(R.decorumFranch.)的种子萌发,且浓度越高,抑制作用越强。因而,在育苗培育过程中,不同类型的生物炭除影响杜鹃属植物种子萌发外,对杜鹃属植物的生长和生理代谢有何影响?其作用机制和规律如何?这些问题还需要深入的研究。
为进一步明确不同类型生物炭和泥炭复合基质对杜鹃花属植物生长和生理代谢的影响,作者以马缨杜鹃(RhododendrondelavayiFranch.)1年生幼苗为研究对象,以不同比例的木炭或稻壳炭与泥炭组成复合基质,比较各组复合基质的理化性质差异,并研究马缨杜鹃幼苗生长及其叶片的光合生理特性和矿质元素含量的变化,分析幼苗生长和叶片光合生理指标与复合基质理化性质的相关性,以期揭示马缨杜鹃幼苗对生物炭-泥炭复合基质的适应特性,为杜鹃花属植物育苗栽培生产过程中生物炭的合理应用提供基础研究数据。
1 材料和方法
1.1 材料
试验用木炭和稻壳炭(最高热解温度500 ℃,保温 1 h)购自大连松森物产有限公司;木炭和稻壳炭研碎后过100目筛,备用。泥炭(Klasmann 876)购自德国Klasmann-Deilmann公司,粒径0~5 mm。供试1年生马缨杜鹃幼苗购自四川省甘孜藏族自治州泸定县新兴苗圃。
1.2 方法
1.2.1 基质配比和幼苗栽培 以泥炭为基本基质,分别添加体积分数10%、20%、30%和40%的木炭(W)和稻壳炭(R)组成复合基质。CK(对照):泥炭;W10:V(木炭)∶V(泥炭)=10∶90;W20:V(木炭)∶V(泥炭)=20∶80;W30:V(木炭)∶V(泥炭)=30∶70;W40:V(木炭)∶V(泥炭)=40∶60;R10:V(稻壳炭)∶V(泥炭)=10∶90;R20:V(稻壳炭)∶V(泥炭)=20∶80; R30:V(稻壳炭)∶V(泥炭)=30∶70;R40:V(稻壳炭)∶V(泥炭)=40∶60。按上述比例将过筛后的生物炭与泥炭混匀,装入口径16.0 cm、底径12.5 cm、高17.0 cm的塑料花盆中,每盆装2.5 L复合基质,每处理设置3个重复,每个重复5盆,每盆定植1株幼苗,共计135盆。
于2019年5月将供试盆栽幼苗置于四川省甘孜藏族自治州泸定县新兴苗圃的温室中,于自然光照、昼夜平均温度分别为15 ℃和8 ℃的条件下培养;每盆一次性施入缓释肥0.6 g(奥绿肥501,以色列化工集团),生长季(3月至10月)3~4 d浇水1次,浇水量标准为基质达最大持水量,其他时间进行常规栽培管理。2020年9月生长期结束(定植时间16个月),将所有供试幼苗完整挖出,用蒸馏水清洗根部后用于各项指标测定。
1.2.2 栽培基质理化指标测定 参照相关文献[14],采用环刀法测定泥炭、木炭和稻壳炭的容重,以及各组复合基质的容重、通气孔隙度、总孔隙度和持水孔隙度。按固液比(m∶V)1∶20加入去离子水浸提2 h,分别采用PHSJ-5型酸度计(上海仪电科学仪器股份有限公司)和NDDS-11A型电导率仪(南京南大万和科技有限公司)测定泥炭、木炭和稻壳炭以及各组复合基质的pH值和电导率。
采用Elementar Vario EL Ⅲ元素分析仪(德国Elementar公司)测定泥炭、木炭和稻壳炭中N含量;采用NexION 300X电感耦合等离子体质谱仪(美国PerkinElmer公司)测定泥炭、木炭和稻壳炭中P、K、Mg、Na、Fe、Mn和Zn含量;采用Quanta 200环境扫描电子显微镜(美国FEI公司)观察生物炭的孔径大小及表面形态特征。
将各组复合基质烘干至恒质量,研磨并过筛(孔径1 mm),称取约0.05 g样品进行元素含量测定。采用碱解扩散法[15]150-151测定复合基质中有效氮(N)含量;采用NaHCO3浸提-钼锑抗比色法[15]168-169测定复合基质中速效磷(P)含量;采用乙酸铵提取法[15]194-195测定复合基质中速效钾(K)含量;采用NexION 300X电感耦合等离子体质谱仪测定复合基质中Mg、Na、Fe、Mn和Zn含量。
1.2.3 植株各项指标测定 每个处理随机选择10株样株。分别用钢卷尺(精度1 mm)和电子游标卡尺(精度0.01 mm)测量株高和地径,并统计单株叶片数;采用YMJ-B手持叶面积仪(浙江托普云农科技股份有限公司)测量单叶的叶面积,并计算单株叶面积;将根、茎和叶片分开,分别置于105 ℃干燥箱中杀青30 min,然后于80 ℃烘干至恒质量,冷却后称量单株的根、茎和叶的干质量,并计算单株总干质量。
取干燥叶片,研磨并过筛(孔径1 mm),称取约0.05 g样品进行元素含量测定。采用Elementar Vario EL Ⅲ元素分析仪测定叶片中N含量;采用NexION 300X电感耦合等离子体质谱仪测定叶片中P、K、Mg、Na、Fe、Mn和Zn含量。
在每个重复处理中随机选择10枚完全展开的叶片,用SPAD-502 Plus叶绿素测定仪(日本Konica Minolta公司)测定叶绿素相对含量(SPAD),结果取平均值。于2020年9月连续晴天的8:30至11:30,在每个处理中随机选择5株样株,选取样株自上而下第5枚完全展开叶,采用LI-6400便携式光合作用测定仪(美国LI-COR公司)测定叶片的净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)和气孔导度(Gs),并根据公式“WUE=Pn/Tr”计算水分利用效率(WUE)。
上述涉及的生长和理化指标均重复测量3次,结果取平均值。
1.3 数据分析
使用SPSS 13.0软件进行数据统计分析,使用Origin 8.5软件绘图。采用双因素分析(two-way ANOVA)方法,运用LSD多重比较分析各组复合基质对马缨杜鹃生长和光合生理参数的影响效应;并对幼苗各项生长指标和光合生理指标与复合基质理化指标进行相关性分析。
2 结果和分析
2.1 栽培基质的理化性质
2.1.1 泥炭、木炭和稻壳炭的理化性质 供试泥炭、木炭和稻壳炭理化性质见表1。结果显示:木炭的容重和电导率明显高于泥炭和稻壳炭,其容重分别为泥炭和稻壳炭的3.1和2.5倍,电导率分别为泥炭和稻壳炭的3.5和2.4倍。2种生物炭均呈弱碱性,且木炭的pH值略高于稻壳炭,而泥炭则呈弱酸性。除N和Zn含量外,2种生物炭的P、K、Mg、Na、Fe和Mn含量均高于泥炭,且木炭各项指标与泥炭的差异更明显。木炭和稻壳炭中K、Fe和Mn较丰富,木炭中Mg和Na也较丰富,其中,木炭和稻壳炭中K含量分别为泥炭的4.3和3.4倍,Fe含量分别为泥炭的3.2和1.7倍,Mn含量分别为泥炭的2.4和9.6倍;木炭中Mg含量为泥炭的2.4倍,Na含量为泥炭土的3.1倍。

表1 供试泥炭、木炭和稻壳炭的理化性质比较
2.1.2 木炭和稻壳炭的结构 对木炭和稻壳炭的结构进行观察,结果(图1)显示:木炭整体呈薄片状,形状不规则;具有疏松多孔结构,孔径较小且较为接近,孔径范围为1.15~2.31 μm(图1-A)。稻壳炭形状不规则;孔径较大,排列致密,孔径范围为3.15~10.00 μm(图1-B)。
2.1.3 生物炭-泥炭复合基质的理化性质 不同比例生物炭-泥炭复合基质的理化性质见表2。结果显示:分别添加体积分数20%、30%和40%木炭的W20、W30和W40复合基质的容重较对照(泥炭)分别增大了38.9%、66.7%和77.8%,差异达显著(P<0.05)水平;而添加不同比例稻壳炭的复合基质的容重与对照无显著差异。添加木炭和稻壳炭的复合基质的pH值总体增加,其中,W30和W40复合基质的pH值显著高于对照,W40复合基质的pH值最高(pH 6.56);其他6组复合基质的pH值与对照无显著差异。在8组复合基质中,仅W20、W30和W40复合基质的电导率较对照分别增加了17.7%、26.2%和52.5%,差异达显著水平;其他5组复合基质的电导率与对照无显著差异。
由表2可见:添加木炭和稻壳炭的复合基质的通气孔隙度大于对照,其中,W30和W40以及分别添加体积分数20%、30%和40%稻壳炭的R20、R30和R40复合基质的通气孔隙度较对照分别增大了83.3%、128.1%、57.7%、96.9%和162.2%,差异达显著水平;其他3组复合基质的通气孔隙度与对照无显著差异。除添加体积分数10%稻壳炭的R10复合基质,其他7组复合基质的持水孔隙度不同程度小于对照,其中,W30、W40和R40复合基质的持水孔隙度较对照分别减小了13.0%、25.3%和15.6%,差异达显著水平;其他5组复合基质的持水孔隙度与对照无显著差异。各组复合基质的总孔隙度也不同程度大于对照,其中,R20、R30和R40复合基质的总孔隙度较对照分别增大了9.4%、12.1%和17.2%,差异达显著水平;其他5组复合基质的总孔隙度与对照无显著差异。

图1 扫描电子显微镜下木炭(A)和稻壳炭(B)的结构Fig. 1 Structures of wood biochar (A) and rice husk biochar (B) under SEM

表2 不同比例生物炭-泥炭复合基质的理化指标比较
由表2还可见:复合基质中N、P、Mg和Zn含量略有变化,但与对照无显著差异。除R10复合基质外,各组复合基质的K含量高于对照;其中,W20、W30、W40、R30和R40复合基质的K含量较对照分别升高了17.8%、24.1%、38.3%、23.2%和33.1%,差异达显著水平;其他3组复合基质的K含量与对照无显著差异。除R10复合基质外,各组复合基质的Na含量高于对照,其中,W30和W40复合基质的Na含量较对照分别升高了14.3%和29.8%,差异达显著水平;其他6组复合基质的Na含量与对照无显著差异。仅W40复合基质的Fe含量较对照升高25.7%,差异达显著水平;其他各组复合基质的Fe含量与对照无显著差异。各组复合基质的Mn含量不同程度高于对照,其中,W40、R20、R30和R40复合基质的Mn含量较对照分别升高了32.3%、29.0%、77.4%和129.0%,差异达显著水平;其他4组复合基质的Mn含量与对照无显著差异。
综合比较结果显示:与对照相比,添加不同比例木炭和稻壳炭的复合基质的理化性质均发生了不同程度的改变。其中,随木炭体积分数的提高,复合基质的容重、pH值、电导率、通气孔隙度、总孔隙度以及P、K、Mg、Na、Fe和Mn含量总体上逐渐升高,而持水孔隙度和N含量总体上逐渐降低,Zn含量无明显变化;随稻壳炭体积分数的提高,复合基质的pH值、电导率、通气孔隙度、总孔隙度以及P、K、Mg、Na、Fe和Mn含量总体上逐渐升高,而持水孔隙度和N含量总体上逐渐降低,容重和Zn含量无明显变化,P含量波动变化。在8组复合基质中,添加体积分数40%木炭的复合基质的容重、pH值、电导率以及除N和Mn含量外的其他元素含量均最高,通气孔隙度也较高,但持水孔隙度最小。
2.2 马缨杜鹃幼苗生长以及叶片光合特性和矿质元素含量的差异
不同比例生物炭-泥炭复合基质中马缨杜鹃幼苗生长指标的差异见表3,叶片的光合生理指标的差异见表4,叶片中矿质元素含量的差异见表5。
2.2.1 幼苗生长指标的差异 由表3可见:除添加体积分数10%木炭和稻壳炭的W10和R10复合基质外,在其他复合基质中幼苗株高高于对照(泥炭),其中,在添加体积分数20%木炭的W20复合基质中幼苗株高较对照增加了17.5%,差异达显著(P<0.05)水平;而在其他7组复合基质中幼苗株高与对照无显著差异。与对照相比,在各组复合基质中幼苗地径不同程度增大或减小,但与对照无显著差异。与对照相比,在各组复合基质中幼苗单株叶片数不同程度增多或减少,其中,在添加体积分数40%稻壳炭的R40复合基质中幼苗单株叶片数较对照增多了22.9%,差异达显著水平;而在其他7组复合基质中幼苗单株叶片数与对照无显著差异。与对照相比,在各组复合基质中幼苗单株叶面积不同程度增大或减小,其中,在W20和R40复合基质中幼苗单株叶面积较对照分别增大了18.1%和22.4%,差异达显著水平;在其他6组复合基质中幼苗单株叶面积与对照无显著差异。
由表3还可见:在W20复合基质中幼苗单株的根、茎和叶的干质量以及总干质量较对照分别增加了16.7%、24.5%、21.4%和21.2%,差异达显著水平;在添加体积分数30%木炭的W30复合基质中幼苗单株茎干质量和单株叶干质量较对照分别增加了26.6%和17.1%,差异达显著水平;在添加体积分数30%稻壳炭的R30复合基质中幼苗单株叶干质量较对照增加了11.7%,差异达显著水平;在R40复合基质中幼苗单株的根和叶的干质量以及总干质量较对照分别增加了14.1%、21.8%和16.8%,差异达显著水平;在其他复合基质中幼苗单株的根、茎和叶的干质量以及总干质量略有差异,但均与对照无显著差异。
综合比较结果显示:随复合基质中木炭体积分数的提高,幼苗株高和地径以及单株的叶片数、叶面积、根干质量、茎干质量、叶干质量和总干质量总体上先升高后降低;其中,在W20和W30复合基质中,幼苗的各项生长指标不同程度高于对照;但在添加体积分数40%木炭的W40复合基质中,幼苗单株的叶片数、叶面积、根干质量、叶干质量和总干质量均最低,较对照分别减小了10.1%、12.5%、9.0%、7.8%和4.9%。随复合基质中稻壳炭体积分数的提高,幼苗株高先升高后降低,地径以及单株的叶片数、叶面积、根干质量、茎干质量、叶干质量和总干质量总体上逐渐升高;其中,在R10复合基质中,幼苗的各项生长指标总体上不同程度低于对照;而在R40复合基质中,幼苗的各项生长指标不同程度高于对照。
2.2.2 叶片光合生理指标的差异 由表4可见:在W20复合基质中叶片的净光合速率显著高于对照,在W20、W30、R30和R40复合基质中叶片的蒸腾速率显著高于对照;在各组复合基质中,叶片的叶绿素相对含量、胞间CO2浓度、气孔导度和水分利用效率与对照无显著差异。总体上看,随复合基质中木炭体积分数的提高,叶片的叶绿素相对含量、净光合速率、蒸腾速率和气孔导度先升高后降低,水分利用效率逐渐降低;随复合基质中稻壳炭体积分数的提高,叶片的叶绿素相对含量、净光合速率、蒸腾速率和气孔导度逐渐升高,水分利用效率总体上逐渐降低。
在4组木炭-泥炭复合基质中,与对照相比,在W20和W30复合基质中叶片的叶绿素相对含量分别升高了5.8%和5.0%,净光合速率分别升高了19.2%和9.8%,蒸腾速率分别升高了23.8%和20.5%,气孔导度分别升高了16.0%和8.0%,胞间CO2浓度则分别降低了7.4%和5.0%;在W40复合基质中,叶片的叶绿素相对含量、净光合速率、蒸腾速率、气孔导度和水分利用效率均最低,较对照分别降低了6.7%、13.3%、2.4%、12.0%和11.0%,胞间CO2浓度最高,较对照升高了6.7%。4组稻壳炭-泥炭复合基质中,与对照相比,在R30和R40复合基质中叶片的叶绿素相对含量、净光合速率、蒸腾速率和气孔导度均较高,胞间CO2浓度和水分利用效率均较低,其中,叶绿素相对含量分别升高了6.5%和6.6%,净光合速率分别升高了8.7%和11.2%,蒸腾速率分别升高了21.1%和23.2%,气孔导度分别升高了8.0%和12.0%,胞间CO2浓度分别降低了4.8%和6.7%,水分利用效率分别降低了10.1%和9.8%。
表3 不同比例生物炭-泥炭复合基质中马缨杜鹃幼苗生长指标的差异
表4 不同比例生物炭-泥炭复合基质中马缨杜鹃幼苗叶片光合生理指标的差异
表5 不同比例生物炭-泥炭复合基质中马缨杜鹃幼苗叶片中矿质元素含量的差异
2.2.3 叶片中矿质元素含量的差异 由表5可见:在各组复合基质中叶片的N、P和Zn含量与对照无显著差异。在W30、W40和R40复合基质中,叶片的K含量较对照分别升高了17.6%、28.7%和17.9%,差异达显著水平;在其他复合基质中叶片的K含量与对照无显著差异。在W40复合基质中叶片的Mg和Na含量较对照分别升高了17.6%和25.7%,差异达显著水平;在其他复合基质中叶片的Mg和Na含量与对照无显著差异。在W20、W30、W40和R40复合基质中叶片的Fe含量较对照分别升高了20.4%、26.1%、40.1%和17.5%,差异达显著水平;在其他复合基质中叶片的Fe含量与对照无显著差异。在W30和W40以及4组稻壳炭-泥炭复合基质中叶片的Mn含量显著高于对照;其中,在W30和W40复合基质中叶片的Mn含量较对照分别提高了45.2%和62.5%,在4组稻壳炭-泥炭复合基质中叶片的Mn含量较对照提高了112.1%~229.5%。
综合比较结果显示:随复合基质中木炭体积分数的提高,叶片的N、P、K、Mg、Na、Fe、Mn和Zn含量总体上逐渐升高;其中,在W40复合基质中叶片的矿质元素含量总体上均最高。随复合基质中稻壳炭体积分数的提高,叶片的P、Mg、Fe和Mn含量逐渐升高,N、K、Na和Zn含量波动变化,其中N含量均低于对照;在R40复合基质中叶片的N含量最低,P、K、Mg、Na、Fe和Mn含量最高,Zn含量也较高。
2.3 马缨杜鹃幼苗生长和叶片光合生理指标与复合基质理化指标的相关性分析
马缨杜鹃幼苗生长指标和叶片光合生理指标与复合基质理化指标间的相关系数见表6。
由表6可见:幼苗的株高和地径与复合基质的各项理化指标均无显著相关性。幼苗的单株叶片数、单株叶面积、单株根干质量和单株叶干质量与复合基质的通气孔隙度、总孔隙度和Mn含量呈极显著(P<0.01)或显著(P<0.05)正相关,与复合基质的pH值、电导率和Na含量呈显著负相关;幼苗单株茎干质量仅与复合基质总孔隙度呈极显著正相关;叶片的叶绿素相对含量和净光合速率与复合基质的总孔隙度、持水孔隙度、K含量和Mn含量呈极显著或显著正相关,与复合基质的pH值、电导率和Na含量呈显著负相关;叶片的蒸腾速率和气孔导度与复合基质的通气孔隙度、总孔隙度、持水孔隙度、K含量和Mn含量呈极显著或显著正相关,与复合基质的pH值、电导率和Na含量呈显著负相关;叶片胞间CO2浓度与复合基质总孔隙度呈显著负相关,与复合基质的pH值和电导率呈显著正相关;叶片水分利用效率与复合基质的容重、总孔隙度、pH值、电导率、K含量、Na含量、Fe含量和Mn含量呈极显著或显著负相关。
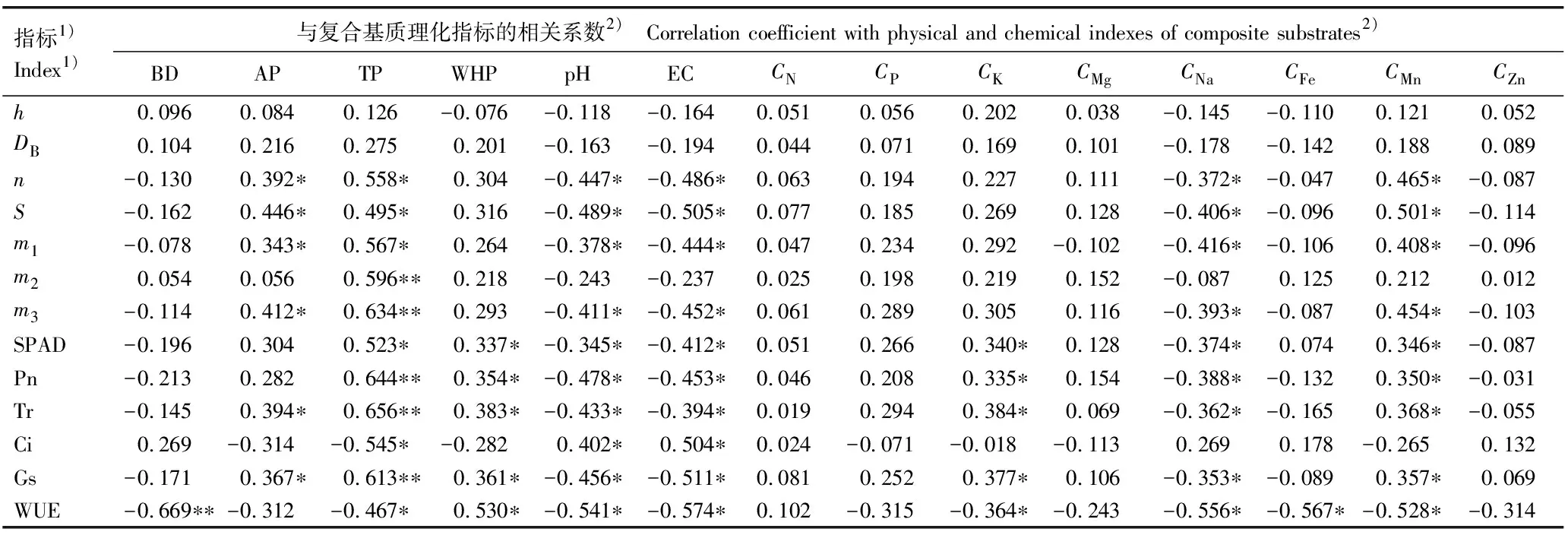
表6 马缨杜鹃幼苗生长和叶片光合生理指标与复合基质理化指标间的相关系数
3 讨论和结论
上述研究结果表明:在泥炭中添加不同比例的木炭和稻壳炭组成复合基质,对马缨杜鹃幼苗的干物质分配有不同程度的影响效应。总体上看,添加体积分数20%和30%木炭的W20和W30以及添加体积分数30%和40%稻壳炭的R30和R40这4组复合基质对马缨杜鹃幼苗的干质量有明显的提升作用;其中,与对照(泥炭)相比,W20复合基质可使幼苗单株的根、茎和叶的干质量显著增加,W30复合基质可使幼苗单株的茎和叶的干质量显著增加,R30复合基质可使幼苗单株叶干质量显著增加,R40复合基质可使幼苗单株的根和叶的干质量显著增加。这4组复合基质均具有较为适度的通气孔隙度和持水孔隙度,适合马缨杜鹃幼苗生长的pH值(pH 5.66~pH 6.11)和电导率(1.43~1.78 dS·m-1),以及充足的N、P、K和微量元素。而在添加体积分数40%木炭的W40复合基质中,马缨杜鹃幼苗单株的根和叶的干质量较对照分别降低了9.0%和7.8%,该复合基质的持水孔隙度已经低于盆栽观赏植物的栽培基质持水孔隙度参考标准下限(45%)[16],且酸碱度(pH 6.56)和电导率(2.15 dS·m-1)均超出杜鹃花属植物的最适酸碱度上限(pH 6.50)和电导率上限(2.00 dS·m-1)[17]。生物炭施用量过高可导致大量的可溶性盐基离子进入栽培基质中,影响其渗透势,进而限制植物根系对水分和养分的吸收,导致植物出现生理干旱[18],抑制植物生长。
Fornes等[19]以体积比75∶25的泥炭和森林废弃物生物炭组成复合基质,对金盏菊品种‘Nana Gitana’(Calendulaofficinalis‘Nana Gitana’)和碧冬茄品种‘Costa Rosa Vivo’(Petuniahybrida‘Costa Rosa Vivo’)进行栽培,这2种观赏植物均能良好生长;Blok等[20]发现,在栽培基质中添加体积分数20%生物炭对非洲菊(GerberajamesoniiBolus)没有产生任何植物毒性,也没有其他减缓生长的作用;Conversa等[21]认为,木炭可用于天竺葵(PelargoniumhortorumBailey)的栽培,但在栽培基质中木炭的体积分数不得超过30%;Bu等[22]的研究结果表明:在添加体积分数10%~40%稻壳炭的栽培基质中,羊踯躅〔Rhododendronmolle(Blume) G. Don〕1年生幼苗的地上部生物量和叶绿素含量均有所增加。这些研究结果均表明:当复合栽培基质中生物炭的体积分数为20%~40%时,观赏植物的生长没有受到明显的抑制作用,而适宜的生物炭比例可根据植物种类及其自身的生长特性确定。
与对照相比,在W20、W30、R30和R40复合基质中马缨杜鹃幼苗叶片的叶绿素相对含量升高了5.0%~6.6%,且净光合速率升高了8.7%~19.2%,反映了植物叶片合成有机质的能力提高,有助于其生物量的积累;叶片的叶绿素相对含量和净光合速率的增加很可能与其叶片的K、Fe和Mn含量明显升高有关,这3种矿质元素在叶绿素的生物合成中起重要作用[23],能够直接或间接影响植物的光合特性[24-25],马缨杜鹃幼苗叶片的叶绿素相对含量和净光合速率与复合基质的K和Mn含量显著正相关,也佐证了这一观点。然而,在W40复合基质中叶片的净光合速率降低了13.3%,净光合速率降低意味着可用于叶片生长的同化物减少[26],这也是该栽培条件下幼苗叶面积减少、叶干质量降低的主要原因。Fascella等[27]的研究结果表明:使用添加体积分数50%木炭的复合基质可使玫瑰(RosarugosaThunb.)叶片的叶绿素相对含量和净光合速率降低、叶面积减少,与叶片中Na元素的积累有关;而在本研究中,用W40复合基质栽植的马缨杜鹃幼苗叶片中Na含量较对照升高了25.7%,这可能是导致其叶片净光合速率降低的原因之一。从相关性分析结果也可见:马缨杜鹃幼苗叶片的叶绿素相对含量和净光合速率与栽培基质的电导率和Na含量呈显著负相关,符合“盐分(Na+或Cl-)引起的光合活性降低与植株叶片Na浓度增加相关”[28-29]的研究结论。
综上所述,不同比例的木炭和稻壳炭与泥炭组成复合基质,对马缨杜鹃幼苗生长和叶片光合生理指标和矿质元素吸收均有不同的影响效应。当木炭添加比例为体积分数20%~30%,稻壳炭添加比例为体积分数30%~40%时,马缨杜鹃幼苗叶片的叶绿素含量和光合能力提高,叶片干物质积累明显增加,其中,当木炭体积分数为20%或稻壳炭体积分数为40%时,马缨杜鹃幼苗的长势较佳、光合能力较强。然而,当复合基质中木炭的体积分数达到40%时,马缨杜鹃幼苗叶片的光合能力下降、叶面积和叶片干物质积累降低。另外,根据相关性分析结果,建议在杜鹃花属植物的生物炭复合栽培基质配制时,注意提升基质的通气孔隙度和总孔隙度、适度降低pH值和总盐度,以促进幼苗生长。
生物炭栽培基质对杜鹃花属植物生长和生理代谢的影响效应具有种间差异性,本研究的供试种类仅1种,涉及的生理指标也不全面,导致研究结果具有一定的局限性,因此,还需针对杜鹃花属多种植物开展生物炭复合基质的适应性研究,全面评估生物炭在杜鹃花属植物栽培过程中的应用效果;同时探索生物炭对植物生长和代谢影响的分子和生理机制,以期为生物炭复合栽培基质的推广应用奠定理论基础。
