温度反馈控制棘轮的最优控制*
2021-11-01刘天宇曹佳慧刘艳艳高天附郑志刚
刘天宇 曹佳慧 刘艳艳 高天附† 郑志刚
1) (沈阳师范大学物理科学与技术学院,沈阳 110034)
2) (华侨大学信息科学与工程学院,厦门 361021)
基于Cao 的误差棘轮模型,通过引入温度因子进一步对反馈棘轮实施控制.本文详细讨论了温度因子、温度相位差和温度频率对耦合布朗粒子定向输运的影响.研究发现,温度因子并不总是减小温度反馈棘轮的定向输运,这意味着在一定条件下温度因子还可以增强反馈棘轮的定向输运.此外,在小温度振幅范围内耦合粒子的质心平均速度和Pe 数随温度频率的变化都呈多峰结构.这一结果表明,合适的温度变化频率能够使反馈棘轮的定向输运获得多次的增强.本文所得结论不仅能够启发实验上通过选取合适的温度反馈信息来优化布朗棘轮的定向输运,还可为实验上的数据分析与处理特别是误差分析提供理论参考.
1 引言
生物分子马达是将化学能转化为机械能的酶蛋白大分子.实验研究表明,生物分子马达的定向运动充分参与了细胞内的物质输运过程,如胞膜穿梭、信号传导和病毒包装等[1−3].最新的实验研究还发现,肌球蛋白的突变与扩张性或肥厚性的心肌病、视网膜色素变性和失聪等症状有关;动力蛋白的失活是导致男性不育的原发性纤毛运动障碍的原因之一.随着生物分子马达的高持续免洗快速检测技术的日渐成熟,实验者们可以把标记物(如荧光探针)快速移动到探测窗口,这一操作省却了传统检测方法中标记物的清洗步骤,进而实现了生物分子马达的快速定量探测[4].由此可见,生物分子马达定向输运的研究对生物学、医学乃至未来分子机器的研发等都具有十分重要的意义[5−7].
理论上,生物分子马达的定向运动都可用棘轮模型进行研究[8−12].然而,其中大多数的研究模型都是不依赖于系统状态的开环棘轮.如Rosalie 和Fabrice[13]详细讨论了外势变化时的开环棘轮,模型为.本模型描述了经典过阻尼布朗粒子在外势、外力和高斯白噪声作用下的随机运动,发现粒子流随外势驱动速度v的增加能够呈现最大值.Pawel 和Felix[14]还研究了振荡周期势中的开环布朗棘轮,发现粒子流会随外力的增加而增大.通过对开环棘轮的不断探索,理论上人们又提出了依赖于棘轮状态的闭环棘轮[15−18].对于这种控制策略,Wang 和Bao[19]对反馈闪烁棘轮进行了研究,耦合粒子的运动方程为该模型主要研究受反馈控制z(t)、外势、耦合力及噪声作用下布朗马达的定向运动.这种闭环控制方式主要通过判断耦合粒子受到外势的平均作用而实现反馈控制.当耦合粒子受到的平均作用力大于零时,外势处于打开状态(z(t)1 );反之,外势处于关闭状态(z(t)0).Wang 和Bao[19]发现闪烁周期是影响粒子流大小的关键因素.此外,Feito 等[20]还详细研究了反馈摇摆棘轮,发现在外力振幅和频率的共同作用下粒子的定向输运速度能够呈现多峰结构.这种闭环棘轮的整流作用主要依赖于系统随时间的演化,也就是说外势的调制与系统的状态有关.如果知道任意时刻粒子的位置,根据上述开关的控制方式布朗粒子能够产生定向运动(几率流).实验上为了实现上述控制方法,研究者们通过监测溶液中胶体粒子的状态,进而可以对外势的打开或关闭实施反馈控制[21].然而,在大多数反馈棘轮的控制中,如文献[15−18]的反馈控制,并没有考虑外界因素对实验操控的影响,特别是实验上不可避免的随机误差、系统误差和人为误差等因素对实验操作的影响.由此,为了进一步研究误差因素对反馈棘轮的影响,Feito 和Cao[22]开创性地提出了误差概率p这一思想,并讨论了误差概率存在下反馈棘轮的输运行为.研究结果表明,粒子流会随误差概率的增加而减小.误差概率p的引入打破了人们对固有反馈棘轮的认识,为反馈控制棘轮模型的研究提供了新方向.
然而,在早期关于误差概率存在下反馈棘轮的研究中,Cao 的研究组仅讨论了误差概率p存在下棘轮的定向输运问题,有关误差概率究竟是由什么因素引起的,特别是p在实验中会受哪些实际因素的影响并不是十分清楚.因此,深入探索p的产生条件以及由此带来的影响是十分必要的.此外,实验研究还发现,生物分子马达在微管间的运动要受介质环境的作用,特别是温度会对分子马达的定向运动有较大影响.例如,Jayannavar 研究组[23]讨论了热浴环境中的反馈布朗粒子,发现热噪声强度是影响粒子流大小的主要因素.我们研究小组也讨论了双阱棘轮势中的反馈布朗粒子,发现在一定条件下粒子流呈周期性变化,且这种周期性会随热噪声强度的增大而变得越来越不明显[24].最近的实验研究表明,环境温度越高,粒子的运动越剧烈,实验上对粒子进行的反馈控制就越不容易实现,因此误差p的产生概率也会随之增大.由此可见,上述和温度相关的反馈控制棘轮的研究实例为分析不同温度下布朗马达的反馈操控问题提供了理论启发.进而深入讨论由温度变化带来的实验误差概率的影响更具实际意义.这一理论研究还可为反馈棘轮的实验操控、数据分析以及误差分析等提供强有力的理论支撑.
本文主要研究了不同临界温度、温度频率和相位差等作用下反馈棘轮的定向输运,讨论了这些因素对耦合布朗马达质心平均速度、质心扩散系数Deff及Pe数的影响.研究发现,合适的温度相位差、频率和临界温度都能促进温度反馈棘轮的定向输运.此外还发现,在温度振幅及温度频率的共同作用下,耦合布朗粒子的定向运动会出现多峰结构.本文所得结论不仅能为实验上的反馈操控提供理论依据,还可为后续的数据分析与处理、误差来源分析与计算提供理论启发.
2 温度反馈棘轮模型
本文主要研究受周期外势V(xi)、相互作用势U0(x1,x2)、反馈控制βi(t) 及噪声作用条件下的耦合布朗马达,其动力学行为可由过阻尼朗之万方程描述:

其中,t时刻两个耦合布朗粒子的位置分别为x1(t)和x2(t);γ是介质的阻尼系数;方程中(i1,2) 项表述的是耦合粒子受到的棘轮作用,V(xi) 是棘轮的周期外势,其表达式为

式中L为棘轮势的周期,V0是势垒高度;U0(x1,x2)是两个耦合布朗粒子的相互作用势,表述如下:

其中k为耦合强度,a是弹簧的自由长度.
在生物分子马达的实验中,发现驱动蛋白马达的步进具有持续性,并且马达的两个头部是交替步进的,进而相应的ATP 水解过程也会交替进行.例如,马达前导头的ATP 水解放热提高了其所在的环境温度,然后温度再传到后随头上并且能够继续进行ATP 水解.正是这种水解方式使结构相同的马达的两个头部间出现了温度差和相位差.这一现象为研究处于不同细胞环境温度下布朗马达的输运特性提供了参考.由此,方程(1)中高斯白噪声ξi(t) 满足如下统计特性[25]:
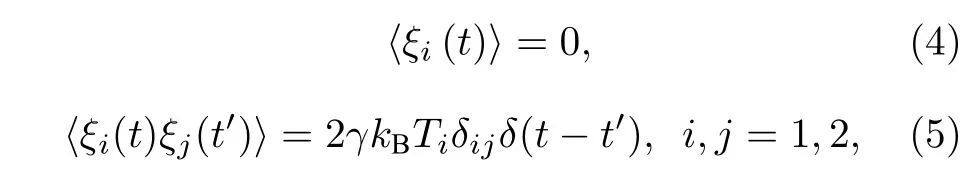
其中kB是玻尔兹曼常数,Ti为布朗马达头部所处的环境温度,其变化形式可写为
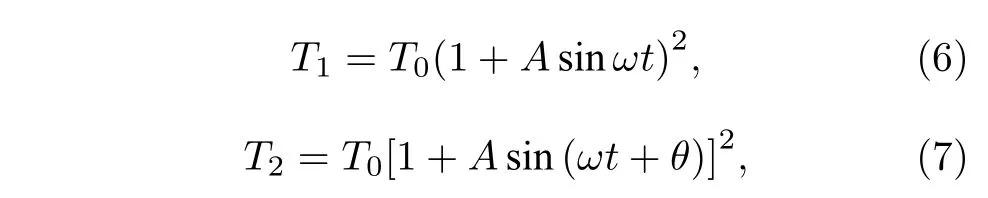
式中A和T0是温度变化的振幅,ω是温度变化的频率,θ是两个温度间的相位差.
方程(1)的βi(t) 是控制棘轮势打开或关闭的有效开关.一般情况下,当耦合布朗粒子受到棘轮的平均作用力大于零时,棘轮势处于打开状态(β(t)1);反之,当布朗粒子受到的平均作用力小于等于零时,棘轮势处于关闭状态(β(t)0).因此,通常的反馈棘轮的控制开关可表示为[16]
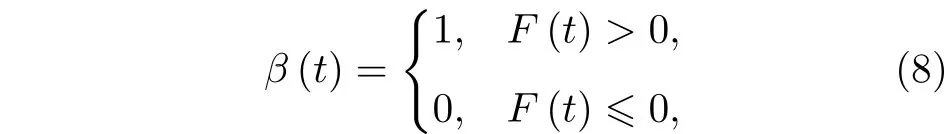
其中β(t) 是反馈棘轮的开关,F(t) 是耦合布朗粒子在棘轮势V(x) 中受到的平均作用力,即

然而,Cao 的研究组[22,26]在上述反馈棘轮基础上引入一个新的想法,即存在p的误差概率影响外势开关的判断,误差概率p的取值范围如下:

相应地,棘轮势的控制开关也将受到影响,不再是(8)式的形式.也就是说,当棘轮势的平均作用力大于零时,棘轮的控制开关将并不完全处于打开状态,即存在p的误差概率使棘轮处于关闭状态,则此时棘轮势处于打开状态的有效概率变为 (1−p) ;类似地,当棘轮势的平均作用力小于等于零时,棘轮也并不是完全处于关闭状态,即存在p的误差概率将使棘轮处于打开状态,则此时棘轮势处于打开状态的概率为p.根据上述分析,反馈棘轮处于打开状态的概率变为:

其中σ(t) 是粒子处于不同位置时棘轮打开的实际概率.
由于分子马达在生物体内水解 A TP 时会受环境温度的影响,温度越高,粒子运动越剧烈,对粒子进行的反馈控制便越不容易实现,相应地实验上也越容易产生随机误差.因此,本文提出利用温度因子αi进一步对反馈棘轮的开关实施控制,αi的表达式为
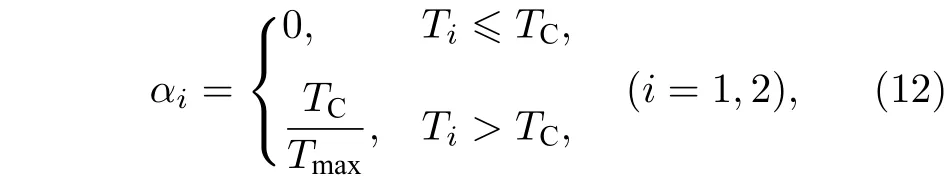
式中TC是临界温度.也就是说,当第i个布朗粒子的环境温度Ti小于等于临界温度TC时,棘轮的开关较易操作,且实验上不容易产生误差,此时温度因子几乎为0.然而,当环境温度一旦大于临界温度TC时,布朗粒子的运动较为剧烈,相对低温情况而言实验上棘轮开关的操作容易产生误差,且其温度因子可表为Tmax为环境温度Ti的最大值.
综合(11)式与(12)式的分析,朗之万方程(1)中总的有效控制开的概率可写成如下形式:

式中fi(t)−V ′(xi)(i1,2),αip表示的 是误差概率,即如果第i个布朗粒子受到外势的作用力大于零,第i个粒子棘轮势的状态有 (1−αip) 的概率是打开的,同时还伴随αip的概率是关闭的;类似地,如果第i个布朗粒子受到外势的作用小于等于零,那么棘轮势的状态有αip的概率是打开的,同时还有 (1−αip) 的概率棘轮处于关闭状态.因此无论粒子处于空间中的什么位置,即粒子受到棘轮的作用无论是大于零还是小于等于零,实际上每个粒子所处的棘轮势都有打开的概率.对于同一时刻、同一位置来说,每个粒子的反馈开关同时存在两种状态,即:正确的开、错误的关或者正确的关、错误的开.因此,方程(1)描述的是误差概率存在下受温度信息控制的反馈布朗棘轮模型.
为了研究耦合粒子的定向输运,本文采用耦合布朗粒子的质心平均速度来描述反馈棘轮的输运行为,即[27]:

其中,〈·〉表示系综平均,S为耦合布朗粒子的演化时间.由于布朗粒子在一段时间内会产生扩散现象,因此本文进一步引入质心扩散系数Deff来描述耦合粒子位移的涨落,表述如下[28]:

对于温度反馈棘轮来说,仅知道质心扩散系数还不能对反馈棘轮的定向输运进行全面了解,为此进一步引入Pe数,如下式[11,29,30]:

Pe数描述的是棘轮的平均速度与质心扩散系数的比,表示粒子的定向运动 〈Vcm〉 与随机扩散Deff之间的竞争关系.当Pe<1,定向输运只起很小的作用;当Pe>1 时,输运以漂移为主.因此反馈棘轮的Pe数越大,表明耦合布朗粒子的定向输运越强[13].
本文采用龙格-库塔算法对方程(1)进行数值模拟,研究布朗粒子的定向输运行为.文中模拟了8×103条轨道,每个轨道演化 1 04步,时间步长取10−3.无特别说明时,模型参数取γ1.0,V01.0,kB1.0,k300,L1.0,a0.3.为方便计算,所有物理量已经过无量纲化处理.
3 结果与讨论
3.1 温度相位差 θ 的影响
为了研究反馈棘轮在不同温度环境下的定向输运,本文详细讨论了温度棘轮的质心平均速度〈Vcm〉、质心扩散系数Deff及Pe数随棘轮不同参量的变化行为.
如图1(a)所示,本文详细讨论了不同温度因子αi(i1,2) 下质心平均速度 〈Vcm〉 随温度相位差θ的变化关系.由(6)式、(7)式和(12)式可知,对于固定的A,ω,T0和θ值,耦合粒子所处的环境温度Ti有相同的最大温度Tmax,进而有αi ∝TC.因此,对αi的研究相当于对临界温度Tc的研究.由(7)式可知,粒子的定向输运随相位差θ的增加呈周期性变化.为了便于分析,本文仅画出质心平均速度变化的一个周期图像,如图1(a)所示.有趣的是,当温度因子较小时,如αi0.2,平均速度随温度相位差θ的变化曲线会产生一个波谷;随着αi的增加,如αi0.3 时,〈Vcm〉 随θ的变化会变为一个波峰曲线;随着αi的继续增加,〈Vcm〉 的变化曲线出现了两个完整的波峰,且随αi的增加双峰结构越来越显著,曲线峰值对应θ的最优值也向两侧移动.产生上述现象的原因主要是在温度相位差θ和温度因子αi这两个和温度有关的因子相互协作和竞争作用下,耦合粒子的定向运动将被促进或抑制,因而温度反馈棘轮能够存在一个或两个极值.更为有趣的是,通过图1(a)可看出,质心平均速度〈Vcm〉关于θπ 对称.由此可见,在温度相位差θ和温度因子αi的共同作用下,可通过选择一个或多个合适的温度相位差增强反馈棘轮的定向输运.

图1 (a) 质 心平均速度 〈 Vcm〉,(b) 质心扩散系数 Deff 和(c) P e 数随温度相位差 θ 的变化曲线,其中 ω =0.1π,A=1.0 ,T0=0.7Fig.1.Curves of (a) the center-of-mass mean velocity 〈 Vcm〉,(b) the center-of-mass diffusion coefficient Deff and (c) Pe number varying with the phase different of temperature θ,where ω =0.1π,A=1.0 ,T0=0.7.
图1(b)给出温度因子αi不同时温度反馈棘轮的质心扩散系数Deff与温度相位差θ的变化关系.研究发现,Deff随θ的增加先减小后增大,而且在这个变化过程中Deff的最小值几乎都出现在θπ附近.产生这一结果的原因可由(7)式即T2T0[1+Asin(ωt+θ)]2进行分析.当θπ 时,温度T2随时间的变化能够取到最小值,这意味着此时耦合棘轮的温度最低,粒子越不容易发生扩散.因此,在相位差变化的一个周期内,温度棘轮的扩散会在θπ 处出现最小值.此外,与图1(a)类似,Deff曲线也是关于θπ 对称.同时,研究还发现,当θ固定时Deff并不随αi单调变化,说明温度因子αi对粒子的质心扩散也具有显著的影响.
图1(c)表示不同温度因子αi下耦合布朗粒子的Pe数和温度相位差θ的函数关系.可以发现Pe数同样不随θ单调变化,与图1(b)的变化趋势刚好相反,当θ≤π时,Pe数随着θ的增加而增大;当θ >π 时,Pe数随着θ的增加而减小.因此温度棘轮存在一个最优θ值能使耦合粒子的Pe数达到最大.根据(16)式,Pe数表示平均速度与扩散系数Deff的比.如图1(a)所示,在速度变化范围不太大的情况下,近似有,因此Pe数图像与Deff图像的变化行为正好相反.这意味着可通过改变温度相位差θ的方法来增强温度棘轮的定向输运特性.
3.2 临界温度 Tc 的影响
图1 的研究结果已表明,温度因子αi会对温度棘轮的定向输运产生影响.此外,根据(12)式可知,当环境温度的最大值Tmax一定时,αi与临界温度TC成正比.因此,通过研究不同临界温度TC时的输运情况可间接地讨论αi对温度反馈棘轮定向输运的影响.此外,由(6)式、(7)式和(12)式可知,对于固定的A,ω和θ值,不同的温度振幅T0将使耦合粒子所处的环境温度Ti有不同的最大值Tmax,因此图2 中每条曲线最右端TC变化范围的最大值(临界值)也是不同的.

图2 (a) 质心平均速度 〈 Vcm〉,(b) 质心扩散系数 Deff 和(c) P e 数随临界温度 TC 的变化曲线,其中 ω =0.1π,A=1.0,θ=0.2πFig.2.Curves of (a) the center-of-mass mean velocity 〈 Vcm〉,(b) the center-of-mass diffusion coefficient Deff and (c) Pe number varying with the critical temperature TC,where ω=0.1π,A=1.0,θ =0.2π.
图2(a)是不同温度振幅T0作用下临界温度TC对棘轮平均速度 〈Vcm〉 的影响.可以看出,随着TC的增加,温度棘轮的平均速度先减小后增加且在某一合适的TC处 〈Vcm〉 产生一个波谷.同时,对于同一T0来说,当TC取最小值0 和相应的最大值时,平均速度都能获得最大值.产生上述现象的原因可由(12)式和(13)式分析得到.当TC0 时,αi0,此时反馈棘轮不存在误差概率,即βi(t)1,进而反馈棘轮处于打开状态的概率最大.因此,反馈信息棘轮的平均速度能够产生最大值.同理,当TC取环境温度的最大值Tmax时,即αi1,这种条件下反馈棘轮处于错误打开的概率也是最大的,进而粒子的质心平均速度也能获得最大值.然而,当临界温度在两个极值区间变化时,粒子的平均速度必然会产生最小值,因此温度棘轮的输运会有波谷的产生.同时,研究还发现,随着T0的增加,温度反馈棘轮的整体输运也随之增大,且粒子达到波谷时所需的TC也随之增大.这是因为随着T0的增大,粒子所处溶液环境的整体温度升高,粒子运动更剧烈,进而粒子越过棘轮势形成定向运动的概率增加,促进了棘轮中粒子质心平均速度的增大.上述结果进一步表明,在一定条件下通过选取合适的临界温度TC(即实验上合适的误差概率的产生)还能够使温度反馈棘轮的质心平均速度达到极值.这一结论可为实验上反馈棘轮的数据分析特别是误差分析提供一定的理论启发.
图2(b)给出温度振幅T0不同时耦合布朗粒子的质心扩散系数Deff与临界温度TC的变化关系.研究发现,Deff随TC的变化先增加后减小,由此在Deff的变化过程中能够产生扩散的最大值.同时,与图2(a)比较可以发现,Deff峰值对应的TC值和〈Vcm〉 波谷对应的TC值大致相同.上述现象可由扩散公式(15)式进行解释.由于质心扩散系数反映的是质心涨落速度变化的物理量,从图2(a)已知质心平均速度的变化过程中将有极小值的产生,也就是说在整个 〈Vcm〉 从极大值到极小值或从极小值到极大值的变化过程中,质心速度的变化存在最大值,相应地,质心涨落的速度即Deff也必然存在最大值,且Deff产生极值处的TC与相应 〈Vcm〉 波谷处的TC大致相等.研究还发现,随着T0的升高耦合粒子的Deff也随之增大,且达到最大Deff值所对应的优化临界温度TC值右移(变大).产生这一现象的原因是环境温度越高,耦合粒子运动越剧烈,因此温度棘轮的扩散现象也越显著.
图2(c)给出不同温度振幅T0下耦合布朗粒子的Pe数随临界温度TC变化的函数关系.显而易见,图2(c)与图2(b)的曲线变化趋势正好相反.结果表明,温度棘轮的Pe数不随TC单调变化,而会出现最小值.产生这一结果主要是因为Pe数的变化将依赖于平均速度与扩散系数的变化情况.根据图2(a)与图2(b)的分析,当 〈Vcm〉 达到最小值时Deff近似达到最大值,因此这两个量的比值会有最小值的产生.特别地,随着T0的增加,发现温度反馈棘轮中粒子的Pe数也随之降低,这一现象说明实验上由于不可避免的误差概率的存在,利用较小的温度振幅T0便能有效促进耦合粒子的定向运动,从而达到增强温度棘轮定向输运的效果.
3.3 温度频率 ω 的影响
由于耦合布朗粒子的定向运动受温度的影响较大,因此本文进一步讨论了(6)式中不同温度振幅A下反馈棘轮的定向输运随温度频率ω的变化情况,如图3(a)所示.插图给出A1.0 时反馈棘轮的整体变化行为.结果表明,整体上温度棘轮的平均速度随温度变化频率ω的增加先减小后增加最后趋于稳定.这是因为当ω趋于0 时,在温度Ti变化的每个周期内耦合粒子接触环境温度的时间非常长,这样会导致耦合粒子越过势垒形成定向运动的概率增加,因此当ω→0 时平均速度较大;同理可分析,当ω在较小的范围内增加时,耦合粒子越过势垒形成定向运动的概率减小,相应地平均速度会变小.然而,当ω非常大时,粒子接触环境温度Ti的时间非常短暂且非常频繁,近似地可看成耦合粒子处于恒温环境中,这样会使耦合粒子越过势垒形成定向运动的概率达到最大,因此反馈棘轮的定向输运能够达到最大并趋于稳定.由上述讨论,可进一步分析当平均速度从最小值到最大值的变化过程中 〈Vcm〉 一定是增加的.因此,平均速度总体的变化趋势是先减小后增加最终趋于稳定.此外,更为有趣的是在小频率变化范围内,如图3(a)所示,温度棘轮的平均速度呈现多峰结构,且随温度频率的增加 〈Vcm〉 峰值变化的幅度逐渐减小.这种多峰结构的产生主要是温度振幅A和温度频率ω这两个温度因子之间相互协作和竞争的结果,使耦合布朗粒子的定向输运将被促进或抑制,因而温度反馈棘轮能够产生多个极值.
本文进一步研究了不同温度振幅A条件下温度反馈棘轮的扩散Deff随温度频率ω的变化关系,如图3(b)所示.研究结果表明,在图3(a)所示的小频率变化范围,随着温度变化频率ω的增加温度反馈棘轮的Deff仍呈多峰结构.这一现象的产生同样是由A和ω这两个温度因子之间的相互协作和竞争所导致.理论上同样可从扩散公式(15)式进行分析,质心扩散系数只和质心涨落的速度有关,因此一定条件下当平均速度呈现多峰结构时,扩散系数Deff也相应地随之产生多峰结构.也就是说,合适的温度变化频率ω也能使温度反馈棘轮的扩散达到最强.此外,研究还发现,对于同一ω值,Deff随温度振幅A的增加而增大,这是因为温度越高反馈棘轮的扩散越明显.

图3 (a) 质心平均速度 〈 Vcm〉,(b) 质心扩散系 数 Deff 和(c) P e 数随温 度频率 ω 的变化曲线,其 中 T0=0.7,θ=0.2π,αi=0.8Fig.3.Curve of (a) the center-of-mass mean velocity 〈 Vcm〉,(b) the center-of-mass diffusion coefficient Deff and (c) Pe number varying with the temperature frequency ω,where T0=0.7,θ =0.2π ,α i=0.8.
类似地,由上述分析同理可得,温度反馈棘轮的平均速度和Deff的多峰结构同样还会导致Pe数多峰结构的产生,如图3(c)所示.此外,研究还发现,对于同一ω值,Pe数随温度振幅A的增大而减小,这是因为在速度变化范围不太大的条件下,近似地 有Pe与Deff成 反比.由于Deff随A的增加 而增大,进而会导致Pe随A的增加而减小.如图1(b)与图1(c),图2(b)与图2(c)所示,类似地也存在同样的结论.此外,在小温度频率变化范围内反馈棘轮存在多个最优温度频率值ωopt能够使耦合粒子的Pe数达到极大值,这也意味着一定条件下温度信息棘轮的定向输运能够获得多次的增强.
4 总结
本文研究了处于不同温度下反馈棘轮中耦合布朗粒子的定向输运问题,详细讨论了温度相位差、临界温度及温度变化频率对耦合粒子的质心平均速度、扩散系数及Pe数的影响.研究发现,温度因子并不总是减小布朗粒子的定向输运,这也意味着在某一变化区间内温度因子(即临界温度TC)的存在还能增强反馈棘轮的定向输运.研究还发现,在小温度振幅范围内,粒子的输运随温度频率的变化呈现多峰结构,即存在多个温度变化频率能使耦合粒子的质心平均速度、质心扩散系数及Pe数都能达到极值,这也意味着合适的温度变化频率可以使反馈棘轮的定向输运获得多次增强.此外,本文研究还发现,温度反馈棘轮的质心平均速度随耦合强度k的增加能够出现极值,这也说明合适的耦合强度能够促进温度反馈棘轮的定向输运.由于篇幅所限,本文讨论的仅是两个耦合粒子的情形,且每个粒子的控制开关βi(t) 都不同.若系统的粒子数增加,实验操作上每个粒子的误差产生率会随之变大,因此温度误差棘轮的粒子流也会相应地减小.
此外,本文主要基于Cao 的开创性模型,即在误差棘轮的基础上通过提出温度因子αi进而对反馈棘轮实施控制的.实验上,在棘轮的具体操作过程中研究者可以通过监控布朗粒子的实时温度,并与环境的最大温度进行比较,进而可以在原有反馈棘轮的控制方式中加入对温度的判断过程.通过这种实验操控能够深入了解温度对误差棘轮输运的影响.然而,本文讨论的内容仅关注于分子马达所处的溶液环境即温度信息对误差概率p造成的影响.当然,在具体实验操作中还有其他因素,如外势结构,时间延迟等因素仍会对误差概率p(随机误差、系统误差等因素)造成一定影响,这也是今后误差棘轮研究的方向.本文所得结论不仅可以通过调节温度信息(如临界温度、温度振幅、频率等参量)来优化反馈棘轮的输运特性,更可为实验上的数据的分析与处理特别是误差来源的分析与计算提供理论启发.
