猴耳环嫩枝和叶组织的代谢物分析*
2021-10-28彭冠明林昌明司徒荣贵毛积鹏
黄 婷 彭冠明 林昌明 司徒荣贵 毛积鹏,2
(1. 台山市红岭种子园,广东 台山529223;2. 华南农业大学 林学与风景园林学院/广东省森林植物种质创新与利用重点实验室,广东 广州 510642)
猴耳环Pithecellobium clypearia为豆科Leguminosae 猴耳环属植物,广泛分布于我国南方各省,是一种重要的药用植物,具有抗炎、抗菌和抗病毒等多种功效[1-7]。猴耳环中化学成分种类繁多,主要有黄酮类、氨基酸类、生物碱类、糖及酚类等[8-9]。其中没食子酸、槲皮素和表没食子儿茶素没食子酸酯等黄酮类化合物为猴耳环代表性药用特征标志物[10-12]。猴耳环的入药部位主要以干燥的幼枝和叶片组织为主,但也有直接以茎干切片入药。前期相关学者利用高效液相色谱法和核磁共振波普等方法于猴耳环的主要药用部位嫩枝和叶中成功鉴定出没食子酸、槲皮素、槲皮苷、儿茶素和表没食子儿茶素等多种高生物活性的黄酮类化合物[13-14]。传统药用植物活性成分的鉴定具有单一性、分析通量有限,难以系统阐明药用植物的有效成分及其植株体内的分布情况。而超高效液相色谱法—串联质谱联用法(UPLC-MS/MS)作为目前代谢组学分析中的常用手段具有高灵敏、高通量、特异性强等特点,能高效快速地掌握植物内源性代谢物成分及其动态变化信息[15]。本研究利用UPLC-MS/MS 代谢组学技术对猴耳环的主要药用部位嫩枝和叶组织部位中的主要代谢物成分进行了测定,旨在为猴耳环代谢物成分鉴定及其分布特性提供重要信息,为猴耳环的质量控制和合理开发利用提供物质基础。
1 材料与方法
1.1 实验材料
猴耳环样本材料来自于广东省台山市红岭种子园。于2019 年9 月份取样,采取了2 a 生且生长旺盛的猴耳环嫩枝(NZ)和叶片(LY)组织部位,每个组织部位3 个生物学重复,适当剪碎后放入50 mL 无菌的离心管中随即放入液氮中,最后-80 ℃保存。
1.2 样本制备
将猴耳环各组织部位样本于冻干机(Scientz-100F)中真空冷冻干燥,干燥后利用研磨仪(MM 400,Retsch)研磨至粉末状。称取约100 mg 粉末溶解于1.2 mL 70%甲醇(色谱级,Merck)4 ℃ 条件下提取12 h,每2 h 涡旋震荡1次,以提高提取率。12 000 rpm 离心10 min 取上清液,用0.22 μm 微孔滤膜过滤样本,随后保存于进样品中用于UPLC-MS/MS 分析。
1.3 色谱和质谱数据采集
分别利用超高效液相色谱(SHIMADZU Nexera X2)和串联质谱(Applied Biosystem 4500 QTRAP)仪器系统采集猴耳环嫩枝和叶组织的色谱和质谱数据。色谱数据收集参数设置如下:色谱柱:Agilent SB-C18, 1.8 μm, 2.1 mm×100 mm;流动相:A 相为超纯水(加入0.1%的甲酸),B相为乙腈(色谱级,Merck);洗脱梯度:0.00 min,B 相比例为5%,9.00 min 内B 相比例线性增加到95%,并维持 1 min;10.00~11.10 min,B相比例降为5%,并以5%平衡至14 min;流速设置为0.35 mL/min,柱温40 ℃,进样量4 μL。
质谱数据采集条件为:电喷雾离子源(Electrospray Ionization,ESI) 温 度550 ℃, 质 谱 电压5 500 V(正模式)/-4 500 V(负模式),帘气(Curtain gas,CUR)25 psi,碰撞诱导电离(Collision-Activated Dissociation, CAD)参数设置为高。在三重四级杆中每个离子对是根据优化的簇电压(Declustering Potential,DP)和碰撞能进行扫描检测[16]。
1.4 代谢物定性与定量分析
基于迈维代谢公司的自建数据库MWDB(metware database),根据二级谱信息进行物质定性分析,分析时去除同位素信号、含K+离子、Na+离子和NH4+离的子重复信号。代谢物定量则是利用三重四级杆质谱的多反应检测模式进行。四级杆首先筛选目标物质的前体离子,前体离子经诱导电离后形成碎片离子,碎片离子通过三重四级杆过滤选择出特征碎片离子。随后进行峰面积积分,并对其中同一代谢物在不同样本中的质谱出峰进行积分校正[17]。
1.5 数据分析
利用MultiaQuant 软件和Analyst 1.6.3 软件处理质谱数据,并进行色谱峰的积分和校正工作,每个色谱峰的面积代表对应物质的相对含量,最后导出所有色谱峰面积积分数据保存。随后对样本数据进行主成分分析(Principal Component Analysis,PCA)和正交偏最小二乘法判别分析(Orthogonal Partial Least Squares—Discriminant Analysis,OPLS-DA),通过载荷图和变量重要性投影(Variable Importance in Projection,VIP)值大于等于1 来筛选差异代谢物。PCA 用R 软件中的Prcomp 函数对进行归一化处理后的数据进行分析。OPLS-DA 在原始数据进行log2 转换后,进行中心化处理,利用R 软件中的MetaboAnalystR 包的OPLSR.Anal 函数进行分析。
2 结果与分析
2.1 代谢物鉴定
利用UPLC-MS/MS 法在猴耳环的嫩枝和叶组织中共鉴定出712 种代谢物成分,其正离子模式下混合样本的质谱分析总离子流图如图1 所示,各峰分离效果良好表明仪器精密度良好结果可靠。其中氨基酸类物质78 种,酚酸类物质127 种,核苷酸及其衍生物44 种,类黄酮化合物148 种,生物碱类22 种,木脂素和香豆素类化合物15 种,有机酸类70 种,脂质类129 种,维生素类11 种,鞣质类9 种,萜类化合物9 种,糖及醇类40 种,其它类化合物10 种。148 种类黄酮代谢物种中,其中查尔酮5 种,二氢黄酮7 种,二氢黄酮醇6种,黄酮49 种,黄酮醇38 种,黄烷醇15 种,异黄酮18 种,原花青素10 种。

图1 正离子模式下混合样本的质谱分析总离子流图Fig.1 Total ions current of mass spectrometry analysis of mixed samples positive ion mode
2.2 多元统计分析
为了解猴耳环嫩枝和叶组织样本之间总体代谢差异和组内样本之间的变异度大小,对各样本数据进行PCA 分析(图2A)。结果表明第一主成分可解释53.96%的原始变量信息,第二和第三主成分共解释了28.55%(PC1:12.56%,PC2:15.99%)的原始变量信息。此外猴耳环嫩枝和叶组织样本具有一定的分离趋势,即存在一定组间差异,说明猴耳环嫩枝和叶中所含的代谢物成分存在一定差异。同时,同一组织部位的不同样本也相对分散,说明组内样本间也存在一定差异。PCA 是一种没有监督的分组方法,难以消除掉随机误差和组内误差,因此OPLS-DA 被用于更准确找到猴耳环嫩枝和叶组织部位中的差异代谢物。OPLS-DA 的S-plot 图如图2B 所示。结果显示猴耳环嫩枝和叶区分明显,且很大一部分代谢物的VIP 值大于或等于1,表明这些代谢物成分在猴耳环嫩枝和叶中的含量差异显著。

图2 猴耳环嫩枝和叶组织样本的多元统计分析Fig.2 Multivariate statistical analysis charts of Pithecellobium clypearia in different tissues
2.3 差异代谢物鉴定
根据单变量分析的差异倍数(Fold change)值和多元统计分析OPLS-DA 模型的VIP 值来筛选差异代谢物。选取了差异倍数大于等于2 或小于等于0.5,且VIP 值大于等于1 的代谢物认为差异显著。在鉴定到712 种代谢物中,393 种代谢物成分的含量在猴耳环的嫩枝和叶组织部位中没有显著差异。另外319 种代谢物成分则在猴耳环两组织部位中的含量差异显著。含量差异显著的代谢物中,162 种在猴耳环嫩枝组织部位中的含量显著高于叶组织,其中包含了19 种类黄酮化合物,如表1 所示。157种代谢物成分在猴耳环叶组织中的含量显著高于嫩枝,其中包含了43 种类黄酮化合物,如表2 所示。此外,比较分析发现高生物活性代谢物成分如柚皮素、芹菜素、槲皮素、二氢槲皮素、表儿茶素苷和表没食子儿茶素没食子酸酯等在猴耳环嫩枝中的含量显著高于叶组织。而表儿茶素、槲皮苷、杨梅苷和芦丁等高生物活性代谢成分则在猴耳环叶组织中的含量显著高于嫩枝。

表1 19 种在猴耳环嫩枝中含量显著高于叶组织的类黄酮化合物Table 1 The 19 flavonoids of Pithecellobium clypearia that the content in twig were higher than in leaves

表2 猴耳环叶组织中43 种含量显著高于嫩枝的类黄酮化合物Table 2 The 43 flavonoids of Pithecellobium clypearia that the content in leaves were higher than in twigs
2.4 差异代谢物分析
为了更方便和直观的了解差异显著代谢物的组织变化规律及其通路分布情况,我们对含量差异显著的代谢物进行了聚类和KEGG 富集分析。结果如图3 和图4 所示。聚类分析结果表明,大部分萜类化合物、鞣质类、核苷酸及其衍生物和黄酮类化合物在猴耳环叶组织中含量显著高于嫩枝。嫩枝中含量显著高于叶组织的代谢物成分主要为脂质类、酚酸类和氨基酸及其衍生物等化合物。KEGG 富集分析结果表明,含量差异显著的代谢物显著富集于缬氨酸、亮氨酸和异亮氨酸的生物合成通路,组氨酸、精氨酸和脯氨酸代谢通路,异黄酮生物合成通路,抗坏血酸和醛酸代谢通路以及ABC 转运蛋白通路。

图3 差异代谢物聚类热图Fig.3 Different metabolites cluster heat map
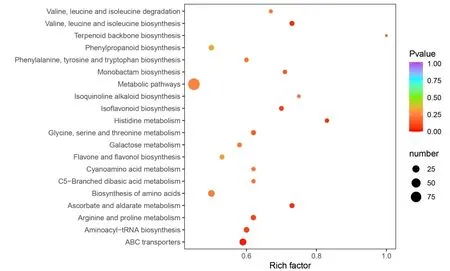
图4 差异代谢物KEGG 富集分析Fig.4 The KEGG enrichment analysis of differential metabolites
3 结论与讨论
本研究利用UPLC—MS/MS 代谢组学技术对猴耳环的主要药用部位嫩枝和叶的代谢物成分进行了鉴定与分析。嫩枝和叶组织中共鉴定出代谢物成分712 种。主要包括了氨基酸类、酚酸类、核苷酸及其衍生物和类黄酮等化合物。其中先前报道的槲皮素、二氢槲皮素、槲皮苷、表儿茶素、表儿茶素苷、杨梅苷和没食子儿茶素没食子酸酯等多种猴耳环药效成分均被检测到[1,3,11],而未检测到没食子酸单体,很可能是因为没食子酸常与鞣质和酯类等结合在一块[10]。综合表明UPLC—MS/MS 及其相关代谢组学技术适用于药用植物高生物活性物质的鉴定及新药源分子的挖掘。先前已有相关学者利用UHPLC-MS/MS、LS-MS/MS和UHPLC -QTOF/MSE 代谢组学技术分别对黄岑Scutellaria baicalensis的不同采收期材料[18]、新疆一支蒿Artemisia rupestris的不同组织部位[19]和文冠果Xanthoceras sorbifolium的不同组织部位[20]进行了代谢组学分析。多元统计、聚类和KEGG 富集分析结果显示,猴耳环嫩枝和叶组织存在显著代谢物成分含量差异。其中脂质类、酚酸类和氨基酸及其衍生物主要积累于猴耳环嫩枝部位,而萜类化合物、鞣质类和类黄酮化合物则主要积累于猴耳环的叶组织中。其中黄酮醇类化合物槲皮素在叶组织中的含量显著高于嫩枝,这与李雪玲等人[10]的研究结果相反,这很可能是因为不同产地,不同年龄的猴耳环组织部位代谢物成分及含量有一定差异[11]。本研究仅以台山市红岭种子园2 a 生的猴耳环嫩枝和叶为研究材料,结果具有一定的局限性。欲系统全面揭示猴耳环代谢物成分及特征活性物质在不同组织部位的分布特征及其相对含量的变化规律,为猴耳环有效部位的合理利用和开发提供重要的物质基础。后续应进一步对不同产地和年份的猴耳环的多个组织部位进行代谢组学研究。
