基于Pro-197-Ser突变的抗双氟磺草胺荠菜的快速检测
2021-10-27李琦,于金萍,郭文磊,刘亦学,张惟
李琦,于金萍,郭文磊,刘亦学,张惟


摘要:双氟磺草胺是防除小麦田荠菜(Capsella bursa-pastoris)等阔叶杂草的主要除草剂品种之一,目前在我国河南、河北、天津等地已有荠菜种群对其产生了抗药性。乙酰乳酸合成酶(ALS)基因第197位位点核苷酸突变是荠菜对双氟磺草胺产生抗药性的主要原因,通过在引物 D197F 序列的 3′ 端引入错配碱基,设计出一种衍生性酶切扩增多态性序列 (dCAPS) 分子标记方法,可用于第197位位点核苷酸突变的快速检测。抗药性及敏感荠菜的ALS基因扩增片段经限制性内切酶AvaⅡ酶切后表现出多态性,敏感荠菜产生了31、192 bp等2个片段;抗药性荠菜产生了31、192、223 bp等3个片段。该方法检测结果准确、可靠,与整株水平测定结果一致,并适用于第197位位点的任意氨基酸取代。这种dCAPS分子标记方法可对田间当季杂草进行检测,可为荠菜种群抗药性的快速检测与监测提供理论支持。
关键词:荠菜;双氟磺草胺;靶标抗药性;ALS基因;dCAPS
中图分类号:S481.4;S482.4文献标志码:A文章编号:1003-935X(2021)02-0014-06
Rapid Detection of Pro-197-Ser Mutation Endowing Target-Site
Resistance to Florasulam in Capsella bursa-pastorisLI Qi1,YU Jinping1,GUO Wenlei2,LIU Yixue1,ZHANG Wei1
(1.Institute of Plant Protection,Tianjin Academy of Agricultural Sciences,Tianjin 300381,China;
2.Plant Protection Research Institute,Guangdong Academy of Agricultural Sciences/Guangdong
Provincial Key Laboratory of High Technology for Plant Protection,Guangzhou 510640,China)Abstract:Florasulam is widely used in wheat fields to control Capsella bursa-pastoris and other broadleaf weeds. However,some C. bursa-pastoris populations have evolved resistance to florasulam in Henan,Hebei and Tianjin,China. The mutation occurring at ALS gene at 197 codon is a main mechanism causing florasulam resistance in C. bursa-pastoris. We introduced a mismatched base at 3′-end of the primer D197F and designed a dCAPS method to detect the mutation of ALS gene at 197 codon rapidly. The ALS fragments amplified between resistant and susceptible C. bursa-pastoris plants showed polymorphisms after digestion by the restriction enzyme AvaⅡ. The susceptible plants produced two fragments (31 bp and 192 bp). Resistant plants produced three fragments (31 bp,192 bp and 223 bp). Results were accurate and reliable,and were consistent with the whole-plant experiments. Moreover,the method was suitable for any mutations at ALS 197 codon by detecting resistance in season. This study provides a procedure to detect and monitor ALS herbicide-resistance in C. bursa-pastoris.
Key words:Capsella bursa-pastoris;florasulam;target resistance;ALS gene;dCAPS
荠菜(Capsella bursa-pastoris)是我国常见的阔叶杂草,广泛分布于长江流域、华北地区和西南地区作物田,主要危害油菜、稻茬麦、冬小麦等冬春作物。荠菜具有很强的耐低温及耐干旱能力,环境适应性强,在田间与作物竞争,严重影响着作物的质量与产量[1]。双氟磺草胺(florasulam)是由美国陶氏化学公司开发的三唑并嘧啶磺酰胺类除草剂,被广泛用于防除小麦田阔叶杂草,其作用机制是通過抑制乙酰乳酸合成酶(ALS)的活性,使杂草无法合成亮氨酸、异亮氨酸和缬氨酸这 3种氨基酸而死亡,是一种典型的乙酰乳酸合成酶抑制剂[2]。
由于具有高选择性、高活性、低毒、低残留的特点,乙酰乳酸合成酶抑制剂类除草剂被长期大量使用,导致杂草抗药性的发生非常严重,截至目前,全球已有165种杂草对 ALS类除草剂产生了抗药性[3-4]。除草剂作用靶标位点的氨基酸发生突变,影响除草剂与靶标酶的结合是抗药性产生最普遍的分子机制。目前,有报道显示,ALS基因中有8个氨基酸位点发生的27种氨基酸突变形式与抗药性相关[4]。根据报道,荠菜对ALS类除草剂产生抗药性的靶标机制包括Pro-197-His、Pro-197-Leu、Pro-197-Arg、Pro-197-Thr、Pro-197-Ser、Pro-197-Ala、 Trp-574-Leu等7种氨基酸突变形式[5-6]。
杂草对除草剂产生抗药性是一个不断发展和演化的过程[7],因此,加强对田间抗药性杂草的调查[8-9],对处于抗药初期的杂草进行准确快速的检测,才能最大程度地减少抗药性杂草对农业生产的危害。目前,常用于杂草抗药性检测的方法主要有温室整株植物鉴定法[10]、抗性当季快速检测(resistance in season quick test,RISQ) 法[11]、种子培养皿法[12]、靶标酶基因测序法[13]、酶切扩增多态性检测法[14]等。其中,温室整株植物鉴定法、RISQ法与种子培养皿法简单易行,结果直观,对试验仪器要求低,可以针对不同杂草的各种抗药性机制进行检测,但是检测效率低、速度慢,需要占用大量的试验空间,不适宜于大量样品同时检测;靶标酶基因测序法的结果准确,已广泛应用于多种除草剂抗药性检测中,但是该方法对试验仪器要求高,试验结果不够直观,须要进行进一步的生物学分析;酶切扩增多态性检测法(derived cleaved amplified polymorphic sequence,dCAPS) 通过在扩增引物中引入错配碱基提供限制性内切酶的识别位点,根据限制性内切酶产生不同长度的酶切片段,判断特定突变的基因和正常的基因,其操作简单、检测方便、结果直观、成本低、周期短,可以对大量样品同时进行检测,但其只能检测已知的单一突变位点。本研究针对在荠菜中最常见的ALS基因197位突变位点设计了 dCAPS 标记方法,以期在荠菜抗药性愈发严重的情况下,为其快速检测提供理论依据。
1材料与方法
1.1供试材料
杂草种子:2018年6月,从天津市蓟州区小麦田采集荠菜种群TJ07;另从天津市农业科学院核心区内非耕地处(该地未使用过任何除草剂)采集1个种群作为敏感对照,编号为TJ12。荠菜种群采集信息见表1。
1.2主要仪器
ASS-4型化学农药实验自动控制喷洒系统,购自北京盛恒天宝科技有限公司;5804R型台式高速冷冻离心机,购自德国 Eppendorf 公司;T100型PCR仪,购自美国Bio-Rad公司;DYY-8C型电泳仪,购自北京六一生物科技有限公司;HDL洁表1荠菜种群采集地点
Table 1Collection location of C. bursa-pastoris populations
种群编号采样点经纬度TJ07天津市蓟州区侯家营镇韩庄子村39°52′36″N,117°16′3″ETJ12天津市农业科学院核心区39°6′14″N,117°3′32″E
净工作台,购自北京东联哈尔仪器制造有限公司;RXZ-280C植物培养箱,购自宁波江南儀器厂;GL2200凝胶成像系统,购自美国柯达公司。
1.3试验方法
1.3.1荠菜对双氟磺草胺的抗药性水平测定采用整株水平测定法测定荠菜对双氟磺草胺的抗药性[15]。荠菜种子催芽方法同文献[16]。将15粒露白的荠菜种子均匀地播种于直径为12 cm的塑料盆内,置于可控温室中培养(自然光照,温度为20~35 ℃,相对湿度为50%~70%)。生长至2叶期时间苗,每盆留长势基本一致的荠菜10株。待长至3~4叶期,用ASS-4型自动农药定量喷雾系统进行茎叶喷雾。喷雾压力为0.275 MPa,喷液量为450 L/hm2。设定双氟磺草胺的处理剂量(有效成分用量):抗药性(R)种群的为0、0.06、030、1.50、7.50、37.50、187.50 g/hm2;敏感(S)种群的为0、0.012、0.060、0.300、1.500、7.500、37.500 g/hm2。每个处理重复3次,试验共重复2次。喷药后21 d,剪取荠菜地上部分,于恒温干燥箱内75 ℃烘干72 h,称量干重并记录。
1.3.2荠菜ALS基因片段克隆待荠菜生长至4叶期后,采用植物基因组DNA提取试剂盒(北京全式金生物技术有限公司),从敏感、抗药性种群中分别随机选取5株荠菜以提取基因组DNA。采用已报道的2对引物序列[Forward 1 (5′-ATTCGTCTCCCGATTTGC-3′)/Reverse 1 (5′-GCCCGACACCAGTGCTTAT-3′)和Forward 2 (5′-GGTATCCCTGTTGCGAGTA-3′) Reverse 2(5′-GCATACAAAGACCGTTTA-3′)]对荠菜ALS基因进行扩增[16]。引物由天津金唯智生物科技有限公司合成。PCR反应体系包括1 μL基因组DNA模板、1 μL 正向引物(10 μmol/L),1 μL反向引物(10 μmol/L)、12.5 μL 10×Easy Taq SuperMix(北京全式金生物技术有限公司),加双蒸水至总体积为25 μL。反应条件:94 ℃预变性5 min;94 ℃变性30 s,56 ℃退火40 s,72 ℃延伸90 s,34个循环;72 ℃终延伸10 min。反应结束后,取5 μL扩增产物用1%琼脂糖凝胶进行电泳检测,并将含目的条带的扩增产物送天津金唯智生物科技有限公司测序。将测序所得序列与NCBI上的荠菜ALS基因进行比对,用软件DNAMAN 6.0.3对扩增出的敏感、抗药性荠菜的ALS基因进行比对分析。
1.3.3ALS 第197位位点 dCAPS 检测分析用dCAPS 在线设计工具 dCAPS Finder 2.0[17]及引物设计软件 Primer Premier 5.0,通过在荠菜ALS基因197位点附近人为引入AvaⅡ酶切位点,使抗药性、敏感荠菜的ALS基因片段经限制性内切酶AvaⅡ酶切后表现出多态性,设计出1对引物 D197F 和 D197R(表2)。敏感荠菜ALS基因片段被完全酶切,含有31、192 bp等2个片段;纯合突变抗药性荠菜无法被酶切,只有223 bp 1个片段;杂合突变抗药性荠菜同时含有31、192、223 bp等3个片段。PCR反应体系同“1.3.2”节。反应条件:94 ℃ 预变性5 min;94 ℃变性30 s,56 ℃退火 40 s,72 ℃延伸30 s,38个循环;72 ℃终延伸 10 min。酶切反应体系包括 4 μL PCR 产物、1 μL限制性内切酶 (10 U/μL)、2 μL 10 × Buffer,加水至总体积为20 μL。限制性内切酶AvaⅡ反应温度为37 ℃,酶切时间为30 min。酶切反应完成后,加入2 μL 10 × Loading Buffer混匀,取10 μL至 2.5%琼脂糖凝胶中,在1×TAE电泳缓冲液中120 V电压下电泳25 min。结束后通过紫外凝胶成像系统观察并拍照。
1.3.4数据处理试验数据采用双逻辑非线性回归模型y=C+{(D-C)/[1+(x/GR50) b]}拟合剂量反应曲线,计算抑制50%荠菜生长所需的除草剂剂量(GR50)。式中:y为除草剂某一剂量下荠菜干重相对于空白对照的百分比;x为除草剂表2用于荠菜 ALS 第197位位点 dCAPS 分析的引物信息
Table 2Primers used for dCAPS in ALS 197 codon of C. bursa-pastoris
引物序列(5′→3′)目标氨基酸位点退火温度
(℃)酶识别位点预计酶切后片段大小(bp)野生型突变型D197FGTTCCTCTTGTAGCAATCACAGGACAGGTCPro197,CCT56AvaⅡ,GGWCC192,31223D197RGTTGCTGGATATCTTTAGGAATA注:引物中加粗及下划线显示的碱基表示人为设计的错配以引入AvaⅡ切酶位点;限制性内切酶位点包含的碱基在引物和目标氨基酸位点中用下划线标出。
剂量;C为剂量反应下限;D为剂量反应上限;b为斜率。
抗性倍数 (resistance index,RI)计算公式为RI=抗性种群的GR50/敏感种群的GR50。参考Beckie等的方法[18]对抗性倍数进行分级:敏感为RI<2,低水平抗性为2≤RI<5,中水平抗性为5≤RI≤10,高水平抗性为RI>10。
2结果与分析
2.1荠菜对双氟磺草胺的抗药性水平
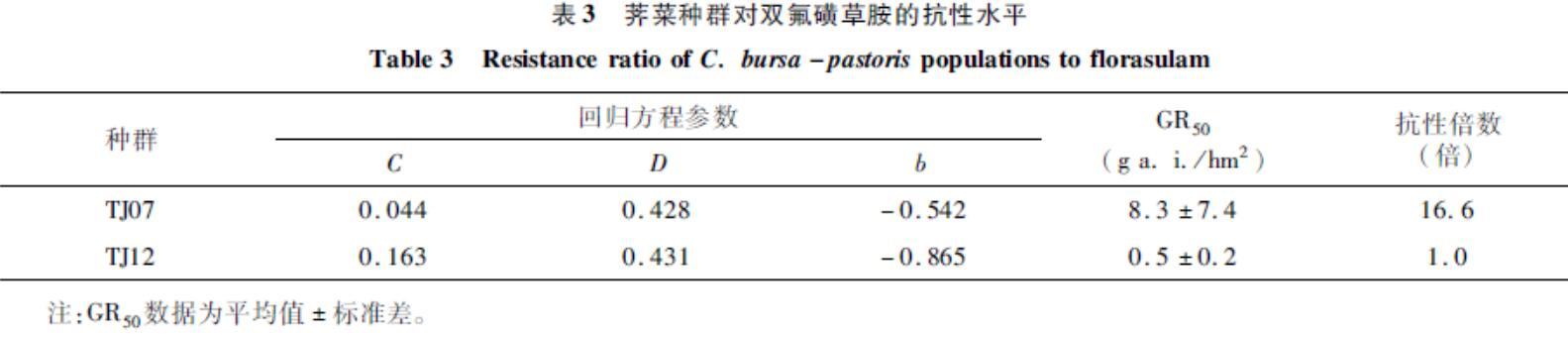
整株水平测定结果(表3)表明,荠菜种群TJ07对双氟磺草胺产生了高水平抗药性,GR50为8.3 g a.i./hm2,与敏感种群TJ12相比抗性倍数为16.6。表3荠菜种群对双氟磺草胺的抗性水平
Table 3Resistance ratio of C. bursa-pastoris populations to florasulam
种群回归方程参数CDbGR50
(g a. i./hm2)抗性倍数
(倍)TJ070.0440.428-0.5428.3±7.416.6TJ120.1630.431-0.8650.5±0.21.0注:GR50数据为平均值±标准差。
2.2ALS基因片段测序
测序结果显示,扩增所得ALS基因片段长度为1 738 bp,与NCBI中荠菜ALS基因序列(HQ880660.1)同源性达 99%以上,并且包含所有已经报道的8个抗性氨基酸突变位点。将抗药性种群和敏感种群的ALS基因序列进行比对,发现抗药性种群TJ07的5株被测植株中均存在 Pro-197-Ser (CCT-TCT)氨基酸突变形式,敏感种群TJ12的5株被测植株中均未发现已证实的ALS基因突变(图1)。
2.3荠菜ALS基因第197位位点 dCAPS 检测分析
本研究针对敏感荠菜ALS基因第197位位点设计酶切位点,使扩增的敏感荠菜ALS基因片段能够被限制性内切酶识别并酶切。在引物D197F中引入1个错配碱基,使得敏感ALS基因在第197位位点生成了1个AvaⅡ的酶切位点(GGWCC)。
以“2.2”节中荠菜基因组DNA为模板,采用引物D197F/D197R对5株抗药性植株、5株敏感植株均扩增出一条223 bp的目的片段,与预期一致。用限制性内切酶AvaⅡ对PCR产物进行酶切,通过观察电泳图谱,5株敏感植株产生了31、192 bp 等2个条带,5株抗药性植株产生了31、192、223 bp 等3个条带,但因 31 bp 的条带较小,无法在电泳图谱上显示(图2)。
3讨论与结论
杂草对除草剂产生抗药性是一个不断发展的过程。据报道,ALS 抑制剂类除草剂在同一地块连续重复使用3年以上,就可导致該地块杂草产生抗药性[19]。双氟磺草胺是应用广泛的一种麦田茎叶处理剂,若因杂草抗药性导致防效差,会增加作物减产风险。因此,加强对田间抗药性杂草的发生区域调查及监测在抗药性杂草的治理中变得尤为重要。
目前常用的抗药性杂草检测方法,根据杂草的检测层次可以分为表型检测、生理生化检测、分子检测等[20]。表型检测一般是指以生存率、死亡率、生物量等检测指标去比较除草剂对抗敏植株效果的生物测定方法。邹红梅等采用琼脂法实现了菵草对3种乙酰辅酶A羧化酶(ACCase)抑制剂的快速检测[21],该类方法简单易行,可检测不同抗药性机制,但需要较大空间,试验周期偏长,费时费力,人为因素影响较大。生理生化检测一般是指通过测定除草剂靶标酶及相关代谢酶的活性,判断是否存在抗药性,该方法对试验技术及试验环境要求较高。分子检测是基于核酸水平,对目的植物基因进行分析的一种检测方法,该类方法较多,不同的方法之间差别较大,对试验技术、试验设备以及试验成本的要求也不同。崔海兰等通过对靶标酶基因的扩增及测序,建立了猪殃殃靶标抗药性快速检测方法[22],但该方法在2次测序反应以后,需要专业软件对测序结果进行生物分析,对试验人员要求较高,不适宜大规模推广。
ALS基因发生核苷酸突变已被大量研究证明是杂草对ALS抑制剂类除草剂产生抗药性的主要机制之一[3]。本研究通过引物D197F的错配碱基引入内切酶AvaⅡ (GGWCC)酶切位点,采用dCAPS方法可检测荠菜中ALS基因第197位的Pro-197-Ser (CCT-TCT)氨基酸突变形式。该方法对5株抗药性植株和5株敏感植株的测定结果具有良好的一致性,说明本研究设计的 dCAPS分析方法可对双氟磺草胺抗药性荠菜Pro-197-Ser突变进行准确可靠的检测。目前,荠菜被报道的ALS197位点突变形式主要有以下几种:Pro-197-His (CCT-CAT)、Pro-197-Leu (CCT-CTT)、Pro-197-Arg (CCT-CGT)、Pro-197-Thr (CCT-ACT)、Pro-197-Ser (CCT-TCT)、Pro-197-Ala (CCT-GCT)[23],虽然发生突变的碱基不同,但均为第197位密码子的前2位碱基,而这2个碱基均包含在AvaⅡ内切酶的识别位点上,因此,本方法可以检测到第197位位点多种形式的核苷酸变化。本研究中所检测的TJ07种群,所有突变均为杂合,这可能是因为荠菜中含有多个ALS基因拷贝,使用dCAPS方法进行检测,结果显示的杂合是部分同源染色体杂合性,不是等位基因杂合[21]。
加强对田间抗药性杂草的发生区域调查及监测,有利于在杂草抗药性发展初期进行防治,最大程度减少抗药性杂草对农业造成的危害。本研究构建了荠菜ALS基因第197位位点突变的dCAPS检测方法,该方法操作简单、结果直观,可对田间当季杂草进行检测。由于该方法只能对已知的单一靶标突变位点进行检测,对于未知的突变位点以及非靶标抗药性须要结合其他检测方法鉴定。
参考文献:
[1]池吉平. 农田杂草危害及防除[J]. 现代农业,2011(7):24.
[2]Mccourt J A,Pang S S,King-scott J,et al. Herbicide binding sites revealed in the structure of plant acetohydroxyacid synthase[J]. Proc Natl Acad Sci USA,2006,103(3):569-573.
[3]Powles S B,Yu Q. Evolution in action:plants resistant to herbicides[J]. Annu Rev Plant Biol,2010,61:317-347.
[4]Heap I. The international survey of herbicide resistant weeds[Z/OL]. [2020-11-16]. http://www.weedscience.org/Summary/SOASummary.aspx.
[5]Jin T,Liu J L,Huan Z B,et al. Molecular basis for resistance to tribenuron in shepherds purse[Capsella bursa-pastoris (L.) Medik.][J]. Pesticide Biochemistry and Physiology,2011,100(2):160-164.
[6]Cui H L,Li X J,Wang G,et al. Acetolactate synthase proline (197) mutations confer tribenuron-methyl resistance in Capsella bursa-pastoris populations from China [J]. Pesticide Biochemistry and Physiology,2012,102(3):229-232.
[7]吉美静,黄兆峰,崔海兰,等. 一个抗咪唑乙烟酸长芒苋种群的发现[J]. 杂草学报,2020,38(3):23-27.
[8]谷涛,李永丰,杨霞,等. 4种稗属杂草对二氯喹啉酸等常用除稗剂的抗药性[J]. 杂草学报,2019,37(3):29-35.
[9]徐伟东,陆强,黎菊,等. 嘉兴市稻田主要禾本科杂草发生规律及其抗性表现与防治策略研究[J]. 杂草学报,2019,37(2):10-15.
[10]Ryan G. Resistance of common groundsel to simazine and atrazine[J]. Weed Science,1970,18(5):614-616.
[11]Kaundun S S,Hutchings S J,Dale R P,et al. Syngenta ‘RISQ test:a novel in-season method for detecting resistance to post-emergence ACCase and ALS inhibitor herbicides in grass weeds[J]. Weed Research,2011,51(3):284-293.
[12]Tal A,Kotoula-Syka E,Rubin B. Seed-bioassay to detect grass weeds resistant to acetyl coenzyme A carboxylase inhibiting herbicides[J]. Crop Protection,2000,19(7):467-472.
[13]Zhang L L,Guo W L,Li Q,et al. Tribenuron-methyl resistance and mutation diversity of the AHAS gene in shepherds purse [Capsella bursa-pastoris (L.) Medik.]in Henan Province,China[J]. Pesticide Biochemistry and Physiology,2017,143:239-245.
[14]Délye C,Pernin F,Michel S. ‘Universal PCR assays detecting mutations in acetyl-coenzyme A carboxylase or acetolactate synthase that endow herbicide resistance in grass weeds[J]. Weed Research,2011,51(4):353-362.
[15]Ryan G F. Resistance of common groundsel to simazine and atrazine[J]. Weed Sci,1970,18(5):614-616.
[16]張乐乐,郭文磊,李伟,等. 荠菜对乙酰乳酸合成酶抑制剂类除草剂的抗性水平及其分子机制[J]. 农药学学报,2016,18(6):717-723.
[17]Neff M M,Turk E,Kalishman M. Web-based primer design for single nucleotide polymorphism analysis[J]. Trends Genet,2002,18(12):613-615.
[18]Beckie H J,Tardif F J. Herbicide cross resistance in weeds[J]. Crop Prot,2012,35:15-28.
[19]Rubin B. Herbicide-resistant weeds-the inevitable phenomenon:mechanisms,distribution and significance[J]. Z Pflanzenkr Pflanzenschutz,1996,15:17-32.
[20]R4P Network. Trends and challenges in pesticide resistance detection[J]. Trends Plant Sci,2016,21(10):834-853.
[21]鄒红梅,李香菊,崔海兰,等. 菵草对 3 种 ACCase 抑制剂抗性的琼脂快速检测方法建立[J]. 杂草科学,2015,33(3):24-28.
[22]崔海兰,王藏月,徐林林,等. 猪殃殃对 AHAS 抑制剂靶标抗性的快速分子检测[J]. 植物保护学报,2016,43(6):1049-1054.
[23]张乐乐. 麦田杂草荠菜(Capsella bursa-pastoris)对苯磺隆抗性分子机制的研究[D]. 泰安:山东农业大学,2018.
收稿日期:2020-11-30
基金项目:天津市农业科学院植物保护研究所所长创新基金(编号:20190001S)。
作者简介:李琦(1990—),男,山东济宁人,博士,助理研究员,主要从事农田杂草防除及杂草抗性机理研究。E-mail:liqi0309@hotmail.com。
通信作者:刘亦学,硕士,副研究员,主要从事除草剂应用技术研究。E-mail:liuyixue113@163.com。
