上海浦东东滩湿地斑嘴鸭和绿头鸭的越冬期空间行为与栖息地利用
2021-10-23叶思嘉黄美玲金惠宇薄顺奇罗梓菁顾建明王天厚王正寰
叶思嘉,马 硕,周 锋,韦 旭,岳 衢,黄美玲,吴 迪,金惠宇,薄顺奇,袁 晓,罗梓菁,顾建明,王天厚,王正寰,5
(1. 华东师范大学 生命科学学院,上海 200241; 2. 上海市金山区林业站,上海 201500; 3. 上海市绿化和市容管理局,上海 200041; 4. 上海市浦东新区林业站,上海 201200; 5. 华东师范大学 转化科学与技术联合研究院,上海 200241)
雁鸭类是上海地区越冬水鸟的代表类群之一,也是禽流感病毒最主要的野生动物宿主类群之一.了解越冬季鸭类的空间行为和栖息地利用则可为开展水鸟保护和提升城市生态安全提供依据.上海位于长江口区域,有着大面积的湿地分布,是迁徙水鸟重要的栖息地,迄今记录到水鸟达163种[1],雁鸭类是其中越冬水鸟的代表类群之一[2-3].近年来以禽流感为代表的野生动物疫源疫病给人类正常生产生活带来很大影响[4-5].而迁徙性水鸟,尤其是雁鸭类一直以来都是禽流感病毒重要的自然宿主[6].雁鸭类也是上海地区主要的禽流感病毒野生宿主[7].雁鸭类在上海地区传统的分布区域——沿海滩涂又面临着围垦和人类活动加剧的局面[8],这些既影响雁鸭类在上海地区的栖息,也增加了禽流感病毒向人类环境扩散的风险.因此加强上海地区雁鸭类空间活动范围和规律的研究,既是迁徙水鸟保护生物学的需要,更是上海地区生态安全建设的需要.
自上世纪80年代以来,浦东东滩(原南汇东滩)经历了多次大规模围垦[9].围垦区的鸭科鸟类种群数量高于非围垦区域鸭科鸟类[10],这为我们了解人类活动干扰背景下的雁鸭类栖息地利用和空间行为模式提供了研究机会.已有的研究显示,越冬季雁鸭类会利用鱼塘、河道、农田栖息和觅食,但是回避人类活动的区域[12].另一方面,雁鸭类对于围垦地区各种栖息地的利用又存在明显的节律性[13].这与雁鸭类在栖息地偏好和反捕食策略之间的权衡有关[14].然而,以上研究结果均是基于野外观察数据获得的.由于观测能力的限制,制约了精确的、连续的鸟类空间行为数据的可获得性,最终影响分析结果的全面性.
近年来,随着无线电和GPS(Global Positioning System)背负式跟踪设备在鸟类行为学研究中的应用[15-16],结合遥感和地理信息系统(Geographic Information System, GIS)技术[17-19],人们已经可以在更大空间尺度和更精细的时间尺度上对鸟类空间行为进行连续观测,从而极大地推动了鸟类空间行为的研究[20-22].目前这一技术已经广泛运用于鸟类的迁徙行为和栖息地利用的研究中[23-29],这使得我们通过使用GPS追踪技术研究雁鸭类在上海地区越冬季的栖息地利用模式成为可能.
因此,本研究以上海浦东东滩冬季最常见的斑嘴鸭(Anaspoecilorhyncha)和绿头鸭(A.platyrhynchos)两种鸭属鸟类为研究对象,采用GPS定位跟踪技术就以下3个方面展开研究: 1) 鸭科鸟类的迁徙行为;2) 浦东东滩地区鸭科鸟类的活动范围和活动节律;3) 两种鸭物种的栖息地利用特征以及对不同类型生境的时间利用模式.
1 研究方法
1.1 研究地点
上海浦东东滩为长江口南岸潮滩,与之相距10 km的长江河口心滩为九段沙[30].浦东东滩南北长40.3 km,总面积约122.5 km2[31],主要植被为海三棱藨草和芦苇,湿地资源丰富,是上海地区重要的水鸟栖息地,也是上海市第一个野生动物禁猎区所在地.同时浦东东滩又存在大面积围垦,由于农业生产和新城区建设,该区域人为干扰较重[8-9],是典型的人类和野生水鸟活动交错区.
1.2 GPS追踪器的佩戴、设置与数据回收
于2017年1月18日在浦东东滩世纪塘鸟类禽流感检测站(31°0′11.90″N,121°55′28.23″E)选择了4只绿头鸭(Anasplatyrhynchos)和5只斑嘴鸭(A.poecilorhyncha),共9只健康且体质量(m个体)大于600 g的个体,安装了背负式GPS卫星追踪器(HQBG2715S,环球信士),追踪器单个质量(m追踪器)17 g,所有个体均达到佩戴设备小于体重3%的标准[16].所有研究个体的基本情况详见表1.设定GPS追踪器数据周期为每小时1个位点.此外,本研究使用的GPS追踪器内置运动探头,可以记录一个计时周期内(1 h)的振荡次数以反映最终动物在该时段内的活动量.监测个体的GPS位点数据通过Tracker Client 2.0[32]回收和管理.

表1 监测个体基本信息
全部野外工作经上海市林业局和浦东新区林业站批准,程序和操作符合上海市林业局野生动物保护的相关规定.
1.3 监测个体越冬期活动范围计算
因为斑嘴鸭BZ004和绿头鸭LT004个体有效位点数过少(<400),同时绿头鸭LT002整个越冬期主要在嵊泗列岛活动,因此不再对这3个个体进行后续的数据分析.
根据GPS追踪器传回的有效GPS位点对被追踪个体的活动路径和活动范围进行跟踪.关于监测个体越冬期数据的认定,分为3种情况进行: (1) 对于监测到迁徙行为的个体,以最早发生迁徙行为(向北方迁徙)的时间为准,将从开始GPS追踪到该时间点之前的时间段内获得的位点数据作为被追踪个体在上海期间的越冬期数据;(2) 对于在迁徙季节到来前就失去信号的个体,计算从监测开始到信号消失全过程的空间位点数据;(3) 对于整个监测期间都居留上海地区的个体,我们以最晚开始北迁的个体离开上海的时间为节点,将从监测开始到该时间点为止的数据作为这些居留个体的越冬期数据.
使用ArcGIS10.4[33]的Home Range Tool 2.0(HRT 2.0)插件,分别以最小凸多边形法(Minimum Convex Polygon, MCP)和固定核空间法(Fixed Kernel Estimation, FKE)计算每个个体在浦东东滩地区的活动面积[34].由于FKE最大的估计百分比为95%,为方便比较两种方法计算的活动面积,本研究分别提供了MCP和FKE的95%、75%、50%等3个百分比水平的活动面积.
此外,为了明确全部监测个体在浦东东滩越冬期间可能的活动范围并计算其栖息地利用方式,有必要计算基于全部监测个体的整体活动范围.固定核空间法(FKE)需要根据具体个体计算空间平滑度系数k,不适用于全部个体的整体活动范围计算[34],因此使用95%MCP法计算全部监测个体的越冬期整体活动范围.
1.4 研究区域的栖息地GIS解译
使用上述95%MCP法计算的全部监测个体的越冬期整体活动范围作为栖息地GIS解译区域.使用ArcGIS 10.4[33],利用Formosat-2卫星2 m级多光谱地面遥感图像(2012年6月),结合Google Earth图像和实地调查,对研究地区的景观结构进行目视解译.将研究区域的用地类型分为建筑、道路、裸地、农田、林地、草地、内陆水体、沟渠和滩涂等9种.其中,滩涂定义为沿海大堤以外的潮间带区域;沟渠定义为大堤之内的运河和水渠;农田为已经翻耕或已有作物的耕地;裸地为荒弃闲置的土地;内陆水体定义为内陆湖泊、河流、坑塘;道路仅指不透水路面区域;建筑定义为包括大堤、房屋以及非道路不透水地面在内的区域.部分监测个体在上海本土周边的一些岛屿和群岛例如九段沙和嵊泗列岛活动.限于研究能力,这些岛屿的栖息地情况尚无法实际调研,因此,本研究仅探讨活动区域在浦东东滩地区的个体的栖息地利用特征.
1.5 监测个体整体栖息地利用特征
首先,根据GIS解译的监测个体总活动区域内的各种用地类型的面积百分比,作为该种用地类型的资源可获得性.以越冬期全部有效GPS位点数乘以每种用地类型的百分比,作为监测鸭科鸟类对该种用地类型的理论利用率.其次,以越冬期全部有效GPS位点实际落在某种用地类型中的数量百分比作为鸭科鸟类对该种用地类型的观测利用率,并计算该观测利用率的Bonferroni 95%置信区间[35-36].如果观测利用率95%置信区间小于理论利用率,判断被监测鸭科鸟类对该种用地类型的利用率显著小于理论预期;如果观测利用率95%置信区间包含理论利用率,判断监测个体对该用地类型没有显著的倾向性;如果观测利用率95%置信区间大于理论利用率,判断监测个体对该用地类型的利用率显著大于理论预期.
1.6 监测个体对各主要栖息地类型的利用模式
当越冬期全部监测个体GPS位点在某一用地类型中的数量达到全部有效位点数的5%时,将该用地类型判断为主要栖息地类型.统计每小时中各监测个体实际落入各主要栖息地类型中的位点数.如果具体监测个体的主要栖息地类型数≥3,使用Kruskal-Wallis秩检验比较该个体在24 h内对不同主要栖息地类型强度的差异.如果某监测个体只有2种主要栖息地类型,使用Wilcoxon两个独立样本秩检验.
为分析监测个体在对各主要栖息地类型利用的时间分配,定义07∶00~18∶59为昼间,19∶00~06∶59为夜间.首先使用2个独立样本的Wilcoxon秩检验比较每个监测个体在整个监测期间昼、夜间对某个主要栖息地类型的利用强度是否存在显著差异.然后,使用2个相关样本的Wilcoxon秩检验分别就每个监测个体对不同主要栖息地类型昼、夜间利用强度进行两两比较.为控制自由度对检验结果的影响,依据每个个体具体的主要栖息地类型数,对显著值(P值)进行Bonferroni矫正[37].
1.7 监测个体活动节律
本研究使用的GPS卫星追踪器内置运动探测器,通过记录单位时间内(1 h)的振荡次数估计佩戴个体在该时段内的活动强度.因此,我们使用整个越冬期每个监测个体在各个小时内的振荡次数的平均值作为该个体在该计时单位中的活动强度,分析全部个体整体的以及每个个体在整个越冬期的24 h活动节律.所有统计分析均使用R3.5.0[38]完成.
2 结 果
2.1 斑嘴鸭和绿头鸭监测情况
研究时期为2017年1月18日到2018年1月1日,共监测斑嘴鸭5只、绿头鸭4只,总计9只个体.其中跟踪时间最长的个体是斑嘴鸭BZ005(315天、7 189位点),最短的是绿头鸭LT004(33天、363位点)(表2),平均监测时长(125.0±105.98) d、平均数据个数(2 777.3±2 509.60)个.9只监测个体中有4只监测到迁徙行为,包括2只斑嘴鸭(BZ002、BZ003)和2只绿头鸭(LT002、LT003)(表2).我们依据最早发生迁徙行为的个体BZ002的迁徙开始时间,将越冬季的结束时间设定为2017年4月3日(表2).
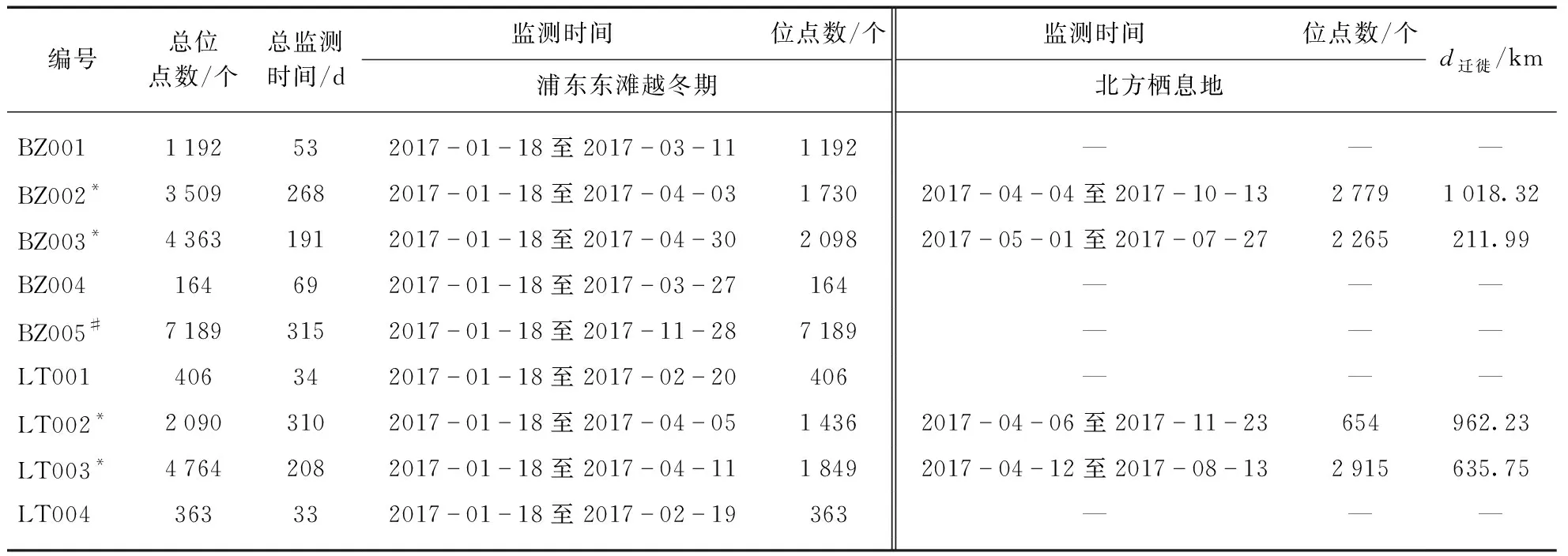
表2 监测个体GPS跟踪数据汇总
2.2 监测个体在浦东东滩的活动范围
6只在上海浦东东滩地区活动个体(4只斑嘴鸭和2只绿头鸭)的主要活动范围都集中在以大治河入海口为中心的广大滩涂和周边区域(图1).所有个体的平均活动面积为(22.206±33.264) km2(95% MCP)或(15.073±25.390) km2(95% FKE)(表3),且物种之间没有显著差异.
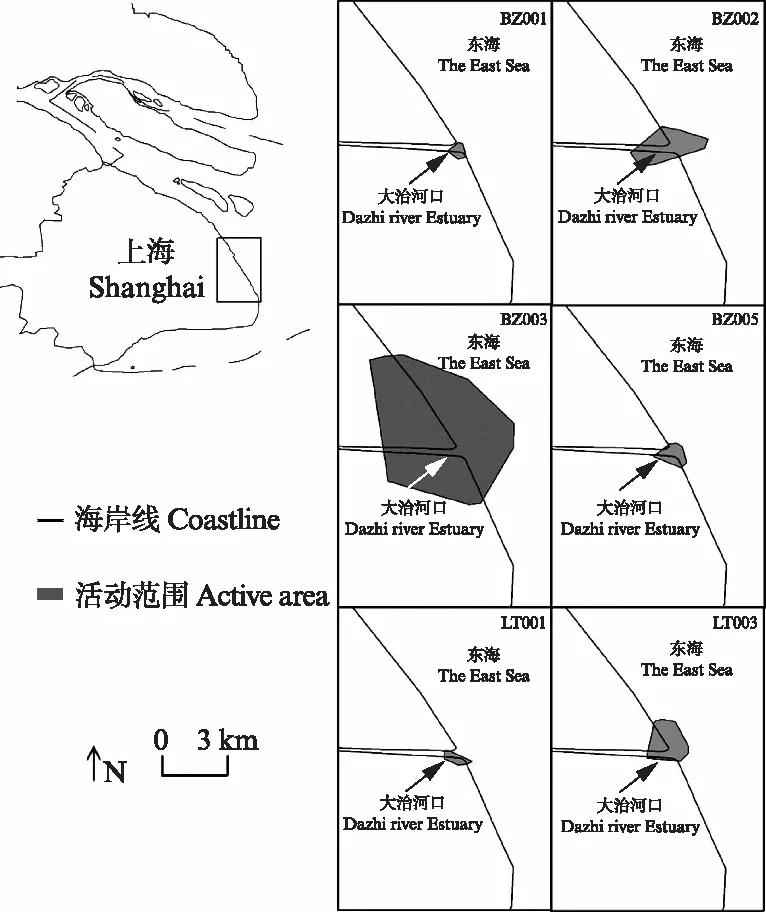
图1 监测个体越冬期在上海浦东东滩地区的活动范围Fig.1 The summary of the activity area of each tracked anatid individual during the wintering period in the Dongtan Wetland of Pudong, Shanghai
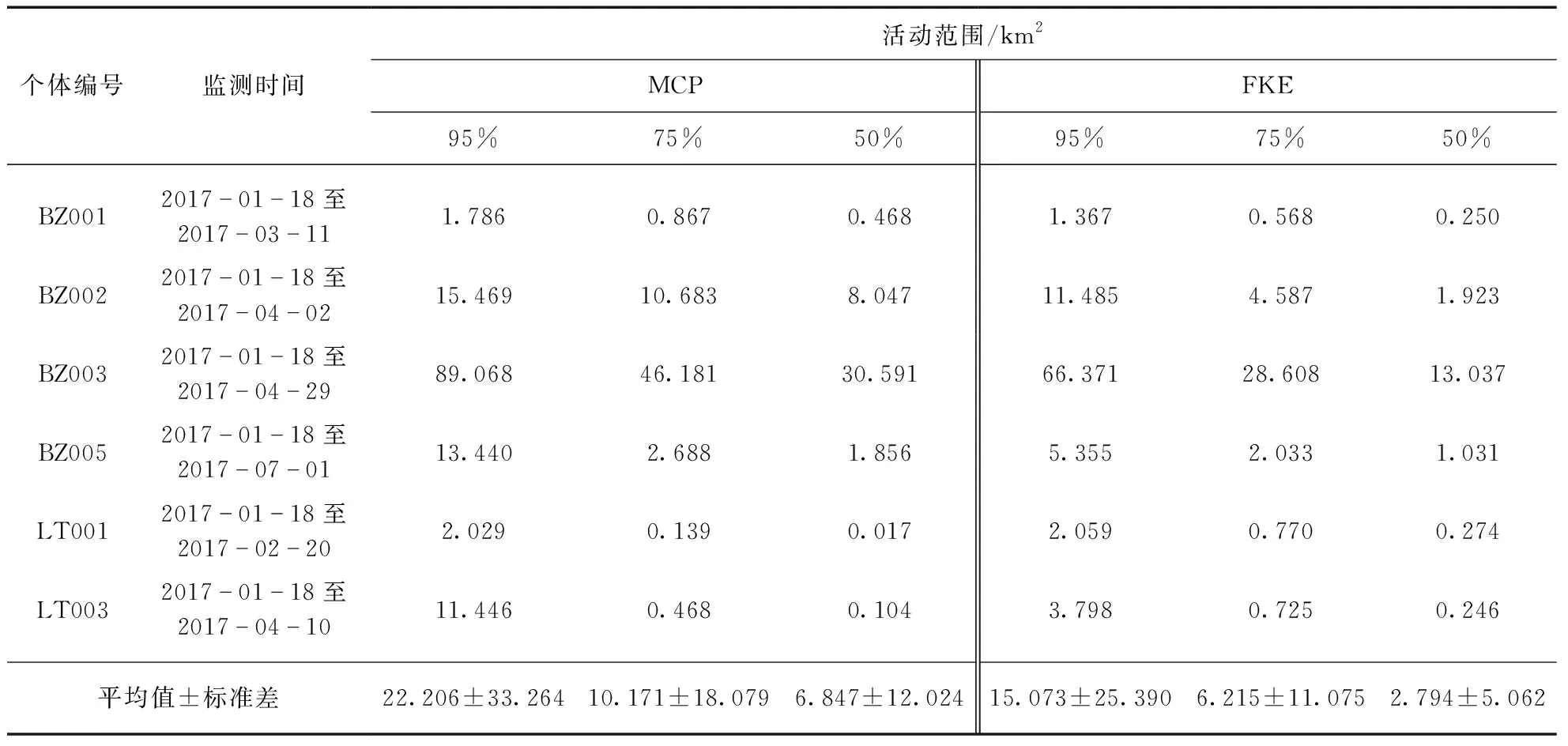
表3 监测个体的活动面积估计
2.3 监测个体整体栖息地利用特征
在所有监测个体的活动范围内(图2),利用率(观测利用率)超过总GPS位点数5%的主要栖息地类型依次为滩涂(38.45%)、沟渠(32.46%)、农田(19.88%)、内陆水体(8.85%)(表4).BonferroniZ检验显示,滩涂、内陆水体和沟渠的观测利用率显著大于理论利用率,说明鸭属鸟类对这3类用地类型的使用存在偏好(表4).监测个体对于建筑、道路、裸地、林地、草地和农田的观测利用率均显著低于该用地类型的理论利用率,均为利用较少的用地类型(表4).

图2 上海浦东东滩地区全部监测个体活动范围内的用地类型分布Fig.2 Landuse distribution in the activity area of all the tracked individuals in Dongtan Wetland of Pudong, Shanghai

表4 汇总监测个体GPS数据反映的各类栖息地利用强度
2.4 监测个体栖息地利用模式
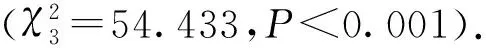
(1) 斑嘴鸭和绿头鸭在一天中主要有两个活动高峰: 第一个活动高峰均出现在7时左右,第二个活动高峰在18∶00左右(BZ001、LT003)或21时左右(BZ002、BZ003、BZ005).相比之下,BZ003和BZ005夜间活动强度均高于白天(图3).
(3) 不同追踪个体对不同类型栖息地昼夜利用程度并不一致.所有追踪个体均利用了滩涂和水渠: 仅BZ002和LT003对滩涂的昼夜利用程度上存在显著性差异;BZ003、BZ005、LT001、LT003 4个个体对水渠的昼夜利用程度存在显著性差异.监测期间,在4个利用农田的追踪个体中,BZ003和LT003对农田昼夜利用程度存在显著性差异;只有BZ003利用了内陆水体,且对内陆水体的昼夜利用存在显著差异.BZ001对滩涂和沟渠的昼夜利用程度均不存在显著性差异(表5).
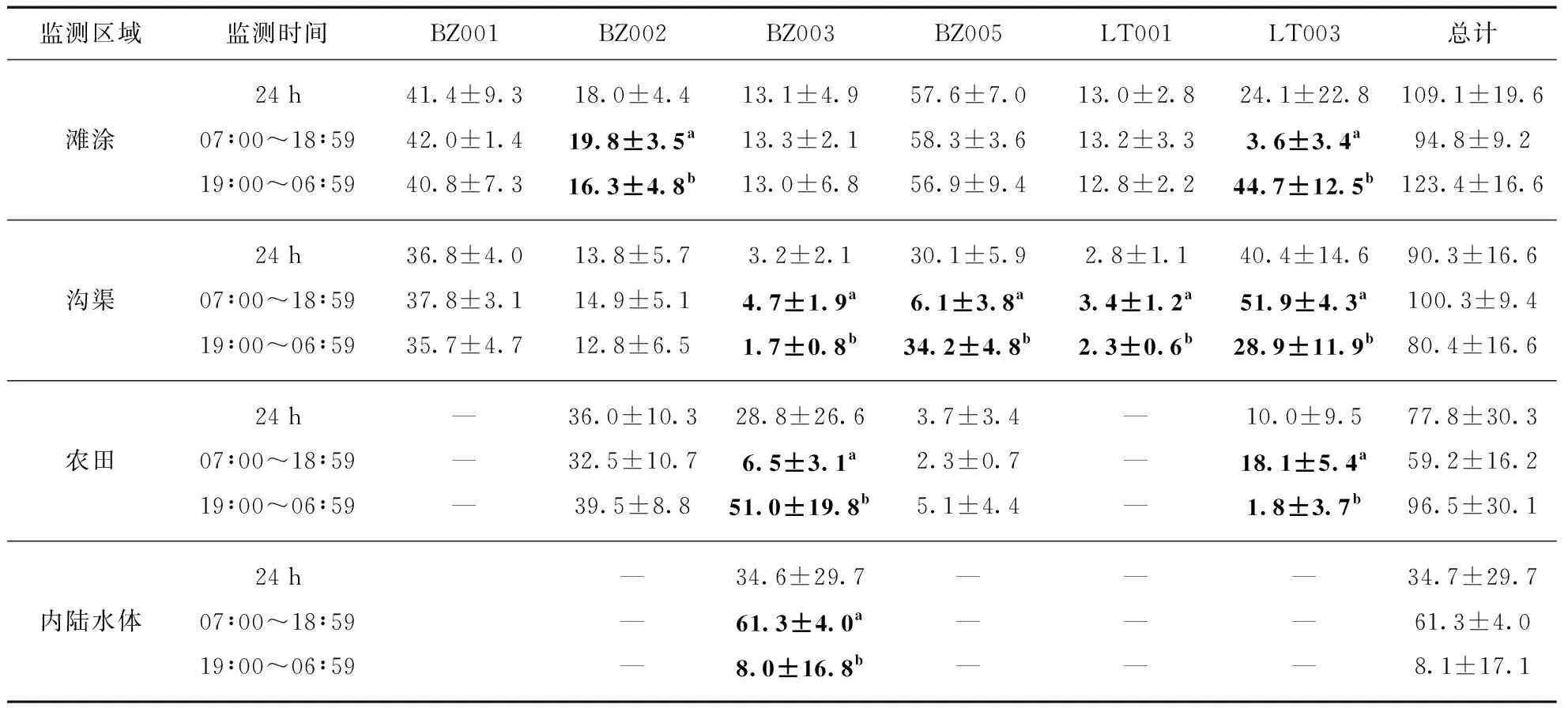
表5 监测个体全天各种栖息地中GPS位点的分布
(4) 同一时期(白天或夜晚)斑嘴鸭和绿头鸭对不同类型栖息地利用强度多存在显著性差异.无论白天还是夜晚,斑嘴鸭和绿头鸭对农田和滩涂利用强度均存在显著性差异;在夜晚,斑嘴鸭和绿头鸭对农田和沟渠的利用强度存在显著性差异(表6).

表6 监测个体各种栖息地类型昼夜间利用强度的比较
3 讨 论
3.1 迁徙的时间与路径
本研究结果表明,4月上旬为浦东东滩斑嘴鸭和绿头鸭普遍的迁飞时间,迁徙成功率高,除1个个体中途折返外,其余个体均到达繁殖地(表2).斑嘴鸭和绿头鸭在浦东东滩的停留时间较长,且部分个体(BZ005)在接下来的繁殖季并不发生迁徙行为(表3).这一现象在研究洪泽湖地区雁鸭类的迁徙行为时也有报道,部分斑嘴鸭和绿头鸭个体甚至会在洪泽湖地区繁衍[12].食物资源以及潜在配偶(家禽)资源可能是改变其迁徙节律的主要原因.
3.2 空间活动范围
本研究结果表明,浦东东滩的斑嘴鸭和绿头鸭都有自己相对固定的活动范围,未观察到监测个体去附近其他适合鸟类越冬的地方(例如崇明东滩和九段沙)活动.监测个体活动范围大小变异较大,且受制于样本量,没有发现越冬期活动范围存在种间差异(表3).但是,所有个体的活动范围都在大治河河口附近,且不同个体之间活动区域有重合(图1),结合鸟群现场观察,支持不同鸭科鸟类物种之间存在明显的种间吸引和互利现象[39]的观点.
有研究指出,雁鸭类的活动范围与物种、年龄和性别没有关系,只与个体使用的觅食地点和栖息地类型的多样性有关[15],本研究中,只利用两种栖息地类型的斑嘴鸭BZ001与绿头鸭LT001的活动范围面积最小,利用三种栖息地类型的斑嘴鸭BZ002、BZ005与绿头鸭LT003的活动范围面积其次,利用了4种栖息地类型的斑嘴鸭BZ003的活动范围面积最大,活动范围的大小之间并不存在种间差异(图1,表3).这是否意味着控制鸭科鸟类的栖息地与觅食地数量,可以减少它们的活动范围面积,降低它们携带的禽流感病毒的影响范围,有待后续研究.
3.3 栖息地利用特征
虽然一些人造栖息地确实可以提供水鸟适宜地栖息,但不能完全替代自然栖息地的生态服务功能[40].本研究发现的4种鸭科鸟类主要栖息地类型中,除了滩涂为自然栖息地外,其余3种均为人造栖息地,但是滩涂的利用率是所有4种栖息地中最高的(表5),这体现了自然栖息地对野生水鸟的重要性.另一方面,虽然6只主要监测个体的栖息地利用方式不尽相同(图3),但是滩涂和沟渠却是它们都利用的栖息地类型(表6),其中沟渠作为人造栖息地成为利用强度仅次于自然栖息地的生境类型(表5).华宁等[41]指出人工湿地环境影响水鸟栖息的主要环境因子是水面积和人为干扰强度.研究区域中的沟渠主要由围垦区的运河和灌溉水渠组成,宽阔的河道上极少有人类活动,因此为鸭科鸟类的栖息提供了条件.
另一方面,已有研究结果表明,越冬期鸭科鸟类很少利用稻田[42-43].本研究中,虽然总体数据上显示农田生境的利用率低于预期(表4),且不同的个体在农田上的使用差异明显(图3),但是一些个体(BZ002和BZ003)在夜间存在大量利用农田生境的现象(图3).现场观察显示,冬季收割后的农田上有大量遗弃稻谷,这为植食性的鸭科鸟类提供了食物资源.鸭科鸟类光顾的农田都在远离村庄的地方,并且利用时间多为夜间(图3),这与人类田间作息时间是相反的.鸭科鸟类避开了人类作业的高峰期,说明人类活动仍然是主要的干扰源.其次,田间有家犬和家猫的活动痕迹,同时观察到一只斑嘴鸭尸体残骸有明显的食肉哺乳动物啃食痕迹.以上现象说明,农田虽然有鸭科鸟类需要的食物资源,但是较强的人类活动干扰和被捕食风险,限制了它们的利用率.因此,虽然浦东东滩农田面积很大,但实际利用率却不高.农田是浦东东滩地区人类活动、家养动物活动较密集的区域,深入研究野生鸭科鸟类对农田的利用问题对于分析人类和野生动物的交互作用,进而分析这种交互作用对野生动物疫源疫病的传人风险等都有重要意义.
本次研究的监测个体较少,出现了单个个体斑嘴鸭BZ003对内陆水体的利用量较大而干扰整个数据集的统计结果的现象(图3).因此,进一步研究鸭科鸟类对内陆水体的利用问题则需要监测更多个体,对每个个体进行单独分析,才能得到更准确的结果.
3.4 活动节律
本次研究结果表明,斑嘴鸭和绿头鸭对栖息地的利用存在时间节律(图3).已有研究[13]表明,越冬季野生鸭属鸟类白天行为主要以休息为主,而夜间有取食行为.结合本次研究的结果,推测不同的栖息地可能对鸭科鸟类的作用不同,有些为栖息地点,有些为觅食地点.
本次研究发现,斑嘴鸭和绿头鸭在清晨与黄昏存在活动高峰,而在夜间均保持较高的活动强度(图3).研究表明,水禽需要依靠摄食抵御夜间低温恶劣的环境[44],所以在黄昏与清晨存在高强度的觅食现象[13].因此,本次研究中鸭属鸟类在清晨与黄昏出现的活动高峰可能是觅食行为.限于技术手段,我们尚无法确定监测个体在各栖息地的具体行为,而传统的观察法如瞬时扫描和目标动物观察法可以直接了解到研究个体的行为规律[45],故今后的研究方向应将GPS追踪法与传统的观察法结合起来.
致谢:感谢上海市浦东新区林业站的工作人员为本项目的开展提供实验动物和后勤保障.
