海拔高度对暗紫贝母叶特征的影响
2021-10-16陈文年蔡平原
陈文年 蔡平原


摘 要:為探索海拔高度对暗紫贝母(Fritillaria unibracteata)叶特征的影响,该研究在岷江上游的卡卡山北坡设置低、中、高三个海拔部位,并在高、低部位之间进行植株的移栽试验,并测定每个部位植株的叶寿命、气孔密度和长度、单叶面积及比叶面积等叶特征。结果表明:(1)叶寿命从低海拔部位的121.3 d,到高海拔部位的108.5 d,缩减了大约13 d。(2)从低海拔到高海拔,气孔密度逐渐升高,低海拔部位只有48.5 number·mm-2,高海拔部位与之相比增加了42.8%。(3)而气孔长度则随海拔升高而降低,从低海拔部位的82.4 μm 到高海拔的71.3 μm,降低了13.5%。(4)从低海拔到高海拔,单叶面积和比叶面积都逐渐增加,其中单叶面积由低海拔部位的1.61×10-4 m2到高海拔部位的2.20×10-4 m2,增加了36.6%;比叶面积则由低海拔部位的7.7×10-4 m2·kg-1到高海拔部位的12.5×10-4 m2·kg-1,增加了62.3%。(5)另外,同一海拔部位的对照植株和移栽植株之间叶特征并无显著差异;植株在移栽后总是表现出移栽后所在部位对照植株的特点,而没有表现出移栽前所在部位对照植株的特点。综上结果表明,海拔高度对暗紫贝母叶特征影响显著,暗紫贝母的叶特征在海拔梯度上具有较大的可塑性。
关键词:暗紫贝母,叶寿命,气孔密度,气孔长度,比叶面积,高山生态
中图分类号:Q948.11
文献标识码:A
文章编号:1000-3142(2021)09-1450-07
Abstract:As an important traditional Chinese medicinal plant,Fritillaria unibracteata is mainly distributed in alpine areas with an elevation above 3 000 m. Unsustainable harvesting of this species has greatly destroyed its natural habitats and made it more valuable. Among all factors affecting its growth,altitude is the most obvious one. To explore the effects of altitude on leaf traits of F. unibracteata,three altitudinal sites (low site,medium site and high site)were set up on the north-facing slope of Kaka Mountain in the upper reaches of Min River,and plant transplant experiments were carried out between low site and high site,which was 108.5 d. Then leaf traits including leaf lifespan,stomatal density and length,single leaf area and specific leaf area in each site were measured. In these experiments,some meaningful results were produced. The results were as follows:(1)From low site to high site,leaf lifespan shortened,and there were significant differences among sites. Leaf lifespan at low site was 121.3 d,about 13 d was shortened more than that at high site,which was 108.5 d. Along with altitude rise,stomatal density increased. (2)Compared with low site with only 48.5 stomas per square milimeter,stomatal density in high site increased by 42.8%. (3)Contrary to stomatal density,stomatal length decreased along with the increase of altitude. Compared with low site with 82.4 μm,stomatal length at high site with 71.3 μm reduced by 13.5%. (4)As to single leaf area and specific leaf area,they both showed a rising trend from low site to high site. At low site with 1.61×10-4 m2,single leaf area at high site reached 2.20×10-4 m2,36.6% more than that at low site; similarly,specific leaf area at high site was 12.5×10-4 m2·kg-1,62.3% higher than that at low site with 7.7×10-4 m2·kg-1. At the same altitudinal site,the control plants and transplants did not differ significantly. (5)For those transplanted plants,they always performed in accordance with control plants at the same altitudinal site after transplant experiment,but did not show characters of control plants at different altitudinal sites before transplant experiment. It can be concluded that altitude exerts an evident influence on leaf traits of F. unibracteata,and that leaf traits of F. unibracteata is of great plasticity when environment conditions change.
Key words:Fritillaria unibracteata,leaf lifespan,stomatal density,stomatal length,specific leaf area,alpine ecology
百合科贝母属的暗紫贝母(Fritillaria unibracteata)是一种珍稀的药用植物,主要生长于青藏高原东部3 200~4 500 m 的高山环境中,分布区相对狭小(四川植物志编辑委员会,1991;徐国钧,1997;青海植物志编辑委员会,1999)。因为其生长条件苛刻,人工栽培困难,种植面积较小,所以,暗紫贝母的药材供应主要来自于野生资源。随着人工采挖的不断进行,其野生生境逐渐被破坏,野生资源库存量逐渐减少,药材供应趋于枯竭。因此,寻找其最佳生长环境,并进行规模化的人工栽培是解决问题的关键。
影响暗紫贝母生长的环境因子有很多,如温度、光照、水份、土壤、坡面及海拔等。其中,海拔与其他生态因子的关系密切,海拔改变会导致其他因子发生相应的级联变化,从而可能会进一步影响暗紫贝母的生长。因此,海拔是众多因子中引人注目的一种。同时,叶通过光合作用影响营养物质的积累,与暗紫贝母药用部分的产量密切相关,在暗紫贝母的生活史中具有十分重要的作用,由此可见,叶特征是暗紫贝母生长的重要度量指标,也是易于测量的性状指标。那么受海拔的影响,暗紫贝母的叶特征会随海拔梯度如何变化呢?
已有的研究主要集中于药理、药化方面(朱丹妮等,1992;高山林等,2000;韵海霞和陈志,2009)、组培方面(蔡朝晖等,1991;李隆云等,1995)、群落及个体生态方面(陈士林等,1989,1997;徐波等,2013;陈文年等,2012,2015,2016),而用海拔作生态因子对暗紫贝母叶特征进行的研究还相对薄弱(陈文年等,2012,2015)。因此,本研究选择岷江源区松潘县的卡卡山作为研究地点,探讨海拔高度对其叶形态特征的影响,以探索暗紫贝母叶最佳的生长海拔高度,从而寻找暗紫贝母的最佳生长因子组合,为人工种植中产量最大化提供理论依据。
1 材料与方法
1.1 研究地概况
研究地位于岷江源头地区的松潘县卡卡山(103°40′ E、32°59′ N),该地区年均温为2.8 ℃,1月均温-7.6 ℃,七月均温9.7 ℃,年降雨量为634.8 mm,其中72%发生在6—8月的夏季,土壤类型为黑毡土,卡卡山山脚、山顶垂直高度差580 m左右(Chen et al.,2011;陳文年等,2012)。
1.2 样方设置及试验设计
研究地点设在卡卡山北坡,取低、中、高三个海拔部位(海拔分别为3 500、3 750、3 950 m)。高、低两个海拔部位沿水平方向设置A、B两个30 m × 15 m(长×宽)的调查样方(两样方间水平距离15 m左右),中间海拔部位只设置一个同样大小的A样方,在每个样方中分别随机选取30株二年生(所谓“一匹叶”阶段,2~3 a后开花结果)的暗紫贝母个体作为研究对象,以A样方中植株作为对照,在不同海拔间作对比分析。所有样方中被选择的植株都作了标记,并于2010年7月开始在各个样方周围设置铁丝围栏以防人畜干扰。
为确定海拔对暗紫贝母的叶特征是否有确切的影响,在高海拔部位(3 950 m)和低海拔部位(3 500 m)的B样方之间进行植株的对换移栽试验,即把高海拔B样方中被选择的30株植株移栽到低海拔部位的B样方中,同时把低海拔部位B样方中被选择的30 株植株移栽到高海拔部位的B样方中。移栽过程中采用连土移栽,即每株暗紫贝母连同其植株下面的土块一起移栽,土块的长、宽、深大小尺寸为0.20 m × 0.20 m × 0.25 m。整个移栽试验于2010年9月上旬进行,所有的移栽植株都作了标记,以A样方中的植株作为对照。移栽实验完成之后,在高、低海拔部位就产生出4种植株类型,即高海拔部位A样方的对照植株(HA)、高海拔部位B样方中的来自低海拔的移栽植株(HB);低海拔部位A样方的对照植株(LA)、低海拔部位B样方中的来自高海拔的移栽植株(LB)。另外,中间海拔部位还有1个样方,即中间部位A样方中的植株(MA)。
1.3 测定指标
1.3.1 气孔长度和气孔密度 于2013年7月中旬,在每一样方中的标记植株上,选择最大的2 张叶片,用小剪刀轻轻剪下,放入预先准备好的信封中,做好标记。然后用印迹法测定气孔数目(Beaulieu et al.,2008;Miller-Rushing et al.,2009)。在10×目镜、20×的物镜下,每张叶片选择5 个不同视野计数气孔数目,然后计算视野面积(张淼丽,2009),并进一步换算出气孔密度。
同时在10×目镜、40×物镜下,用目镜测微尺测量气孔的长度(这里指气孔器保卫细胞长轴的长度),每一叶片取5 个视野,每个视野中选3~5 个气孔测量,各视野中气孔的平均值作为该叶片气孔长度的代表值。
1.3.2 叶面积和比叶面积 在测量完气孔的长度和密度后,把“1.3.1”中所剩下的两片叶片用于测量叶面积和比叶面积。叶面积用叶面积仪(美国 CID 公司,型号 CI-203)测量,在测量完叶面积之后,将叶片放入烘箱中烘48 h,温度设定为恒温65 ℃,取出后用万分之一电子天平(型号:FA1004)测量其重量,并计算其比叶面积(specific leaf area)(单位叶干重所具有的叶面积)。比叶面积(SLA)的计算公式为:SLA= 叶片面积 (m2 )/叶片干物质量 (kg),做好数据记录,以备后续研究之用。
1.3.3 叶的生存期限(叶寿命)各样地中标记的暗紫贝母植株,从其每一张叶片长度达到1 cm算起,到该叶片出现枯黄为止,之间的时间作为这一叶片的叶寿命,叶寿命的计算在2013年完成。每一植株上用于计算气孔密度、叶面积和比叶面积的2 张叶不再计算叶寿命。
1.4 统计方法
用 SPSS 17. 0 以及Excel办公软件进行数据分析和制图,所得数据以“平均值±标准误”表述。
2 结果与分析
2.1 叶的寿命
从表1可以看出,三个海拔部位之间差异显著。在三个海拔部位自然生长的植株中,高海拔部位植株(HA)的叶寿命最短,低海拔部位植株(LA)叶寿命最长,中间部位(MA)介于二者之间。其中,低海拔部位(LA)平均叶寿命达到121.3 d,比高海拔部位(HA)的108.5 d多大约13 d,多11.8%。
从不同样方间t-检验结果(表2)可以看出,同一海拔部位的对照植株(A)和移栽植株(B)之间,叶寿命无显著差异。但不同海拔部位的对照植株和移栽植株之间却差异显著。低海拔部位的对照植株(LA)大于高海拔部位的移栽植株(HB),而高海拔部位的对照植株(HA)小于低海拔部位的移栽植株(LB),即植株被移栽到相应部位后,其叶寿命表现出移栽后所在部位对照植株的特点,而没有表现出移栽前所在部位对照植株的特点。
2.2 气孔密度和气孔长度
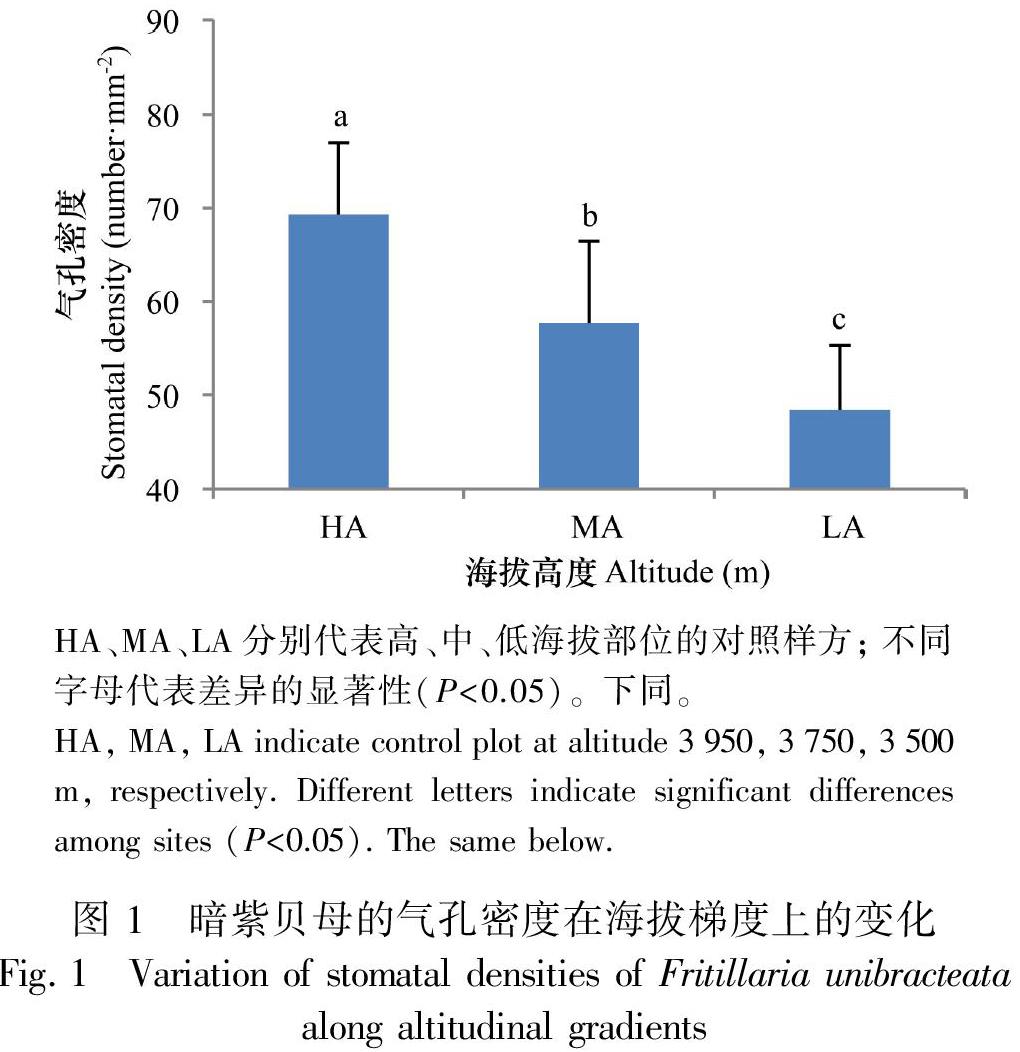
从图1可以看出,从低海拔到高海拔,暗紫贝母的气孔密度逐渐增加,各部位间差异显著。低海拔部位的对照植株(LA),气孔密度只有48.5 number·mm-2,中海拔部位(MA)与之相比气孔密度增加了18.9% ;同样,高海拔部位的对照植株 (HA)和中、低海拔部位的对照相比,其气孔密度分别增加了20.1%和42.8%(图1)。
随着海拔的增加,气孔的长度逐渐变小,这一点与气孔密度的变化趋势不同。低海拔部位的对照植株(LA),气孔长度达到82.4 μm;中海拔部位(MA)与之相比,减少了6.3%;高海拔部位对照植株 (HA)与中海拔部位(MA)相比,又减少了7.6%。另外,除了高、低海拔部位对照植株之间的差异达到显著以外,其余部位对照植株之间的差异都未达到显著,这一点与气孔密度有所区别(图2)。
从表3可以看出,在同一海拔部位(高海拔或低海拔),自然生长的植株(A)与移栽的植株(B)其气孔密度无显著差异,气孔长度也无显著差异。和叶寿命类似,移栽后的植株在气孔密度和气孔长度方面也都表现出移栽后所在部位对照植株的特点,而没有表现出移栽前所在部位对照植株的特点(表3)。
2.3 单叶面积和比叶面积
从低海拔部位到高海拔部位,单叶面积逐渐增加,高、低海拔部位之间差异显著。高海拔部位的对照植株(HA),单叶面积达2.20×10-4 m2,比低海拔部位对照植株(LA)的1.61×10-4 m2大0.59×10-4 m2,大36.6%(表4)。比叶面积的变化趋势和单叶面积类似,高海拔部位的对照植株(HA)比叶面积最大,低海拔部位的对照植株(LA)比叶面积最小,中间部位(MA)于二者之间。低海拔部位的对照植株(LA),其比叶面积只有7.7×10-4 m2·kg-1,相比高海拔部位对照植株(HA)的12.5×10-4 m2·kg-1 ,少38.4%。与单叶面积情况不同的是,比叶面积在三个部位对照植株之间的差异都达到显著(表4)。
在同一海拔部位的对照样方(A)和移栽样方(B)之间,其植株单叶面积的差异不显著,比叶面积差异也不显著,不论是在高海拔部位还是在低海拔部位都是如此(表5)。另外,植株被移栽后,其单叶面积和比叶面积也是表现出移栽后所在部位对照植株的特点,而没有表现出移栽前所在部位对照植株的特点,这与叶寿命、气孔密度和气孔长度等叶特征的表现类似(表5)。
3 讨论
3.1 海拔对叶寿命的影响
作为植物的营养器官,叶是大多数植物进行光合作用的主要场所,叶寿命的长短是植物受其所处环境中各种生态因子综合作用的结果(Krner,1991),也是植物适应环境的重要方式之一(Cordell et al.,2001)。长的叶寿命能使植物获得更多的光合作用产物,对其生长和繁殖过程的完成具有较大影响(Wright et a1.,2004)。在针叶林中的研究表明,随着海拔升高,气温降低,环境条件逐渐恶劣,植物的叶寿命会增加(Smith & Hinckley,1995;Reich et al.,1996)。在本研究中,暗紫贝母叶片所表现出来的情况与上述研究结果并不相符,其原因是暗紫贝母属于低矮的草本植物,生长于地面,其生活期(包含叶寿命)受到地面积雪及融雪时间的强烈影响。在卡卡山地区,由于整体海拔较高,每年积雪覆盖的时间较长,特别是高海拔部位,积雪相对较早,融雪相对较迟,而低海拔部位却积雪相对较迟,融雪相对较早,在高、低海拔部位之间融雪时间相差甚至达到20多天(陈文年等,2015;Chen et al.,2008,2011)。这种积雪的覆盖必然会影响到叶片的生长时间,从而影响到叶片寿命的长短。而针叶林树种,一般为高大乔木,覆盖在地面的积雪对其针叶寿命没有影响或影响很小。
在较高海拔部位,由于雨、雾、冰雹等降水相对较多,再加上晚融的积雪也会补充土壤的含水量,因此,土壤含水量相对较多(陈文年等,2015)。有研究表明,在水分有效性越差的生境下,其物种的叶寿命越长;而水分相对充足的环境中,叶寿命反而越短(Smith & Hinckley,1995;Luo et al.,2005)。顯然,本研究的结果与上述结果一致,说明土壤含水量对叶片寿命的影响具一定的普遍性。本研究结果表明,虽然随着温度的降低,叶寿命也可能会增加(Smith & Hinckley,1995;Luo et al.,2005),但是在积雪覆盖和土壤水分这两种因子的强烈影响下,高海拔部位温度的影响就相对被弱化,因此,多种因子综合作用的结果使叶寿命相对变短。
3.2 海拔对气孔密度和长度的影响
在高海拔部位,由于积雪覆盖的影响,叶寿命相对缩短,相应光合时间也就自然缩短,再加上在高海拔部位雨、雪、冰雹等恶劣天气更加频繁,植物气孔开放时间缩短,与外界能够进行有效气体交换的时间必然减少,所以,这对植物光合产物的积累以及生活史的完成会造成一定的不利影响。另外,随海拔的升高,O2 和CO2浓度也在逐渐减少,这对高海拔部位植物叶片与外界的气体交换也是一个不利因素。不同的海拔部位,植物长期处于这样的环境中,自然会形成与之相适应的不同形态结构特征。在本研究中,随海拔升高,叶片气孔数目增加,同时气孔变小。根据小孔扩散规律,气体扩散的速率不与小孔的面积成比例,而是与小孔的周长成比例。因此,暗紫贝母气孔密度和大小在海拔梯度上的表现在较大程度上增加叶片的气体交换能力,可以看成是植物在长期适应中发展出的对气体交换的一种补偿。本文的这一结果与其他学者的研究结果一致(Krner,1986)。
3.3 海拔对叶面积和比叶面积的影响
植物叶面积的大小很容易受到环境中各种因子的影响。一般而言,生活在干旱环境中的植物,其叶片往往小而厚,栅栏组织发达,有较厚的细胞壁和角质膜(李正理和张新英,1983;Takahashi & Miyajima,2008);相反,生活在荫蔽潮湿环境中的植物则叶片薄而大,角质膜不发达(陆时万等,1991)。
在本研究中,暗紫贝母的叶面积和比叶面积随海拔升高而逐渐升高,其原因有几个方面。其一,在岷江源头地区,存在着明显的季节性积雪现象,高、低海拔之间的融雪时间相差较大,如在本研究地可达25 d(陈文年等,2012,2015),在低海拔部位由于积雪的早融,白天气温会明显地升高,蒸藤作用增强,土壤水分挥发较快,植物会处于相对更为干燥的环境中,植物为适应这样的环境自然会形成相对小而厚的叶型,因而比叶面积较小。而高海拔部位的植株,由于积雪的晚融,植物体的温度不会明显地受白天气温升高的影响,植物体蒸藤作用弱或无,而且土壤因为融雪水的持续灌溉保持较为潮湿的状态,这样的生境自然会使植物的叶片朝向相对大而薄的方向发展,因而比叶面积较大。其二,积雪早融的低海拔部位,由于过早失去积雪的保护作用,夜间温度会降的很低,甚至可以达-6 ℃,而白天温度又会升的很高,可以达到17 ℃(Chen et al.,2008,2011),这种反复的冻融交替对植物来说无疑是一种很苛刻的环境,植物为了自保和减轻冻害,其叶片只能朝向相对小而厚的方向发展,于是其比叶面积相对变小。其三,在高海拔部位,云雾相对更容易聚集,各种降水更容易形成,再加上积雪的晚融,其土壤中的水分含量比起低海拔处反而更充足,可达到55.1%,而低海拔处却只有38.4%(陈文年等,2012,2015)。这样一来,在高海拔部位靠近地表处的微生境中也会更潮湿,这也更利于形成相对大而薄的叶片,于是其比叶面积变大。
Marcelo et a1.(2009)对某些木本植物作了研究,认为随着海拔的升高,叶面积和比叶面积降低,并认为这是由于海拔升高后,辐射增强、风力加大、环境干燥而造成的。显然,本研究的结果与之不一致。其原因可能是这些木本植物相对比较高大,积雪覆盖对其叶不会造成明显的影响,因此气温、风和辐射可以成为主要的影响因子。而本研究中情况不一样,暗紫贝母属于是比较低矮的草本,特别是1年生、2年生个体几乎是贴着地面生长,即使是3年生、4年生的个体也只有20 cm高或稍高一点,其周围的许多灌丛和草本都会对其造成一种荫蔽的小生境,在这种情况下,尽管随着海拔高度的增加,紫外辐射和风的作用会有所增强,但由于这些荫蔽小生境的存在,就削弱或抵消了紫外辐射和风的影响,即积雪的覆盖可屏蔽或削弱其他因子的作用,甚至积雪的存在,可使其他生态因子出现倒置的情况,因此,这里的结果不同。
3.4 高低海拔部位之间移栽植物的验证作用
无论是从叶片寿命、气孔密度和大小来看还是从叶面积及比叶面积来看,移栽植株都倾向于移栽后部位中植株的特征,而与之前所在的部位无关。这说明暗紫贝母这些叶特征很大程度上是由积雪融化时间、土壤水分及小生境局部温度等的影响造成的,而与不同海拔部位的紫外辐射及移栽前土壤的性质没有太大的关系。同时从移栽实验还可以看出,暗紫贝母的这些叶特征,由于受环境因子作用,可塑性比较大,这种叶特征在海拔梯度上的变化应该不遗传。
4 结论
随着海拔的升高,暗紫贝母的叶寿命缩短、气孔密度增加同时气孔长度减小、单叶面积和比叶面积增加。暗紫贝母叶特征在海拔梯度上的变化显著,其叶特征在海拔梯度上具有较大的可塑性。
参考文献:
BEAULIEU JM,LEITCH IJ,PATEL S,et al.,2008. Genome size is a strong predictor of cell size and stomatal density in angiosperms[J]. New Phytol,179(4):975-986.
CAI CH,GAO SL,XUE DR,et al.,1991. Effect of different cultural conditions and methods on the growth of cultured bulb of Fritillaria unibracteata Hsiao et K. C. Hsia[J]. J Chin Pharm Univ,23(6):367-369.[蔡朝晖,高山林,徐德然,等,1991. 不同培养条件及方法对组培暗紫贝母生长的影响[J]. 中国药科大学学报,23(6):367-369.]
CHEN WN,CHEN FJ,XIE YH,et al.,2012. Variation of phenology and bulbs of Fritillaria unibracteata along altitudinal gradients[J]. Acta Pratac Sin,21(5):319-324.[陈文年,陈发军,谢玉华,等,2012. 暗紫贝母的物候和鳞茎在海拔梯度上的变化[J]. 草业学报,21(5):319-324.]
CHEN WN,XIAO XJ,CHEN FJ,et al.,2015. Reproductive organ variation in Fritillaria unibracteata along an altitudinal gradient[J]. Acta Pratac Sin,24(12):155-163.[陈文年,肖小君,陈发军,等,2015. 暗紫贝母的繁殖器官在海拔梯度上的变异[J]. 草业学报,24(12):155-163.]
CHEN WN,WANG H,XIAO XJ,et al.,2016. Effects of slope aspect on growth and reproduction of Fritillaria unibracteata(Liliaceae)[J]. Acta Ecol,36(24):8174-8182.[陳文年,王辉,肖小君,等,2016. 坡向对暗紫贝母生长和繁殖特征的影响[J]. 生态学报,36(24):8174-8182.]
CHEN WN,WU Y,WU N,et al.,2008. Effect of snow-cover duration on plant species diversity of alpine meadows on the eastern Qinghai-Tibetan Plateau[J]. J Mt Sci,5(4):327-339.
CHEN WN,WU Y,WU N,et al.,2011.Effect of snowmelt time on growth and reproduction of Pedicularis davidii var. pentodon in the eastern Tibetan Plateau[J].Plant Biosyst,145(4):802-808.
CHEN SL,XIAO XH,CHEN XY,1989. Study on community ecology of Fritillaria unibracteata[J]. J Chin Med Mat,12(11):5-8.[陈士林,肖小河,陈善墉,1989. 暗紫贝母的群落生态研究[J]. 中药材,12(11):5-8.]
CHEN SL,XIAO XH,CHEN XY,1997. Unmerical studies on spatial distribution pattern of Fritillaria unibracteata community[J]. J SW Chin Norm Univ (Nat Sci Ed),22(4):416-420.[陈士林,肖小河,陈善墉,1997.暗紫贝母植被分布格局的数值分析[J]. 西南师范大学学报(自然科学版),22(4):416-420.]
CORDELL S,GOLDSTEIN G,MEINZER FC,et al.,2001. Regulation of leaf life-span and nutrient-use efficiency of Metrosidero polymorpha trees at two extremes of a long chronosequence in Hawaii[J]. Oecologia,127(2):198-206.
Editorial Board of Sichuan Flora,1991. Sichuan Flora (Vol. 7 )[M]. Chengdu:Sichuan National Press:57-59.[四川植物志編辑委员会,1991. 四川植物志(第七卷)[M]. 成都:四川民族出版社:57-59.]
Editorial Board of Qinghai Flora,1999. Qinghai Flora ( Vol. 4)[M]. Xining:Qinghai Pelples Publishing House:277-278.[青海植物志编辑委员会,1999. 青海植物志(第四卷)[M]. 西宁:青海人民出版社:277-278.]
GAO SL,XIA Y,TAN FP,2000. The pharmacological action of cultured bulb of Fritillaria unibracteata Hsiao et K. C. Hsia[J]. J Plant Resour Environ,9(1):4-8.[高山林,夏艳,谭丰苹,2000. 组织培养暗紫贝母的药理作用[J]. 植物资源与环境学报,9(1):4-8.]
KRNER C,BANNISTER P,MARK AF,1986. Altitudinal variation in stomatal conductance,nitrogen content and leaf anatomy in different plant life forms in New Zealand[J]. Oceologia (Berlin/West),69:177-188.
KRNER C,1991. Some often overlooked plant characteristics as determinants of plant growth:A reconsideration[J]. Funct Ecol,5(2):162-173.
LI LY,ZHOU YS,DAI M,et al.,1995. Research on chemical components and primary pharmaceutical actions of cultured bulb of China Fritillaria unibracteata[J]. Chin J Chin Mat Med,20(2):78-80.[李隆云,周裕书,代敏,等,1995. 暗紫贝母鳞茎再生组织培养技术研究[J]. 中国中药杂志,20(2): 78-80.]
LI ZL,ZHANG XY,1983. Plant anatomy[M]. Beijing:Higher Education Press:246-280.[李正理,张新英,1983. 植物解剖学[M]. 北京:高等教育出版社:246-280.]
LU SW,XU XS,SHEN MJ,1991. Botany (Vol. 1)[M]. 2nd ed. Beijing:Higher Education Press:142-168.[陆时万,徐祥生,沈敏健,1991. 植物学(上册)[M]. 2版. 北京:高等教育出版社:142-168.]
LUO TX,LUO J,PANY D,2005. Leaf traits and associated ecosystem characteristics across subtropical and timberline forests in the Gongga Mountain,Eastern Tibetan Plateau[J]. Oecologia,142(2):261-273.
MARCELO DB,JORGE LF,LAURA LR,et al.,2009. Structural and functional changes in Nothofagus pumilio forests along an altitudinal gradient in Tierra del Fuego,Argentina[J]. J Veg Sci,11(2):l79-188.
MILLER-RUSHING AJ,PRIMACK RB,TEMPLER PH,et al.,2009. Long-term relationships among atmospheric CO2,stomata,and intrinsic water use efficiency in individual trees[J]. Am J Bot,96(10):1779-1786.
REICH P B,OLEKSYN J,MODRZYNSKI J,et al.,1996. Evidence that longer needle retention of spruce and pine populations at high elevations and high latitudes in largely a phenotypic response[J]. Tree Physiol,16(1):643-645.
SMITH WK,HINCKLEY TM,1995. Eeophysiology of coniferous forest[M]. San Diego:Academic Press:225-254.
TAKAHASHI K,MIYAJIMA Y,2008. Relationships between leaf life span,leaf mass per area,and leaf nitrogen cause different altitudinal changes in leaf δ13C between deciduous and evergreen species[J]. Botany,86(11):1233-1241.
WRIGHT I J,REICH P B,WESTOBY M,et al.,2004. The worldwide leaf economics spectrum[J]. Nature,428(6985):821-827.
XU B,WANG JN,SHI FS,et a1.,2013. Morphological adaptation of wild Fritillaria unibracteata to alpine conditions in the eastern Qinghai-Tibet Plateau[J]. Chin J Plant Ecol,37(3):187-196.[徐波,王金牛,石福孫,等,2013. 青藏高原东缘野生暗紫贝母生物量分配格局对高山生态环境的适应[J].植物生态学报,37(3):187-196.]
XU GJ,1997. Colored illustrations of Chinese traditional and herbal drugs[M]. 2nd ed. Fuzhou:Fujian Science and Technology Press:210-213.[徐国钧,1997. 中草药彩色图谱(修订本)[M]. 2版. 福州:福建科学技术出版社:210-213.]
YUN HX,CHEN Z,2009. Analysis of chemical composition of Fritillaria unibiacteata by GC- MS[J]. J Qinghai Med Coll,30(2):133-135.[韵海霞,陈志,2009. 暗紫贝母乙醚萃取物的GC-MS分析[J]. 青海医学院学报,30(2):133-135.]
ZHANG ML,2009. How to calculate the field area under a microscope[J]. Chin J Diagn Pathol,16(1):79-79.[张淼丽,2009. 如何计算显微镜下视野面积[J]. 诊断病理学杂志,16(1):79-79.]
ZHU DN,JIANG Y,CHEN T,et al.,1992. Research on chemical components and primary pharmaceutical actions of cultured bulb of Fritillaria unibracteata[J]. J Chin Pharm Univ,23(2):118-121.[朱丹妮,蒋莹,陈婷,等,1992. 组织培养川贝母化学成分和药理作用的研究[J]. 中国药科大学学报,23(2):118-121.]
(责任编辑 李 莉)
