乐昌含笑不同家系的叶形态与生长差异分析
2021-10-13梁龙胡德活王润辉郑会全晏姝曾碧欣韦如萍
梁龙, 胡德活, 王润辉, 郑会全, 晏姝, 曾碧欣, 韦如萍*
乐昌含笑不同家系的叶形态与生长差异分析
梁龙1, 胡德活2, 王润辉2, 郑会全2, 晏姝2, 曾碧欣2, 韦如萍2*
(1. 肇庆市国有大坑山林场,广东 肇庆 526400;2. 广东省森林培育与保护利用重点实验室,广东省林业科学研究院,广州 510520)
为了解乐昌含笑()叶形态和生长性状的家系变异特征,对15个家系的叶形态性状和生长指标及其相关性进行了分析。结果表明,乐昌含笑叶片形态在家系间和家系内均存在显著差异,不同家系间以帽子峰家系叶形态的平均变异系数最大(23.55%),乐九5家系的最小(12.63%);不同叶形态性状间以干物质质量的变异系数最大(25.50%),叶柄长宽比的最小(5.14%);树冠浓密度、胸径和树高在家系内的变异比家系间的显著。相关分析表明,叶长与叶宽、叶干物质质量与叶面积之间均呈显著正相关关系(<0.05)。根据聚类分析结果,15个家系可分为4类,第I类家系具有树冠浓密、树体高大等特征;第II、III类家系具有树冠较稀疏且树体较矮小等特征;第IV类家系具有树冠较稀疏但树体较高大等特征。因此,这为乐昌含笑树种的合理开发利用提供了理论依据。
乐昌含笑;家系;叶;形态;生长
乐昌含笑()又名南方白兰花、广东含笑、景烈含笑等,是华南地区优良的乡土阔叶树种,其材质优良、树干挺拔、花香四溢,可用于城市园林绿化,是营造风景林和生态公益林的主要树种,具有重要的经济价值和生态价值[1]。对乐昌含笑的研究近十余年来进展较快,已在群落特征[2]、种源家系选择[3–4]、育苗和栽培技术[5–6]、园林绿化[7–8]等方面取得较大成果。近些年各地积极践行“绿水青山就是金山银山”的生态理念,包括乐昌含笑在内的一大批优良乡土阔叶树种得到大力推广应用,因而对其生物学特性的研究也受到重视[9]。叶片是植物进行光合作用、蒸腾作用、气体交换功能的主要场所,也是植物感知外界光照、温度、水分等环境变化的重要器官[10]。叶片的性状特征,不仅体现了其系统发生和遗传信息[10],还是植物内在生理行为的外在展示以及个体适应环境变化所形成的一种生存对策,对植物的基本行为和功能具有重要影响[9,11]。因此,对植物叶片性状的研究已成为当前植物生态学等领域的热点[9],但目前还尚未见关于乐昌含笑家系叶片形态特征的研究报道。本文通过对多年生乐昌含笑家系试验林中15个家系叶片形态特征和生长情况的调查测定,分析了不同家系的叶形态差异及其对植株生长的影响, 以期为乐昌含笑优良基因资源的挖掘和利用提供参考。
1 材料和方法
1.1 试验地概况
乐昌含笑家系试验林位于广东省肇庆市国有大坑山林场,北纬23º58ʹ,东经112º10ʹ,海拔高度120~200 m,坡度35º,年均温20.9℃,年均积温6 520.5℃,年均降水量1 770 mm,试验林土壤为黄岗岩成土,红壤,土层深1.1 m以上。2005年3月造林,株行距3 m×3 m,初植密度1 110 ind./hm2, 4株小区,8次重复。
1.2 材料
在广东省北部和江西省南部乐昌含笑自然分布区进行乐昌含笑优树选择,于2003年分单株(家系)采种,2004年培育容器苗,2005年造林。参试家系共15个,包括来自广东省乐昌县大源镇的乐大1、乐大6、乐大7家系,乐昌县九峰镇的乐九3、乐九4、乐九5、乐九7,乐昌县两江镇的乐两3A、乐两4、乐两6、乐两12、乐两14、乐两15、乐两16,以及来自南雄市帽子峰镇的帽子峰家系。
1.3 方法
于2019年11月,测量参试家系所有重复的植株胸径和树高,然后从每个家系中选择3株胸径接近家系内群体平均值的植株,采集植株中下部侧枝中部的叶片,每株随机采集100片叶,要求叶形完整,叶色正常。叶片采集后放入封口袋中带回实验室。叶长指叶基部(不含叶柄)至叶尖的长度,叶宽指叶中部的宽度。采用Expression 11000XL (EPSON)扫描叶片后用万深LA-S叶面积分析系统获取叶形态参数指标。然后,所有叶片样品先105℃杀青30 min,再在85℃下烘至恒重,称量每片叶的干物质质量。树冠浓密度采用评分制:3分为浓密,2分为较浓密,1分为稀疏[3]。
1.4 数据的统计分析
利用Microsoft Excel 2010和SAS 8.1软件进行数据的统计分析。表型变异系数=/,为标准差,为平均数[12];相对极差ʹ=R/0×100%,R为第性状家系内极差,0为第性状的家系间总体极差[12]。
2 结果和分析
2.1 叶形态的变异分析
由家系间叶片形态指标多重比较结果(表1)可知,除叶柄长宽比在家系间的差异较小外,其余6个叶形态指标在家系间存在较大差异,其中,来自乐昌九峰镇的4个家系叶长、叶长宽比的变异较大,分别为10.52~14.15 cm和2.35~3.14;来自乐昌两江镇的7个家系叶宽、叶面积、干物质质量的变异较大,分别为3.63~5.23 cm、34.04~56.75 cm2和0.19~0.38 g; 乐昌大源镇的3个家系的比叶面积变异较大, 为128.93~ 149.41。表明乐昌含笑叶片形态在不同来源地的家系间和同一来源地的家系间均存在明显差异。
乐昌含笑家系内叶片形态指标的变异系数不同(表2),以帽子峰的最大(23.55%),乐九5的最小(12.63%)。叶片形态指标在不同家系间的变异系数也存在较大差异,为5.14%~25.50%,以叶柄长宽比的最小,叶干物质质量的最大。
2.2 生长特征的变异分析
从表3可知,除胸径在家系间有差异外,树高和树冠浓密度的差异不显著;但在家系内,3个指标的变异幅度均较大,分别为24.38%~42.55%、25.33%~39.88%和31.02%~44.41%。15个家系的平均胸径为9.76 cm、平均树高为7.71 m、树冠浓密度均值为1.61。乐九4、乐两12、乐两15、帽子峰4个家系生长较好,胸径为11.07~11.27 cm,树高为8.07~8.48 m,树冠浓密度为1.64~1.93;而乐大6、乐九7、乐两16、乐两6家系生长较差,胸径为8.11~8.94 cm,树高为6.35~7.42 m,树冠浓密度为1.45~1.67。表明乐昌含笑生长指标在家系间和家系内均存在较大变异,并以家系内的变异更明显。
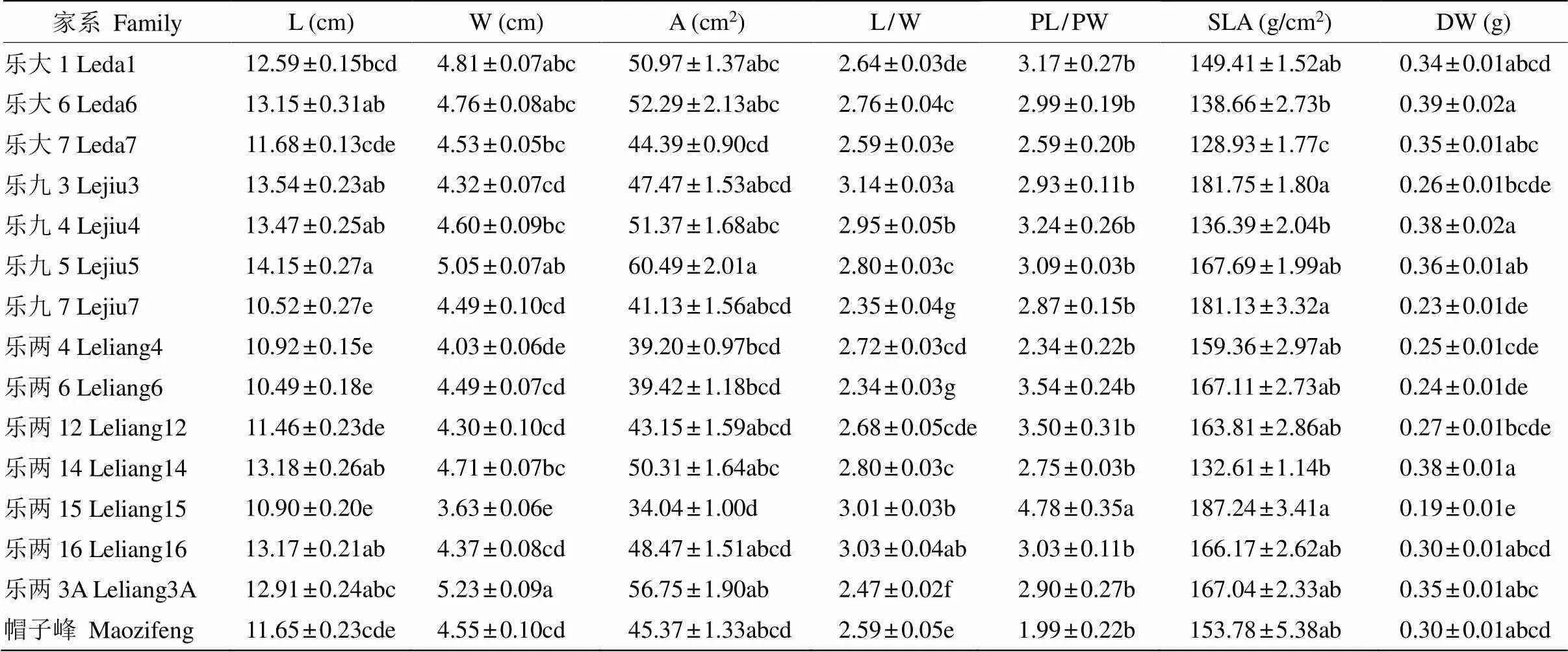
表1 乐昌含笑家系叶形态指标的多重比较
同列数据后不同字母表示差异显著(<0.05)。L: 长; W: 宽; A: 面积; PL/PW: 叶柄长宽比; SLA: 比叶面积; DW: 干物质质量。下表同。
Different letters within column indicate significant differences at 0.05 level. L: Length; W: Width; A: Area; PL/PW: Petiole length-width ratio; SLA: Specific leaf area; DW: Dry weight. The same is following Tables.
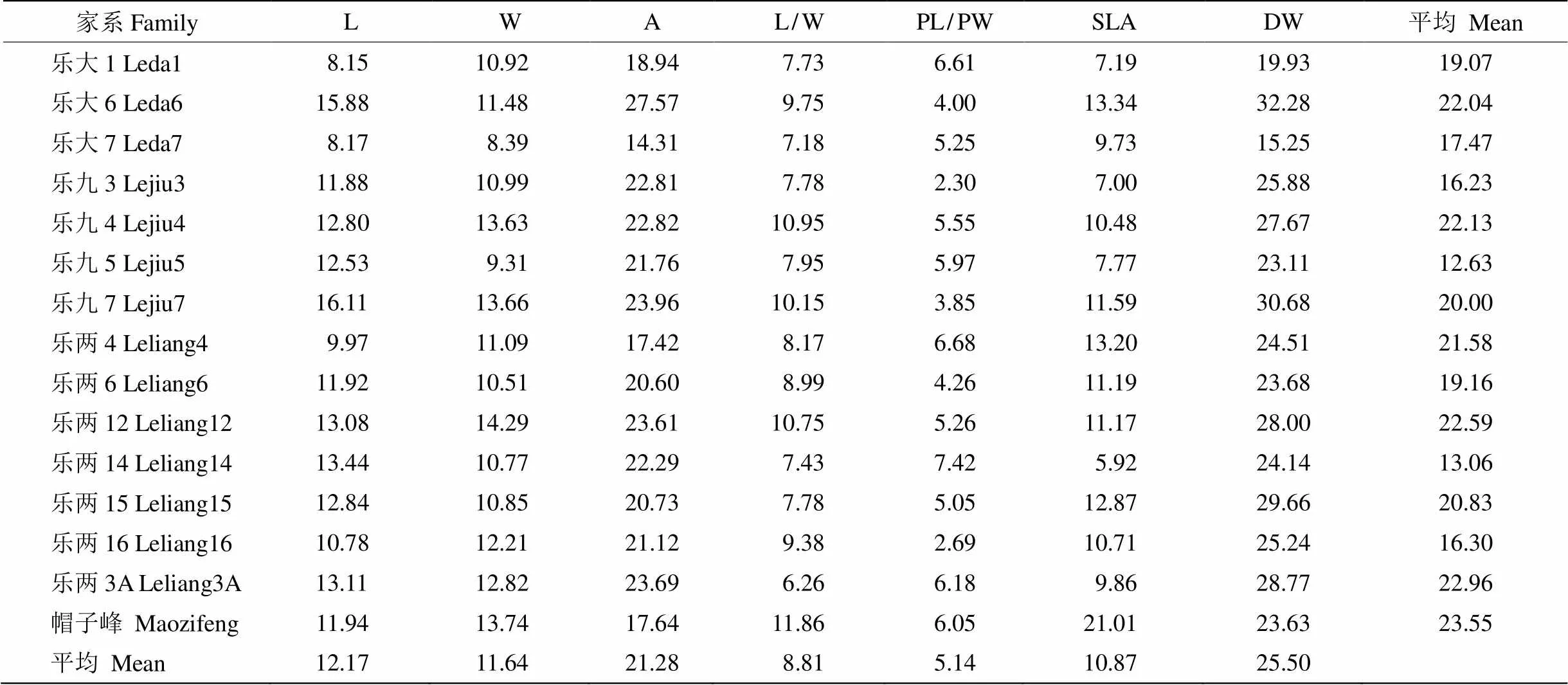
表2 乐昌含笑叶形态指标的变异系数(%)
2.3 相关性分析
15个参试家系形态和生长指标的偏相关分析结果表明(表4),叶长与叶宽呈显著正相关,与叶长宽比和叶面积呈极显著正相关;叶宽与叶长宽比呈极显著负相关,与叶面积呈极显著正相关。叶干物质质量与叶面积呈显著正相关,与比叶面积呈极显著负相关。由此可见,7个叶片形态指标与树冠浓密度、胸径和树高3个指标间均未有显著相关性,但叶面积和叶柄长宽比对树冠浓密度有积极影响,且叶宽与胸径间有一定的负相关趋势,与树高间有一定的正相关趋势。
2.4 聚类分析
对15个家系进行多指标系统的聚类分析,结果表明(图1),在阈值为1.00时,可把15个家系分为4个类别。第I类家系只有乐两15,其叶面积较小、叶柄细长、叶片短细呈窄倒卵形、树冠浓密且树体高大;第II类家系有乐九7、乐两4和乐两6,具有叶面积较小、叶片短宽呈短圆状倒卵形、树冠较稀疏且树体较矮小等特征;第III类家系有乐大6和乐两16, 具有叶面积较大、叶片细长呈窄倒卵形、树冠较稀疏且树体矮小等特征;第IV类家系有乐大1、乐大7、乐九3、乐九4、乐九5、乐两12、乐两14、乐两3A、帽子峰共9个家系,具有叶面积较大、叶片细长呈窄倒卵形、树冠较稀疏但树体较高大等特征。
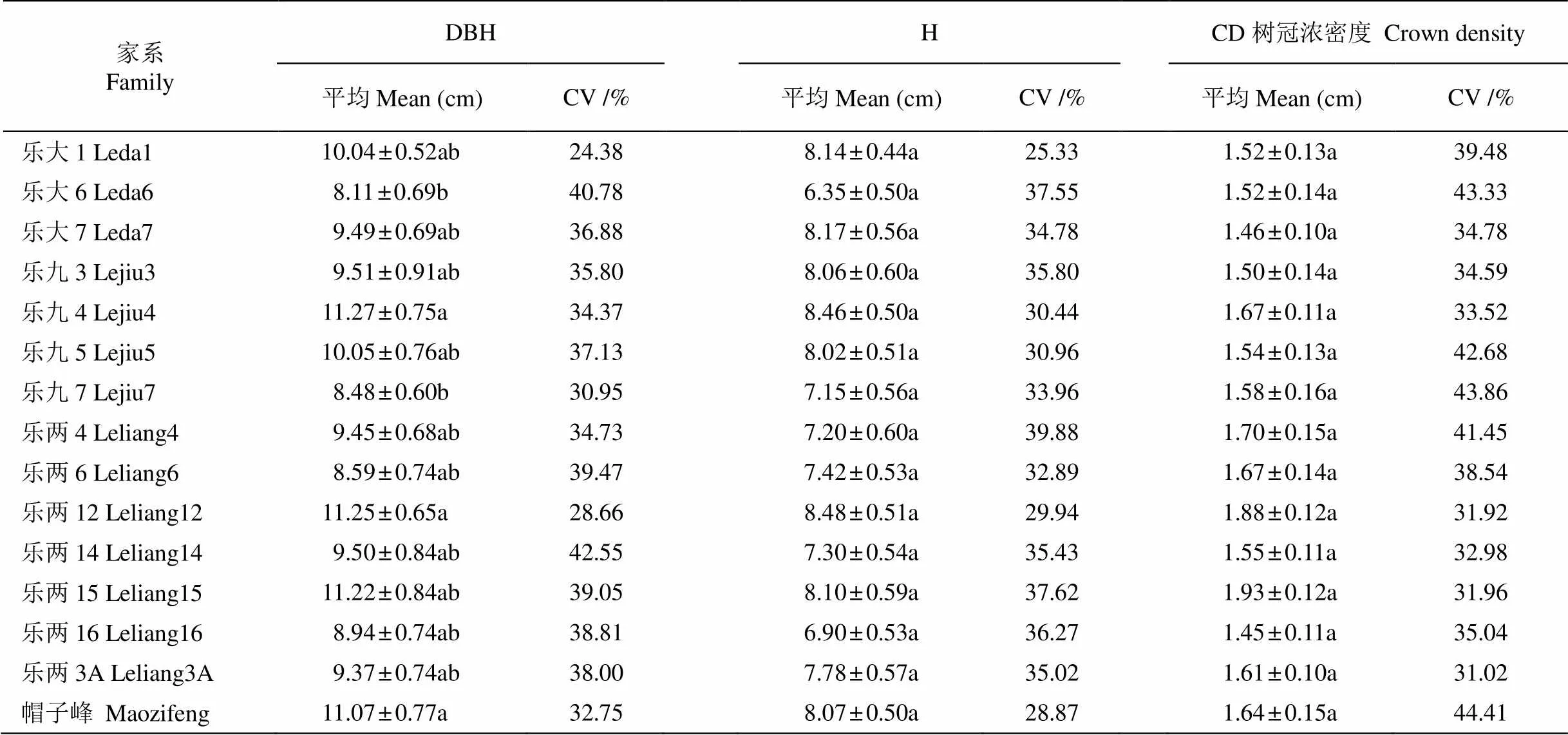
表3 乐昌含笑不同家系生长差异分析
DBH: 胸径; H: 树高; CD: 树冠浓密度; CV: 变异系数。下表同。
DBH: Diameter at breast height; H: Tree height; CD: Crown density; CV: Coefficient of variation. The same is following Tables.
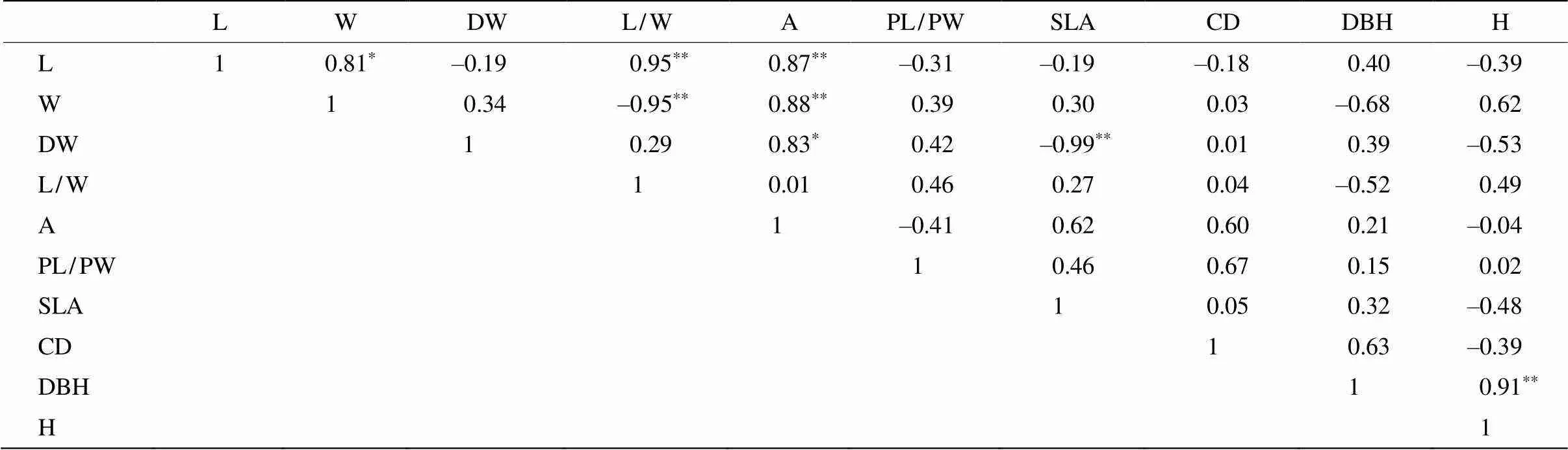
表4 乐昌含笑家系叶形态和生长指标的偏相关系数
**:<0.01; *:<0.05.
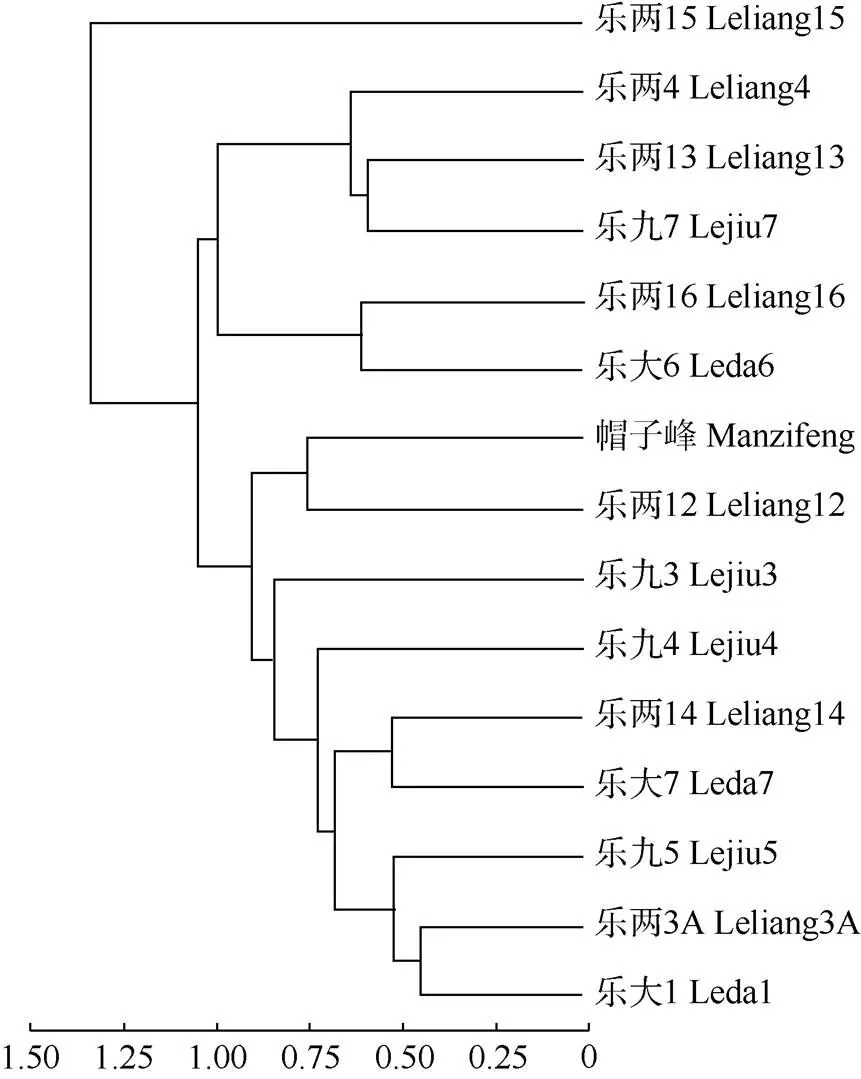
图1 乐昌含笑家系的表型聚类图
3 结论和讨论
植物叶片的发育由叶原基起始,期间涉及转录因子、小分子RNA及生长调节剂参与的一系列细胞分裂和分化程序,最终发育成形态大小固定的成熟叶[13];整个发育过程是植物内因和外部环境因素共同作用的结果,因而即使遗传背景相同,同一株树上也长不出两片完全相同的叶子[12–14]。由15个乐昌含笑家系叶片形态分析结果可知,叶长、叶宽、叶面积、叶长宽比、比叶面积、叶干物质质量在不同来源地和同一来源地的家系间和家系内均存在明显差异。由此推测,位于同一试验林内的15个家系,其叶片形态发育过程除了受到外部环境因素的影响外,可能更多的是内部遗传因素的调控。由于本试验限于一片家系试验林,今后还应通过多个试验点数据的系统分析,以深入解析乐昌含笑家系叶形态发育与环境的互作效应。
叶的基本功能是光合作用和蒸腾作用[15],叶面积越大,叶片对光的吸收量得到增强,叶片的光合效率提高,对植株接收和利用光能有重要意义[16]。蔡锡安等研究表明,低光下梅叶冬青()的叶厚度变小,而叶长、叶宽、比叶面积及单叶面积明显增大,并指出叶片形态结构的改变是梅叶冬青对低光的一种适应策略[17]。乐昌含笑属于喜光树种,在温暖、湿润、土壤肥沃、阳光充足的环境中生长良好[18]。本研究对15个乐昌含笑家系的10个形态和生长指标的相关性分析表明,叶宽与胸径间呈现出较密切的负相关趋势,与树高呈正相关关系(表4),即叶片越宽,植株高生长可能越明显,胸径生长则越小。这可能也暗示了试验林中存在着较激烈的个体竞争,为了获取更好的光照条件,植株更趋于向上层空间伸展。
乐昌含笑是兼具景观和用材特性的优良乡土阔叶树种,基于多个指标的聚类分析结果,可对15个乐昌含笑家系进行合理应用,对于第I类家系的乐两15, 第IV类家系的乐大1、乐大7、乐九3、乐九4、乐九5、乐两12、乐两14、乐两3A、帽子峰等,可在林业生态建设和城市园林景观建设中应用。第I类家系乐两15具有树体高大、树冠浓密等特性,尤其适合在城市(道路)绿化美化中单植、列植或群植;第IV类家系因具有树体高大、树冠较稀疏等特性,可在相似区域的工业用材林基地中应用;对于第II、III类家系,可作为乐昌含笑基因资源进行适当保存。当然,由于本研究局限于单一地点,今后还应扩展区域试验或适应性试验,以充分了解其形态变异特征,尤其是生长特性和适生性后再大面积的推广应用,以期为乐昌含笑这一优良乡土阔叶树种的合理开发提供科学依据。
[1] ZHANG W H, WANG R H, ZHEGN H Q, et al. Multi-site family trials and selection of superior families for[J]. J Zhe- jiang Agric For Univ, 2015, 32(5): 763–769. doi: 10.11833/j.issn.2095- 0756.2015.05.016.
张伟红, 王润辉, 郑会全, 等. 乐昌含笑优树多点子代测定及优良家系选择 [J]. 浙江农林大学学报, 2015, 32(5): 763–769. doi: 10. 11833/j.issn.2095-0756.2015.05.016.
[2] MA L, WU L F, YE H G, et al. Studies on community characteristics ofin Yejingbei, Zengcheng, Guangzhou [J]. J Trop Subtrop Bot, 2012, 20(2): 169–176. doi: 10.3969/j.issn.1005-3395.2012. 02.011.
马磊, 吴林芳, 叶华谷, 等. 广州增城叶迳背乐昌含笑林群落特征研究 [J]. 热带亚热带植物学报, 2012, 20(2): 169–176. doi: 10.3969/ j.issn.1005-3395.2012.02.011.
[3] WANG R H, ZHANG W H, ZHEGN H Q, et al. Study on mutil-site provenance trials and superior provenances selection of[J]. J CS For Technol, 2015, 35(5): 16–21,34. doi: 10.14067/ j.cnki.1673-923x.2015.05.003.
王润辉, 张伟红, 郑会全, 等. 乐昌含笑多点种源试验与优良种源选择 [J]. 中南林业科技大学学报, 2015, 35(5): 16–21,34. doi: 10. 14067/j.cnki.1673-923x.2015.05.003.
[4] ZOU S M, YAN S, WANG R H, et al. Study on early trials ofdandy families [J]. Guangdong For Sci Technol, 2009, 25(2): 10–15. doi: 10.3969/j.issn.1006-4427.2009.02.002.
邹寿明, 晏姝, 王润辉, 等. 乐昌含笑家系试验初报 [J]. 广东林业科技, 2009, 25(2): 10–15. doi: 10.3969/j.issn.1006-4427.2009.02.002.
[5] ZHANG W H, YAN S, WANG R H, et al. Grafting techniques for plus tree ofDandy [J]. Mod Agric Sci Technol, 2018(6): 147–148. doi: 10.3969/j.issn.1007-5739.2018.06.094.
张伟红, 晏姝, 王润辉, 等. 乐昌含笑优树嫁接技术 [J]. 现代农业科技, 2018(6): 147–148. doi: 10.3969/j.issn.1007-5739.2018.06.094.
[6] LIE Z Y, LI J, ZHOU T T, et al. Density effect on nutrient content and accumulation ofplantations [J]. Hunan For Sci Technol, 2016, 43(1): 44–47,60. doi: 10.3969/j.issn.1003-5710.2016. 01.008.
列志旸, 李洁, 周彤彤, 等. 密度对乐昌含笑幼苗养分积累的影响 [J]. 湖南林业科技, 2016, 43(1): 44–47,60. doi: 10.3969/j.issn.1003- 5710.2016.01.008.
[7] LIU S Z, XUE K N, KONG G H, et al. Effects of air pollution on the growth of 35 garden plants [J]. J Trop Subtrop Bot, 2003, 11(4): 329– 335. doi: 10.3969/j.issn.1005-3395.2003.04.004.
刘世忠, 薛克娜, 孔国辉, 等. 大气污染对35种园林植物生长的影响[J]. 热带亚热带植物学报, 2003, 11(4): 329–335. doi: 10.3969/j. issn.1005-3395.2003.04.004.
[8] QIU E F, XU F, WANG C, et al. Population distribution and structure characteristics of village roadside forest in Fujian Province, eastern China [J]. J Beijing For Univ, 2012, 34(6): 68–74. doi: 10.13332/j. 1000-1522.2012.06.023.
邱尔发, 许飞, 王成, 等. 福建省乡村道路林种群分布及结构特征研究 [J]. 北京林业大学学报, 2012, 34(6): 68–74. doi: 10.13332/j. 1000-1522.2012.06.023.
[9] XUE L, ZHANG R, XI R C, et al. Seasonal change of leaf morpho- logical traits of six broadleaf seedlings in South China [J]. Acta Ecol Sin, 2012, 32(1): 123–134. doi: 10.5846/stxb201011291691.
薛立, 张柔, 奚如春, 等. 华南地区6种阔叶幼苗叶片形态特征的季节变化 [J]. 生态学报, 2012, 32(1): 123–134. doi: 10.5846/stxb 201011291691.
[10] KE J X, CHEN D, GUO Y P. Designing leaf marginal shapes: Regula- tory mechanisms of leaf serration or dissection [J]. Biodiv Sci, 2018, 26(9): 988–997. doi: 10.17520/biods.2018127.
柯锦秀, 陈多, 郭延平. 植物叶缘形态的发育调控机理 [J]. 生物多样性, 2018, 26(9): 988–997. doi: 10.17520/biods.2018127.
[11] WANG Y L, WANG W F, ZHANG Y X, et al. Responses of leaf morphological structure and physiological characteristics ofcv. ‘BYu’ to drought stress [J]. Sci Silv Sin, 2019, 55(4): 42–50. doi: 10.11707/j.1001-7488.20190405.
王怡霖, 王卫锋, 张芸香, 等. 碧玉杨叶形态结构与生理特性对干旱的响应[J]. 林业科学, 2019, 55(4): 42–50. doi: 10.11707/j.1001- 7488.20190405.
[12] YAN D T. Leaf morphological differences of(Ulmaceae) [D]. Linfen: Shanxi Normal University, 2017: 1–53.
闫冬婷. 大果榆叶形态差异研究 [D]. 临汾: 山西师范大学, 2017: 1–53.
[13] LE L N, HUANG M R, CHEN Y. The research progress on molecular regulation of leaf morphogenesis [J]. Mol Plant Breed, 2016, 14(11): 3205–3213. doi: 10.13271/j.mpb.014.003205.
乐丽娜, 黄敏仁, 陈英. 植物叶形态建成的分子机理研究进展 [J]. 分子植物育种, 2016, 14(11): 3205–3213. doi: 10.13271/j.mpb.014. 003205.
[14] RONG W T, XING F W, YI Q F. Effects of climatic factors on leaf discoloration peak period of 5 species in Macao [J]. J Trop Subtrop Bot, 2020, 28(6): 574–582. doi: 10.11926/jtsb.4232.
容文婷, 邢福武, 易绮斐. 澳门5种植物叶变色盛期对气候因子的响应[J]. 热带亚热带植物学报, 2020, 28(6): 574–582. doi: 10. 11926/jtsb.4232.
[15] WANG S, YU Y M, ZHOU Z L, et al. Different clones ofBureau simple leaves morphological characteristics growth dynamic and correlation studies [J]. J Gansu Agric Univ, 2019, 54(5): 135–139. doi: 10.13432/ j.cnki.jgsau.2019.05.017.
王桑, 于永明, 周卓玲, 等. 灰楸不同无性系单叶形态特征生长动态及其相关性 [J]. 甘肃农业大学学报, 2019, 54(5): 135–139. doi: 10.13432/j.cnki.jgsau.2019.05.017.
[16] HE M H, LIU Z W, CHEN W N. Study on morphology and structure of leaves ofBail [J]. Anhui Agric Sci Bull, 2018, 24(13): 94–96. doi: 10.16377/j.cnki.issn1007-7731.2018.13.045.
何明惠, 刘中伟, 陈文年. 珙桐叶形态结构研究 [J]. 安微农学通报, 2018, 24(13): 94–96. doi: 10.16377/j.cnki.issn1007-7731.2018.13.045.
[17] CAI X A, RAO X Q, LIU Z F, et al. Effects of shading on leaf morphology, photosynthetic characteristics, and growth of[J]. J Trop Subtrop Bot, 2020, 28(1): 25–34. doi: 10.11926/jtsb.4077.
蔡锡安, 饶兴权, 刘占锋, 等. 遮荫处理对梅叶冬青叶片形态、光合特性和生长的影响 [J]. 热带亚热带植物学报, 2020, 28(1): 25–34. doi: 10.11926/jtsb.4077.
[18] ZHANG F Q, LI X Q, PAN W, et al. Cultivation Techniques of Ecological Landscape Trees in Guangdong Province [M]. Beijing: China Forestry Press, 2012: 95–97.
张方秋, 李小川, 潘文, 等. 广东生态景观树种栽培技术 [M]. 北京: 中国林业出版社, 2012: 95–97.
Analysis of Leaf Morphology and Growth Differences amongFamilies
LIANG Long1, HU Dehuo2, WANG Runhui2, ZHENG Huiquan2, YAN Shu2, ZENG Bixing2, WEI Ruping2*
(1. Dakengshan Forest Farm,Zhaoqing 526400, Guangdong, China; 2. Guangdong Provincial Key Laboratory of Silviculture, Protection and Utilization, Guangdong Academy of Forestry,Guangzhou 510520, China)
In order to explore the variability of leaf characters and growth traits among different families of, the leaf morphology, growth indexes and their correlations of 15 families were analyzed. The results showed that there were significant differences in leaf characters amongfamilies. The coefficient of variation (CV) of leaf characters in Maozifeng family was the largest (23.55%) among 15 families, and that in lejiu5 family was the smallest (12.63%). Among seven leaf characters, the CV of dry matter mass was the largest (25.50%), and that of petiole length-width ratio was the smallest (5.14%). The variation of crown density, DBH and tree height (H) within families was more significant than that between families. The correlation analysis showed that leaf length had significant positive correlation with leaf width (<0.05),as well as the correlation between leaf dry matter mass and leaf area. According to cluster analysis, 15 families could be divided into 4 categories. The families in category I had dense crown and tall tree, those in categories II and III had sparse crown and short tree, and those in category IV had sparse crown but tall tree. Therefore, these would provide a theoretical basis for the rational development and utilization of.
; Family; Leaf; Phenotype; Growth
10.11926/jtsb.4331
2020-11-03
2021-01-07
中央财政林业科技推广示范项目([2019]GDTK-03)资助
This work was supported by the Project for Forestry Science and Technology Promotion Demonstration of Central Finance (Grant No. [2019]GDTK-03).
梁龙(1978~ ),男,硕士,林业工程师,主要从事森林培育研究和林业管理工作。E-mail: vonlong2008@163.com
. E-mail: wrpgx@163.com
