亚热带常绿阔叶林89种木本植物一级根碳氮浓度变异规律
2021-10-13王雪闫晓俊范爱连贾林巧熊德成黄锦学陈光水姚晓东
王雪, 闫晓俊,2, 范爱连,贾林巧, 熊德成, 黄锦学, 陈光水, 姚晓东*
亚热带常绿阔叶林89种木本植物一级根碳氮浓度变异规律
王雪1, 闫晓俊1,2, 范爱连1,贾林巧1, 熊德成1, 黄锦学1, 陈光水1, 姚晓东1*
(1. 福建师范大学地理科学学院/湿润亚热带生态地理过程教育部重点实验室, 福州 350007; 2. 江苏省木渎高级中学, 江苏 苏州 215101)
为了解亚热带地区常绿阔叶林木本植物一级根养分元素变化规律,基于根序法对福建省建瓯市万木林自然保护区天然常绿阔叶林群落的89种树种一级根进行碳、氮浓度测定。结果表明,89种树种一级根的C、N质量浓度分别为433.9 和13.7 mg/g, C∶N为36.7,变异系数分别为6.4%、39.2%和39.9%;一级根C浓度在叶片习性和生长型间存在显著差异,而N浓度和C∶N在不同叶片习性和生长型间的差异不显著;6主要科[樟科(Lauraceae)、壳斗科(Fagaceae)、冬青科(Aquifoliaceae)、山矾科(Symplocaceae)、五列木科(Pentaphylacaceae)和杜英科(Elaeocarpaceae)]树种间一级根C、N浓度和C∶N均差异显著;一级根N浓度随物种系统发育由低级向高级呈现增加的趋势。因此,亚热带常绿阔叶林一级根C浓度种间变异低于N浓度;一级根N浓度受系统发育的影响,而C浓度则受叶片习性和生长型影响,表现出一定的趋同效应。
吸收根;化学计量;变异特征;天然林
细根作为决定植物生长、存活的重要地下器官[1],在获取、运输、存储营养物质和水分方面具有重要作用[2–4],对生物地球化学循环具有重要意义[5–7]。细根化学计量学性状在细根代谢、分解、土壤微生物活性以及土壤碳输入方面起着不可替代的作用[8–9]。因此,开展细根化学计量的研究,将有助于加深对植物养分获取、元素限制等过程的理解。根据根功能模块的划分,细根被分为吸收根和运输根,位于根系系统中最远端的根(即一级根,为吸收根)[3–4],具有快速的周转率和高的代谢活性,对水分和养分吸收更灵敏[4,10],因此对植物一级根化学性状进行研究更为重要。
目前关于细根化学计量学已在各尺度开展了大量研究。Liu等[11]对中国亚热带地区16优势种细根功能性状变异的研究表明,树种间细根的C、N浓度和C∶N差异显著;张涛等[12]报道摩天岭地区草本植物根系C浓度的变异系数小于N浓度和C∶N,刘璐等[13]的研究表明,27种亚热带植物细根的C浓度变异系数低于N浓度。Geng等[14]报道不同地理区域(温带内蒙古草地和青藏高寒草地)的草地物种细根的C、N、P浓度没有差异,但细根N浓度和N∶P在5种草原类型间有显著差异; Chen等[15]的研究表明亚热带和温带被子植物间的一级根N浓度没有显著差异;Wang等[16]报道,中国东部南北横断面上的181种木本、非木本物种间C、N浓度及C∶N无显著差异。Yuan等[17]通过分析全球200多项研究的植物根性状数据,认为细根的N∶P在不同生物群落间存在显著差异;而Ma等[18]搜集全球369种物种的一级根数据,认为一级根N浓度在不同生物群落间没有差异。虽然已有研究从局地、区域和全球尺度探讨了植物细根化学计量学特征,但绝大多数研究中细根以直径≤2 mm定义, Guo等[19]和McCormack等[4]认为这种划分未能充分考虑不同植物间细根分枝在形态、结构和功能上的等级差异,导致种间的可比性存在问题,同时使结果存在较大不确定性。此外,同一立地的研究样本通常只有10多种,很难说明同一森林群落中木本植物一级根C、N变异的基本规律。亚热带地区具有较好的光、水、热等资源条件,森林资源比较丰富,木本植物类型多样,但是该区木本植物一级根的化学性状变异程度与跨区域和全球尺度的研究相比,处于哪个区间位置还不清楚。
最近对不同叶片习性和生长型的植物根系化学计量学性状的研究表明,亚热带地区59种常绿和落叶树种一级根C浓度无差异,而落叶树种的N浓度显著高于常绿树种[20];延河流域的灌木、半灌木的根N浓度显著高于乔木[21];亚热带地区的天然米槠()林乔木层与灌木层树种根系C、N、P浓度没有差异[22];王钊颖等[23]的研究表明乔木与灌木细根N、P浓度和N∶P没有差异。可见,目前同一地区不同叶片习性、生长型物种的根C、N变异研究仍主要以细根为主,且对常绿与落叶树种、乔木与灌木树种根化学性状变异还不够深入。此外,科水平的植物生态化学计量学研究还主要集中于草本植物[24]且常以不同功能群组进行区分[25–26],而对多个科水平的木本植物细根特别是一级根的研究较为少见[27]。相同立地的植物常受到趋同效应的影响,不同科之间的根系在同一立地上化学元素含量如何变化还不是很明确。近年来,系统发育结构被认为是影响根功能性状变异的重要原因,对温带草原少量物种的研究表明,吸收根只有N浓度表现出显著的系统发育信号[28];且系统发育关系对植物细根C和N浓度具有显著影响[10,29–30];但Ma等[18]却认为,系统发育对一级根C、N浓度和C∶N没有影响。可见,系统发育关系研究主要集中在同一立地的少数草本植物或者大尺度的全球性木本、非木本植物,对同一地区大量木本植物影响的研究还不多。
本研究在亚热带天然常绿阔叶林典型地段上采集89种树种的根系样品,根据叶片习性、生长型、科系进行分类,测量一级根的C、N浓度,计算C∶N,旨在从多个角度揭示亚热带地区木本植物一级根化学计量学特征,并认识亚热带植物一级根C、N浓度、C∶N变异在全球细根化学元素变异的区间位置。
1 材料和方法
1.1 研究区概况
研究区位于福建省建瓯市房道镇境内的万木林自然保护区(27.05° N,118.15° E),处于武夷山脉东南侧,总面积189 hm2。该区属于亚热带季风气候,年均降水量1 663.8 mm,年均温18.7℃,相对湿度81%,无霜期为277 d。地貌类型为低山丘陵;地带性土壤以红壤和黄壤为主。主要森林植被类型为亚热带常绿阔叶林,维管植物种类丰富,以壳斗科(Fagaceae)、樟科(Lauraceae)、冬青科(Aquifoliaceae)等植物为主。
1.2 根系样品采集与处理
于2018年6、7月进行根系采样,在保护区常绿阔叶林群落内选择典型地段采集89种木本植物(表1)根系,每树种采集3棵(胸径或地径相近),共计267棵。采用完整土块法[31]进行根系挖掘,即在离树干基部一定距离挖掘1~2个长、宽、高各20 cm的土块,距乔木基部约1.2 m,距小乔木基部约0.5 m,灌木贴近基部。通过与主根相连的侧根从根的颜色、表皮质地、根直径和根的构型进行比对, 在野外迅速挑出土块内的目标根系。将根系用湿纱布包裹后装进自封袋,贴好标签,置于放有冰块的冰盒内(<5℃)暂时保存。冰盒于3 h内带回实验室, 将根系样品放于–20℃的冰箱中冷冻保存,待采样完成后统一进行清洗和根系分级。
1.3 一级根C、N浓度和C∶N的测定
细根分级采用Pregitzer等[3]的根序法,位于根系分支最末端的细根为一级根。先将一级根在65℃烘干,用球磨仪进行粉碎。每树种(3个重复)的一级根样品粉末用锡杯精确称取8~10 mg,包样并记录质量。用CN元素分析仪(Elemental Analyzer) Vario EL III测定一级根的全C和全N。C∶N为一级根的全C与全N浓度的比值。
1.4 系统发育树的构建
在“植物智”网站(http://www.iplant.cn/frps)中“中国植物志”板块查询89种木本植物的学名,通过R 3.3.0中的“”程序包生成科属种列表,并提交给在线软件Phylomatic (http://phylodiversity.net/ phylomatic/),最后输出NEWICK树,该树以Zanne等[32]发表的系统发育进化树为骨架。本文构建的系统发育树见王雪等[33]的研究。
1.5 数据处理
利用Microsoft Excel 2013软件对木本植物一级根C、N浓度、C∶N化学计量学数据进行处理,计算平均值、中位值、极值和变异系数等。数据分析前进行以10为底的对数转换以符合正态分布的要求。利用SPSS 20.0软件中的单因素方差分析检验叶片习性、生长型和主要科系对一级根C、N浓度和C∶N的影响,用LSD进行多重比较(=0.05)。在R 3.3.0统计平台上利用“”程序包计算得到Blomberg’s值[34],检验天然常绿阔叶林一级根C、N浓度、C∶N的系统发育信号,同时用线性回归分析检验科水平分化时间与一级根N浓度的相关性。参照前人研究[35],将科内最早属的分化时间定义为该科的分化时间[15,18,36]。采用Origin 9.0和Microsoft Excel 2013软件绘制图表。
2 结果和分析
2.1 一级根化学计量学变异特征
89种木本植物的一级根C质量浓度为307.87~ 500.82 mg/g,平均(433.9±2.9) mg/g;N质量浓度为5.54~30.19 mg/g,平均(13.7±0.6) mg/g;C∶N为13.97~75.94,平均(36.7±1.6)。89种树种的C浓度分布比N浓度和C∶N更为集中,C浓度变异系数最小,为6.4%;而N浓度和C∶N接近40%,约为C浓度的6.2倍(图1)。
从叶片习性来看,常绿树种的一级根C质量浓度(346.30~500.82 mg/g)要大于落叶树种(307.87~ 457.45 mg/g);常绿与落叶树种一级根N质量浓度分别为5.54~28.97和5.84~30.19 mg/g;C∶N分别为37.85±1.7和31.90±3.7。常绿树种与落叶树种一级根C浓度数据分布比N浓度和C∶N相对集中,常绿树种一级根C浓度变异系数5.7%, 小于N浓度和C∶N的变异系数,落叶树种一级根C∶N的变异系数最大(48.5%),约是C浓度(8.0%)的6倍。常绿树种一级根C浓度显著高于落叶树种(<0.05), 而N浓度和C∶N无显著差异。
从生长型来看,灌木一级根C质量浓度最大(307.87~457.45 mg/g),乔木的N质量浓度最大(5.84~ 30.19 mg/g)。灌木一级根C浓度的变异系数最大(10.2%);乔木一级根N浓度的最高(41.1%),灌木最低(26.3%);一级根C∶N变异系数为乔木>小乔木或灌木>灌木。不同生长型一级根C浓度的差异显著,其中灌木一级根C浓度显著低于乔木、小乔木或灌木(<0.05),但N浓度和C∶N的差异不显著。这表明,亚热带常绿阔叶林树种一级根C浓度受叶片习性和生长型的影响,表现出一定的趋同适应。

表1 亚热带常绿阔叶林群落中的89种树种
由图2可见,6主要科一级根C浓度变异系数均低于N浓度和C∶N,冬青科的C质量浓度为346.30~445.30 mg/g,平均423.4 mg/g,变异系数最大,为8.2%;杜英科的C质量浓度为411.96~432.25 mg/g,平均422.4 mg/g,变异系数最小,为2.1%。樟科一级根的N质量浓度为6.15~28.97 mg/g, 平均19.4 mg/g,变异系数最大,为33.5%;五列木科平均仅9.4 mg/g,变异系数最小,为17.8%。一级根C∶N以樟科的变异系数最大(59.4%),其次是冬青科(43%),五列木科最小(15.7%)。6主要科间一级根C、N浓度、C∶N的差异显著,其中壳斗科一级根C质量浓度(448.56 mg/g)显著高于冬青科、山矾科和杜英科(<0.05);樟科一级根N质量浓度(19.39 mg/g)显著高于其他科(<0.05),其他5科间差异不显著;五列木科和壳斗科一级根C∶N最高,且显著高于樟科、冬青科和山矾科(<0.05)。这表明樟科一级根倾向于高C、N浓度,低C∶N,壳斗科一级根倾向于高C浓度和C∶N及低N浓度,山矾科一级根倾向于低C、N浓度和C∶N。
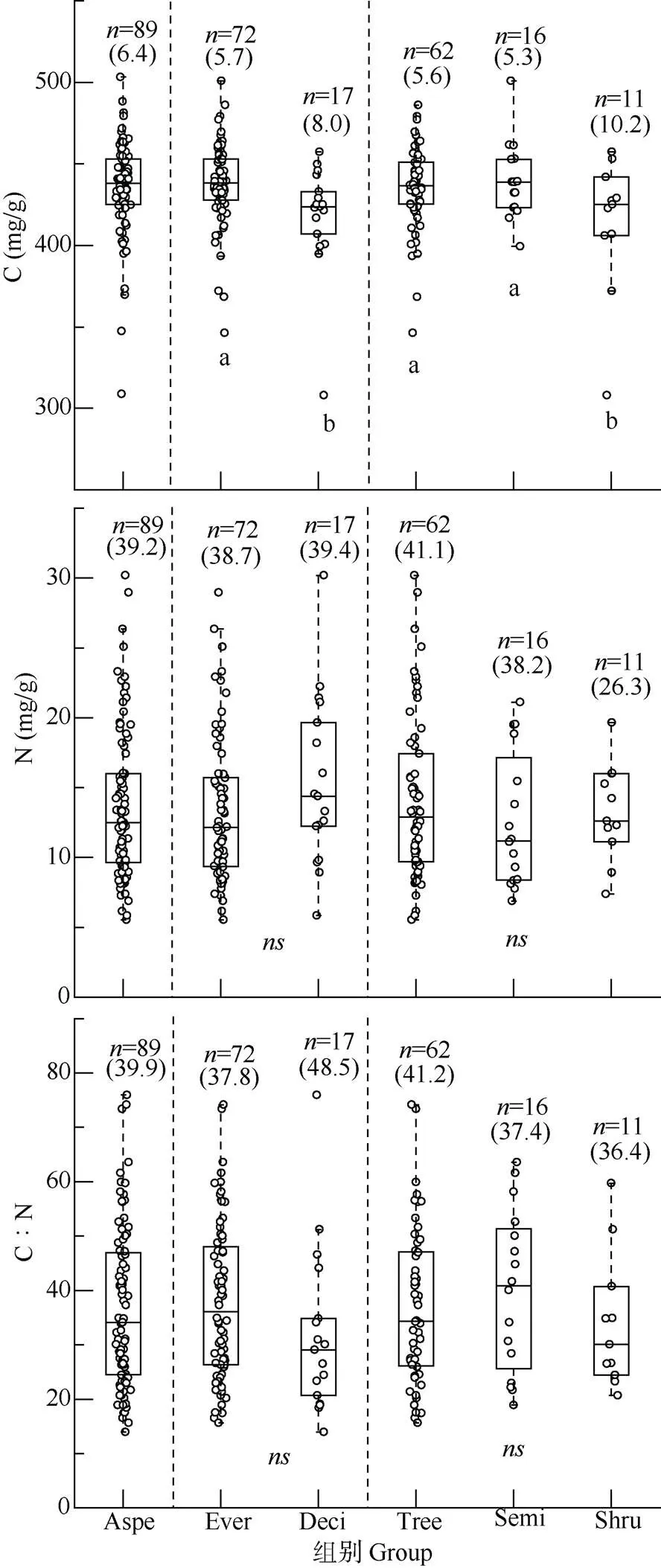
图1 木本植物一级根的C、N质量浓度和C∶N。Aspe: 所有树种; Ever: 常绿树种; Deci: 落叶树种; Tree: 乔木; Semi: 小乔木或灌木; Shru: 灌木; 括号内数值为变异系数(%); n: 样本数量; 不同字母表示差异显著(P<0.05); ns: 无差异。下图同。

图2 不同科植物一级根的C、N质量浓度和C∶N。Laur: 樟科; Faga: 壳斗科; Aqui: 冬青科; Symp: 山矾科; Pent: 五列木科; Elae: 杜英科。
2.2 一级根化学计量学的系统发育信号
89种木本植物一级根N浓度系统发育信号显著,Blomberg’s=0.142,=0.003,表明系统发育对亚热带天然常绿阔叶林一级根N浓度具有显著影响;而C浓度和C∶N的Blomberg’s分别为0.068和0.078,系统发育信号不显著(>0.05)。线性回归分析表明,一级根N浓度与分化时间呈显著正相关(2=0.08,<0.05, 图3),表明一级根N浓度受系统发育的影响较弱。

图3 科级一级根N浓度与分化时间的线性回归。MYA: 百万年前; 圆圈越大表示科所包含的物种数量越多。
3 结论和讨论
3.1 一级根C、N浓度和C∶N变异
本研究结果表明,89种木本植物一级根C浓度和C∶N的变异程度小于全球尺度的植物[18],但N浓度的变异程度则相反。万木林地区亚热带天然常绿阔叶林一级根平均C浓度低于全球平均水平[18],也低于绝大多数亚热带地区植物[10–11,13,16],但却高于摩天岭地区草本植物细根[12],这可能与样本数量有关,样本数量会直接影响整体结果的平均值。此外,植被类型不同也会造成根C浓度差异,如亚热带地区草本植物细根的C浓度远小于木本植物[12–13]。本研究中一级根C浓度的变异系数小于全球尺度, 变化范围处于全球水平(307.7~684.4 mg/g)[18]的前半段。
本研究中万木林地区亚热带常绿阔叶林群落的89种木本植物一级根平均N浓度低于全球植物[18],也低于其他地区植物[10–11,16],但却高于同属亚热带地区的神农架[13]和武夷山[23]植物。这很可能与植物根系取样方法相关,径级法和序级法会使植物根C、N浓度存在显著差异[11]。植物根系解剖结构,例如根维比值大小也会影响根组织C、N浓度[37]。本研究一级根N浓度的变异系数稍大于全球尺度,变化范围处于全球水平(5.6~50.3 mg/g)[18]的中前区间。
值得注意的是,本研究的89种木本植物一级根平均C∶N高于全球植物[18]和其他地区植物[11,16],却低于全国植物[38],这可能是一级根C、N浓度差异造成的,主要与样本量、植被类型[12–13]、根系取样方式[11]和根解剖结构[37]有关。本研究一级根C∶N变异系数小于全球尺度,其变化范围几乎与全球水平(2.1~77.8)[18]持平。
3.2 叶片习性、生长型和科级水平的差异
本研究叶片习性对一级根C浓度影响差异显著,可能与植物对环境的适应性有关,即表现出一定的趋同效应。不同叶片习性植物根化学计量学的差异可能与其生态策略相关,从叶片来看,常绿物种会通过更长的生长周期和更高的氮利用效率来减少养分流失(即减少落叶)[36],就根系而言, 则会增加木质素含量来提高根C含量[39],使植物代谢速率减慢,可见常绿树种在资源获取方面较为保守。本研究结果表明,常绿树种与落叶树种的一级根N浓度没有显著差异,与刘璐等[13]的研究结果一致,而周永姣等[20]报道落叶树种一级根N浓度显著高于常绿树种。这可能与样本数量及物种进化史有关。
本研究生长型对万木林地区89种木本植物一级根C浓度影响显著,对N浓度和C∶N没有影响,这可能与植物趋同效应有关。本文乔木、小乔木或灌木的一级根C浓度显著大于灌木,这与王钊颖等[23]研究结果相似,与张亚兴等[40]不同。与灌木相比,乔木树种高的林冠层能获取更多光能,其光合作用强度大于灌木,可能分配更多的C到植物根系。不同生长型植物生态位的分化可能造成其对地理环境(光、热、水等)适应性的差异,因而表现出不同的养分适应策略[41],使得不同生长型植物根C、N浓度存在一定差异。
本研究结果表明在土壤养分和水分条件相似的同一立地环境下,不同科的植物根系在资源分配方面存在差异。贾全全[42]研究结果表明,樟科一级根N浓度显著高于其他科植物(如壳斗科、杜鹃花科和山茶科),这与本研究结果一致。樟科植物一级根N浓度高可能与自身化学特性有关,其体内含有一种含氮的次生化合物生物碱。同时,不同植物的根C、N浓度差异最终会影响根的C∶N。
3.3 与系统发育的关系
植物功能性状的跨物种分析阐明了与生活史适应和资源经济策略相关的权衡[11]。本研究结果表明, 系统发育关系对亚热带地区木本植物一级根N浓度影响较小,化学性状中N更具有系统发育保守性。与本研究结果相似,Kong等[10]报道96种植物一级根化学性状受到系统发育关系的影响,N浓度系统发育信号显著。然而,Liu等[11]和Ma等[18]报道植物一级根化学性状几乎不受系统发育关系的影响。这可能与物种选择有关,热带地区天然林树种进化历史久远,存在着大量的古老树种,当把温带、亚热带地区新进化的树种囊括在一起时,可能会削弱系统发育关系的影响。一级根N浓度与科级水平分化时间呈显著正相关,表明进化历史古老的物种具有高的N浓度,这同植物一级根直径随物种科级水平分化时间增加而增大的结论一致[15,18,43], 从侧面反映出一级根N浓度可能与直径具有相关性。本研究中一级根C∶N没有表现出显著的系统发育信号,可能是受到了C的干扰。
本研究主要揭示了亚热带常绿阔叶林群落内具有种间可比性的一级根化学计量特征,及其受叶片习性、生长型和科级水平的影响。结果表明,亚热带天然常绿阔叶林一级根C浓度变异系数远低于N浓度和C∶N,反映出一级根C浓度的相对稳定性。叶片习性和生长型对一级根C浓度影响显著, 对N浓度和C∶N无影响, 可能与植物的趋同适应有关;主要科植物间一级根C、N浓度和C∶N均差异显著,说明不同科植物根系的资源利用与分配策略可能存在差异。亚热带天然常绿阔叶林一级根N浓度受系统发育影响较强,且与分化时间呈显著正相关,表明亚热带地区植物一级根N浓度具有强烈的系统发育保守性。
[1] Bloom A J, Chapin III A F, Mooney H A. Resource limitation in plants: An economic analogy [J]. Ann Rev Ecol Syst, 1985, 16: 363– 392. doi: 10.1146/annurev.es.16.110185.002051.
[2] ZHANG X Q, WU K H, MURACH D. A review of methods for fine-root production and turnover of trees [J]. Acta Ecol Sin, 2000, 20 (5): 875–883. doi: 10.3321/j.issn:1000-0933.2000.05.026.
张小全, 吴可红, MURACH D. 树木细根生产与周转研究方法评述 [J]. 生态学报, 2000, 20(5): 875–883. doi: 10.3321/j.issn:1000-0933. 2000.05.026.
[3] PREGITZER K S, DEFOREST J L, BURTON A J, et al. Fine root architecture of nine North American trees [J]. Ecol Monogr, 2002, 72(2): 293–309. doi: 10.1890/0012-9615(2002)072[0293:FRAONN]2.0.CO;2.
[4] MCCORMACK M L, DICKIE I A, EISSENSTAT D M, et al. Rede- fining fine roots improves understanding of below-ground contribu- tions to terrestrial biosphere processes [J]. New Phytol, 2015, 207(3): 505–518. doi: 10.1111/nph.13363.
[5] GILL R A, JACKSON R B. Global patterns of root turnover for terrestrial ecosystems [J]. New Phytol, 2000, 147(1): 13–31. doi: 10. 1046/J.1469-8137.2000.00681.X.
[6] NORBY R J, JACKSON R B. Root dynamics and global change: Seeking an ecosystem perspective [J]. New Phytol, 2000, 147(1): 3–12. doi: 10.1046/j.1469-8137.2000.00676.x.
[7] CHEN X P, GUO B Q, ZHONG Q L, et al. Response of fine root carbon, nitrogen, and phosphorus stoichiometry to soil nutrients inalong an elevation gradient in the Wuyi Mountains [J]. Acta Ecol Sin, 2018, 38(1): 273–281. doi: 10.5846/stxb201701040034.
陈晓萍, 郭炳桥, 钟全林, 等. 武夷山不同海拔黄山松细根碳、氮、磷化学计量特征对土壤养分的适应 [J]. 生态学报, 2018, 38(1): 273–281. doi: 10.5846/stxb201701040034.
[8] NADELHOFFER K J. The potential effects of nitrogen deposition on fine-root production in forest ecosystems [J]. New Phytol, 2000, 147(1): 131–139. doi: 10.1046/j.1469-8137.2000.00677.x.
[9] BASSIRIRAD H. Kinetics of nutrient uptake by roots: Responses to global change [J]. New Phytol, 2000, 147(1): 155–169. doi: 10.1046/j. 1469-8137.2000.00682.x.
[10] KONG D L, MA C E, ZHANG Q, et al. Leading dimensions in absorptive root trait variation across 96 subtropical forest species [J]. New Phytol, 2014, 203(3): 863–872. doi: 10.1111/nph.12842.
[11] LIU C, XIANG W H, ZOU L M, et al. Variation in the functional traits of fine roots is linked to phylogenetics in the common tree species of Chinese subtropical forests [J]. Plant Soil, 2019, 436(1): 347–364. doi: 10.1007/s11104-019-03934-0.
[12] ZHANG T, TIAN Q, LUO L J, et al. The biomass of herbaceous plant communities and its chemometrics characteristics of root C, N and P in the Motianling Northern Slope at low altitudes [J]. For Sci Technol, 2019(5): 3–6. doi: 10.13456/j.cnki.lykt.2018.07.10.0001.
张涛, 田青, 罗立娇, 等. 摩天岭北坡低海拔区草本植物生物量及根C、N、P化学计量学特征 [J]. 林业科技通讯, 2019(5): 3–6. doi: 10. 13456/j.cnki.lykt.2018.07.10.0001.
[13] LIU L, GE J L, SHU H W, et al. C, N and P stoichiometric ratios in mixed evergreen and deciduous broadleaved forests in Shennongjia, China [J]. Chin J Plant Ecol, 2019, 43(6): 482–489. doi: 10.17521/cjpe. 2019.0064.
刘璐, 葛结林, 舒化伟, 等. 神农架常绿落叶阔叶混交林碳氮磷化学计量比 [J]. 植物生态学报, 2019, 43(6): 482–489. doi: 10.17521/ cjpe.2019.0064.
[14] GENG Y, WANG L, JIN D M, et al. Alpine climate alters the relation- ships between leaf and root morphological traits but not chemical traits [J]. Oecologia, 2014, 175(2): 445–455. doi: 10.1007/s00442-014- 2919-5.
[15] CHEN W L, ZENG H, EISSENSTAT D M, et al. Variation of first- order root traits across climatic gradients and evolutionary trends in geological time [J]. Glob Ecol Biogeogr, 2013, 22(7): 846–856. doi: 10.1111/geb.12048.
[16] WANG R L, WANG Q F, ZHAO N, et al. Different phylogenetic and environmental controls of first-order root morphological and nutrient traits: Evidence of multidimensional root traits [J]. Funct Ecol, 2018, 32(1): 29–39. doi: 10.1111/1365-2435.12983.
[17] UAN Z Y, CHEN H Y H, REICH P B. Global-scale latitudinal patterns of plant fine-root nitrogen and phosphorus [J]. Nat Commun, 2011, 2: 344. doi: 10.1038/ncomms1346.
[18] MA Z Q, GUO D L, XU X L, et al. Evolutionary history resolves global organization of root functional traits [J]. Nature, 2018, 555 (7694): 94–97. doi: 10.1038/nature25783.
[19] GUO D L, XIA M X, WEI X, et al. Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species [J]. New Phytol, 2008, 180(3): 673–683. doi: 10.1111/j.1469-8137.2008.02573.x.
[20] ZHOU Y J, WANG M T, WANG Z Y, et al. Nutrient and ecological stoichiometry of different root order fine roots of 59 evergreen and deciduous tree species in subtropical zone [J]. Acta Ecol Sin, 2020, 40 (14): 4975–4984. doi: 10.5846/stxb201907031401.
周永姣, 王满堂, 王钊颖, 等. 亚热带59个常绿与落叶树种不同根序细根养分及化学计量特征 [J]. 生态学报, 2020, 40(14): 4975– 4984. doi: 10.5846/stxb201907031401.
[21] GONG S H. The influence of environment and phylogenic background on plant functional trait in Yanhe River Catchment [D]. Yangling: Northwest Agricultural and Forestry University, 2012: 35.
龚时慧. 环境与系统发育背景对延河流域植物群落功能性状的影响 [D]. 杨凌: 西北农林科技大学, 2012: 35.
[22] CHANG Y N, ZHONG Q L, CHENG D L, et al. Stoichiometric characteristics of C, N, P and their distribution pattern in plants ofnatural forest in Youxi [J]. J Plant Resour Environ, 2013, 22(3): 1–10. doi: 10.3969/j.issn.1674-7895.2013.03.01.
常云妮, 钟全林, 程栋梁, 等. 尤溪天然米槠林植物碳氮磷的化学计量特征及其分配格局 [J]. 植物资源与环境学报, 2013, 22(3): 1– 10. doi: 10.3969/j.issn.1674-7895.2013.03.01.
[23] WANG Z Y, CHENG L, WANG M T, et al. Fine root traits of woody plants in deciduous forest of the Wuyi Mountains [J]. Acta Ecol Sin, 2018, 38(22): 8088–8097. doi: 10.5846/stxb201712262331.
王钊颖, 程林, 王满堂, 等. 武夷山落叶林木本植物细根性状研究 [J]. 生态学报, 2018, 38(22): 8088–8097. doi: 10.5846/stxb201712262331.
[24] XU Y, BAO Y J, LI Z H, et al. Stoichiometry characteristics of carbon and nitrogen of grassland plants in the Agro-pastoral Ecotone of Inner Mongolia and Liaoning Border [J]. Chin J Gras, 2019, 41(4): 101–109. doi: 10.16742/j.zgcdxb.20180148.
徐媛, 鲍雅静, 李政海, 等. 蒙辽农牧交错区草地植物碳氮化学计量特征 [J]. 中国草地学报, 2019, 41(4): 101–109. doi: 10.16742/j. zgcdxb.20180148.
[25] LIU W X, ZHU K J. Characteristics of nitrogen and phosphorus stoi- chiometry of plants in different functional groups on Alpine Meadow in the eastern edge of Tibetan Plateau [J]. Chin J Gras, 2013, 35(2): 52–58. doi: 10.3969/j.issn.1673-5021.2013.02.0100.
刘雯霞, 朱柯嘉. 青藏高原东缘高寒草甸不同功能群植物氮磷化学计量特征研究 [J]. 中国草地学报, 2013, 35(2): 52–58. doi: 10.3969/ j.issn.1673-5021.2013.02.0100.
[26] YU H L, FAN J W, ZHONG H P, et al. Characteristics of N and P stoichiometry of plants in different functional groups in the Qinghai- Tibet Plateau regions [J]. Acta Ecol Sin, 2017, 37(11): 3755–3764. doi: 10.5846/stxb201604040609.
于海玲, 樊江文, 钟华平, 等. 青藏高原区域不同功能群植物氮磷生态化学计量学特征 [J]. 生态学报, 2017, 37(11): 3755–3764. doi: 10.5846/stxb201604040609.
[27] ZHUO M X. Stoichiometric characteristics of eight Lauraceae species in a subtropical evergreen broad-leaved forest [J]. J Subtrop Resour Environ, 2019, 14(1): 17–22. doi: 10.19687/j.cnki.1673-7105.2019.01.003.
卓鸣秀. 亚热带常绿阔叶林8种樟科树种细根化学计量特征 [J]. 亚热带资源与环境学报, 2019, 14(1): 17–22. doi: 10.19687/j.cnki.1673- 7105.2019.01.003.
[28] ZHOU M, BAI W M, ZHANG Y S, et al. Multi-dimensional patterns of variation in root traits among coexisting herbaceous species in temperate steppes [J]. J Ecol, 2018, 106(6): 2320–2331. doi: 10.1111/ 365-2745.12977.
[29] VALVERDE-BARRANTES O J, SMEMO K A, BLACKWOOD C B. Fine root morphology is phylogenetically structured, but nitrogen is related to the plant economics spectrum in temperate trees [J]. Funct Ecol, 2015, 29(6): 796–807. doi: 10.1111/1365-2435.12384.
[30] VALVERDE-BARRANTES O J, FRESCHET G T, ROUMET C, et al. A worldview of root traits: The influence of ancestry, growth form, climate and mycorrhizal association on the functional trait variation of fine-root tissues in seed plants [J]. New Phytol, 2017, 215(4): 1562– 1573. doi: 10.1111/nph.14571.
[31] GUO D L, MITCHELL R J, HENDRICKS J J. Fine root branch orders respond differentially to carbon source-sink manipulations in a longleaf pine forest [J]. Oecologia, 2004, 140(3): 450–457. doi: 10.1007/s00442- 004-1596-1.
[32] ZANNE A E, TANK D C, CORNWELL W K, et al. Three keys to the radiation of angiosperms into freezing environments [J]. Nature, 2014, 506(7486): 89–92. doi: 10.1038/nature12872.
[33] WANG X, CHEN G S, YAN X J, et al. Variations in the first-order root diameter in 89 woody species in a subtropical evergreen broadleaved forest [J]. Chin J Plant Ecol, 2019, 43(11): 969–978. doi: 10.17521/cjpe. 2019.0189.
王雪, 陈光水, 闫晓俊, 等. 亚热带常绿阔叶林89种木本植物一级根直径的变异 [J]. 植物生态学报, 2019, 43(11): 969–978. doi: 10. 17521/cjpe.2019.0189.
[34] BLOMBERG S P, GARLAND JR T, IVES A R. Testing for phylo- genetic signal in comparative data: Behavioral traits are more labile [J]. Evolution, 2003, 57(4): 717–745. doi: 10.1111/j.0014-3820.2003.tb00285.x.
[35] WIKSTRÖM N, SAVOLAINEN V, CHASE M W. Evolution of the angiosperms: Calibrating the family tree [J]. Proc Roy Soc B Biol Sci, 2001, 268(1482): 2211–2220. doi: 10.1098/rspb.2001.1782.
[36] ZHANG J H, HE N P, LIU C C, et al. Variation and evolution of C∶N ratio among different organs enable plants to adapt to N-limited environ- ments [J]. Glob Chang Biol, 2020, 26(4): 2534–2543. doi: 10.1111/gcb. 14973.
[37] XU Y, GU J C, DONG X Y, et al. Fine root morphology, anatomy and tissue nitrogen and carbon contents of the first five orders in four tropical hardwood species in Hainan Island, China [J]. Chin J Plant Ecol, 2011, 35(9): 955–964. doi: 10.3724/SP.J.1258.2011.00955.
许旸, 谷加存, 董雪云, 等. 海南岛4个热带阔叶树种前5级细根的形态、解剖结构和组织碳氮含量 [J]. 植物生态学报, 2011, 35(9): 955–964. doi: 10.3724/SP.J.1258.2011.00955.
[38] MA Y Z, ZHONG Q L, JIN B J, et al. Spatial changes and influencing factors of fine root carbon, nitrogen and phosphorus stoichiometry of plants in China [J]. Chin J Plant Ecol, 2015, 39(2): 159–166. doi: 10. 17521/cjpe.2015.0015.
马玉珠, 钟全林, 靳冰洁, 等. 中国植物细根碳、氮、磷化学计量学的空间变化及其影响因子 [J]. 植物生态学报, 2015, 39(2): 159–166. doi: 10.17521/cjpe.2015.0015.
[39] NOVAES E, KIRST M, CHIANG V, et al. Lignin and biomass: A negative correlation for wood formation and lignin content in trees [J]. Plant Physiol, 2010, 154(2): 555–561. doi: 10.1104/pp.110.161281.
[40] ZHANG Y X, ZHU L W, LIU N. C, N, and P concentrations and their stoichiometry of leaves and roots with different life forms in Hainan Province [J]. J Trop Subtrop Bot, 2020, 28(2): 131–135. doi: 10.11926/ jtsb.4115.
张亚兴, 朱丽薇, 刘楠. 海南不同生活型植物叶片和根系C、N、P化学计量特征 [J]. 热带亚热带植物学报, 2020, 28(2): 131–135. doi: 10.11926/jtsb.4115.
[41] WANG X J. The geographical differences of plant functional traits and functional diversity in broad-leaved Korean pine forests in the north- east of China [D]. Beijing: Beijing Forestry University, 2015: 42.
王晓洁. 东北阔叶红松林植物功能性状与功能多样性的地理差异研究 [D]. 北京: 北京林业大学, 2015: 42.
[42] JIA Q Q. Functional traits of fine roots and their relationship with leaf traits of 50 major species in a subtropical forest in Gutianshan [D]. Qiqihar: Qiqihar University, 2011: 58.
贾全全. 古田山亚热带森林50个主要树种细根功能属性及其与叶片相关性研究 [D]. 齐齐哈尔: 齐齐哈尔大学, 2011: 58.
[43] ST JOHN T V. Root size, root hairs and mycorrhizal infection: A re- examination of Baylis’s hypothesis with tropical trees [J]. New Phytol, 1980, 84(3): 483–487. doi: 10.1111/j.1469-8137.1980. tb04555.x.
Variation Patterns in C and N Concentrations in the First-order Roots of 89 Woody Species in Subtropical Evergreen Broad-leaved Forest
WANG Xue1, YAN Xiaojun1,2, FAN Ailian1, JIA Linqiao1, XIONG Decheng1, HUANG Jinxue1, CHEN Guangshui1, YAO Xiaodong1 *
(1. School of Geographical Sciences, Fujian Normal University, Key Laboratory for Humid Subtropical Eco-geographical Processes of the Ministry of Education,Fuzhou 350007, China; 2. Mudu High School in Jiangsu Province,Suzhou 215101, Jiangsu, China)
In order to understand the variation patterns of nutrient elements for the first-order roots of woody plants in subtropical evergreen broad-leaved forests,the carbon and nitrogen concentrations of the first-order roots of 89 tree species in Wanmu Forest Nature Reserve, Jian’ou, Fujian Province, were determined based on root order method. The results showed that the mean C, N concentrations in the first-order roots of 89 tree species were 433.9 and 13.7 mg/g, and C∶N ratio was 36.7, which variation coefficients were 6.4%, 39.2% and 39.9%, respectively. There were significant differences in C concentration of the first-order roots among different leaf habits (such as evergreen and deciduous trees) and growth forms (including tree, semi-tree or shrub and shrub). However, there was no significant difference in N concentration and C∶N ratio. The differences in C, N concentrations and C∶N ratio of the first-order roots among six main families (Lauraceae, Fagaceae, Aquifoliaceae, Symplocaceae, Pentaphylacaceae and Elaeocarpaceae) were significant. The N concentration of first-order roots increased with the phylogeny level from low to high. Therefore, it was indicated that the interspecific variation of the first-order roots in C concentration was lower than N concentration in the subtropical evergreen broad-leaved forest, the N concentration of the first-order roots was influenced by phylogeny, however, the concentration of C was affected by leaf habits and growth forms, showing a certain convergence effect.
Absorptive root; Stoichiometry; Variation characteristics; Nature forest
10.11926/jtsb.4372
2020-12-29
2021-02-28
国家自然科学基金项目(31830014)资助
This work was supported by the National Natural Science Foundation of China (Grant No. 31830014).
王雪,女,硕士研究生,从事植物生理生态研究。E-mail: 937621220@qq.com
. E-mail: xdyao@fjnu.edu.cn
