格氏栲自然保护区常绿阔叶林群落优势树种种间联结性分析
2021-10-13宋育红邢建宏邓贤兰
*宋育红, 邢建宏, 邓贤兰
(1.三明学院应用生物技术研究所,福建,三明 365004; 2.井冈山大学生命科学学院,江西,吉安 343009)
格氏栲(Castanopsis kawakamii)是我国中亚热带南缘特有的壳斗科(Fagaceae)常绿大乔木,属国家二级保护植物。其自然分布范围较窄,多数零星生长在海拔200~1000 m 的丘陵地带的常绿阔叶林中,但唯独在福建省三明小湖地区格氏栲自然保护区内有近700 hm2的格氏栲天然林,是我国面积最大的格氏栲天然林,多由百年以上的大树组成,属亚热带常绿阔叶林中罕见的群落类型[1-2],具极高的物种价值和生态价值。为了保护这片独特罕见的格氏栲天然林,有不少学者对其进行了研究,如阳含熙、樊后保、刘金福与洪伟等分别对格氏栲群落的数量分类、群落结构特征和种群生态学进行了研究,研究结果为格氏栲种群的保护提供了有价值的参考[3-8]。然而在保护区常绿阔叶林中,除了优势种群格氏栲林外,尚有其他阔叶树种群[2],如米槠(Castanopsis carlesii)林、甜槠(Castanopsis eyrei)林、木荷(Schima superba)林和闽楠(Phoebe bournei)林等[9],其他阔叶树种群存在和扩张有可能对格氏栲林的更新演替产生影响。目前对该保护区常绿阔叶林优势种群种间联结关系的研究尚未见报道。
种间联结是指不同物种在空间分布的相互关联性,同时也隐含着物种之间功能依赖关系[10],种群间关联性直接影响着群落物种组成和结构、更新和演替方向[11-13]。通过对种间关联的研究,能客观反映物种在空间上的相互关系,全面了解群落种类组成间的相互依存和相互制约的关系,对揭示群落动态和演替等机制具有重要意义[14-16]。种间联结性研究对珍稀濒危植物的保护具有一定的参考价值[17-20]。本研究采用种间联结的方差比率法VR、x2检验、联结系数AC 值、Jaccard 指数及点相关系数等系列关联测度值,对格氏栲自然保护区常绿阔叶林群落中优势乔木种群的种间联结进行了分析,研究结果有助于了解该保护区乔木层主要优势种间的联结关系,揭示格氏栲这一珍稀濒危植物与其他树种间的关系,为该保护区常绿阔叶林物种多样性保护,尤其是格氏栲种群保护提供资料和依据。
1 研究区自然环境和植被概况
格氏栲自然保护区位于福建三明市西南方,总面 积 1125.6 hm2。 地 处 26°07′~26°10′N ,117°24′~117°27′E,位于中亚热带南缘,武夷山东伸支脉地带,属低山丘陵。海拔一般在250~500 m 之间,最高峰海拔604 m[2]。年平均气温19.5°C,年无霜期300 d 左右,年平均降水量1700 mm;地带性土壤为红壤,多数达100~150 cm,土层腐殖质丰富,土壤从海拔低到高依次为:暗红壤、山地红壤、黄红壤[1-2]。地带性植被为常绿阔叶林,群落垂直层次分化比较明显,分为乔木层、灌木层和草本层,乔木层高约25 m,可分为3 个亚层。第一亚层高20 m 以上,以格氏栲、木荷、马尾松(Pinus massoniana)等占优势;第二亚层高约10~20 m,植物种类比较丰富,主要由米槠、甜槠、多毛茜草树(Aidia pycnantha)、中华杜英(Elaeocarpus chinensis)等组成;第三亚层高度约为5~10 m,主要由木荚红豆(Ormosia xylocarpa)、华南木姜子(Litsea greenmaniana)、华幌伞枫(Heteropanax chinensis)等小乔木和部分高大乔灌木如连蕊茶(Camellia fraterna) 、 乌 饭 树(Vaccinium bracteatum)、鼠刺(Itea chinensis)等组成,群落乔木层的郁闭度在80%左右。灌木层盖度为30%~40%,主要由连蕊茶、乌饭树、鼠刺、绒楠(Machius velatina)、杜茎山(Maesa japonica)、狗骨柴、沿海紫金牛(Ardisia punctata)等组成。草本层植物种类少,以山姜(Alpinia japonica)、黑莎草(Gahnia tristis)、芒萁(Picranopteris dichotoma)、狗脊蕨(Woodwardia japonica)占优势。层间植物主要由藤本植物组成,以网脉叶酸果藤(Embelia rudis)、香花崖豆藤(Millettia dielsiana)、省藤(Calamus platyacanthoides)等占优势。
2 研究方法
2.1 野外调查
在对保护区常绿阔叶林群落作全面踏勘的基础上,根据海拔高度、环境条件的不同设置16 个样地,每个样地设置4 个10 m×12.5 m 的样方,共计64 个样方;在每个样方内调查记录乔木层树种(树高>5 m)的植物名称、胸围、树高、冠幅,同时在每一样方中设置1 个5 m×5 m 灌木样方调查记录灌木层中乔木幼树、幼苗的种名和数量以了解乔木种群自然更新情况,并测定各样地的海拔、坡度、坡向、土壤状况、郁闭度等环境因子。
2.2 数据处理
依据野外调查资料,选取群落样方乔木层中出现频率大于10%的25 个树种进行分析(表1)。采用2×2 联列表的形式排列并计算种对间的a(种A与种B 同时出现的样方数)、b(只有种B 出现的样方数)、c(只有种A 出现的样方数)、d(种A 与种B 都不出现的样方数)值,作为种间联结测定的基础数据。树木重要值=(相对频度+相对密度+相对显著度)/3 。
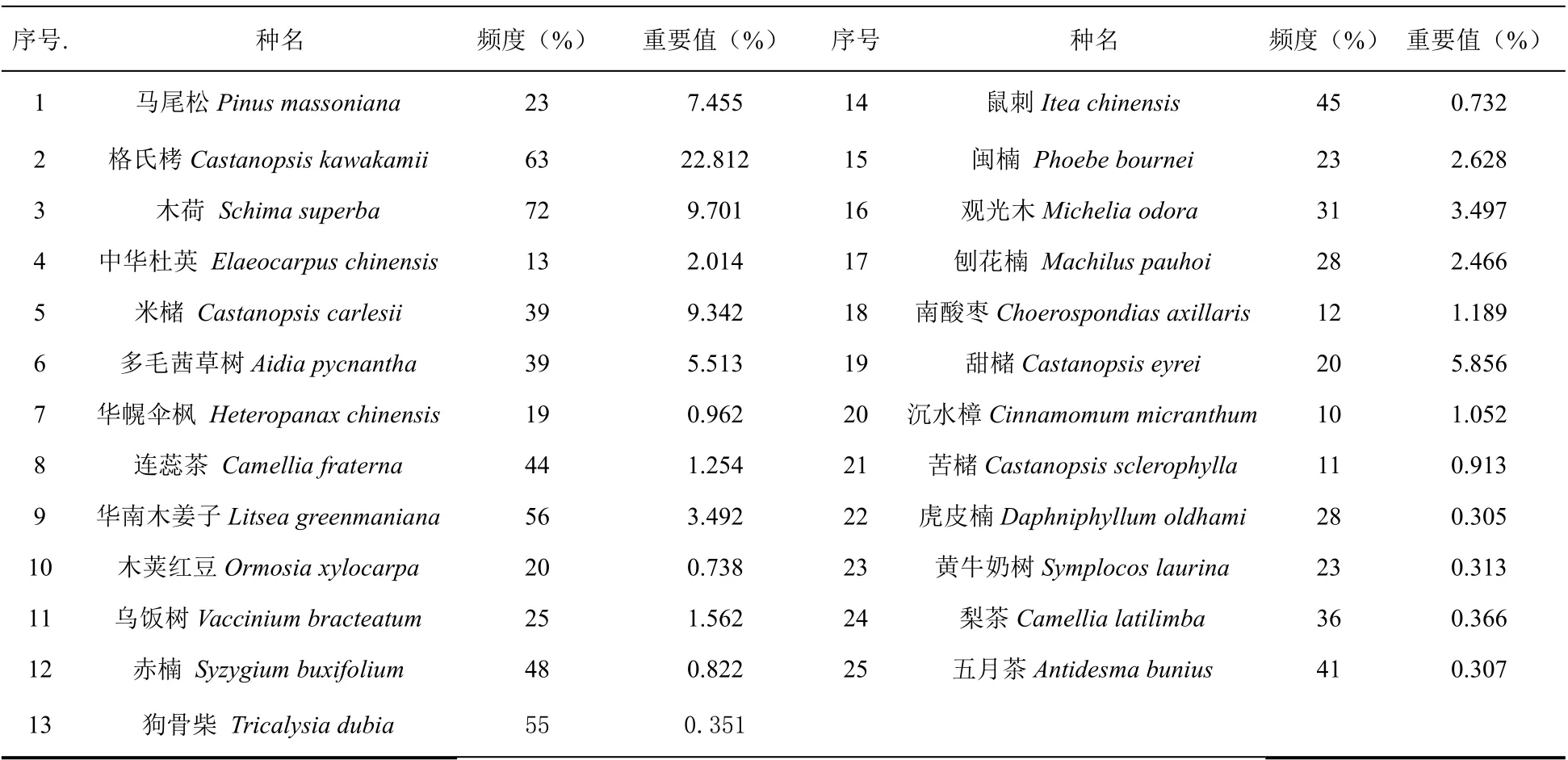
表1 25 个乔木层优势种的情况Table 1 The information of 25 dominant tree species
2.3 总体关联性测定
采用D.Schluter 提出的方差比率法(VR)检验整个群落物种间的总体关联性。计算方法和公式参见文献[21,22]。
2.4 种对间联结性测定
将各样方调查资料统计数据排列成2×2 联列表,计算4 项关联测值:联结系数AC、Jaccard 指数(共同出现百分率)、点相关系数Φ、x2检验,计算方法和公式见文献[21-23],并以各指标计算值作半矩阵图。联结系数AC 和点相关系数Φ用于说明种间联结的程度,Jaccard 指数用来测度物种间相伴出现机率和正联结程度,x2检验用来测度种间联结性的显著程度。x2本身没有负值,判断种间正负联结的方法是:(ad-bc)>0 为正联结;反之,为负联结[23]。
3 结果与分析
3.1 总体关联性分析
根据25 个优势种群之间存在与不存在矩阵,计算VR值:VR=ST2/δT2=1.541>1, 说明25 个种群间表现出净正关联。再根据统计量W来检测VR值偏离1 的显著性:W=VR×N=98.62,查表(文献[25]),得相应x2值:x20.95(64)= 46.595,x20.05(64)=83.675,W落在x20.95(64)和x20.05(64)之外,说明VR偏离1 显著,显示25 个种群间的整体联结性为显著的正联结。种间总体关联性反映了群落稳定性[26],随着植被群落演替的进展,群落结构及其种类组成将逐渐趋于完善和稳定,种间关系也将逐步趋于正相关,以求得物种间的稳定共存[27-28]。格氏栲自然保护区常绿阔叶林群落种间整体联结性表现为显著的正联结,表明群落该群落已处于和该地区环境条件相适应的稳定的顶级阶段。
3.2 种间关联测度值分析
根据原始资料矩阵,运用上述公式计算出25个优势种群(表1)种间关联测度各指标值,分别作半矩阵图,包括联结系数AC 值(图1)、Jaccard指数JI 值(图2)、点相关系数Φ值(图3)及x2值(图4)。
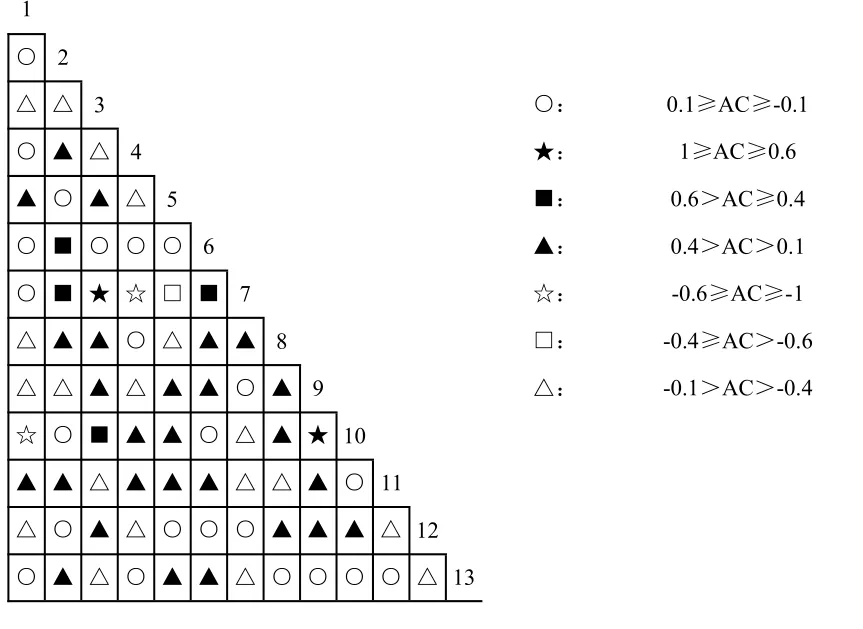
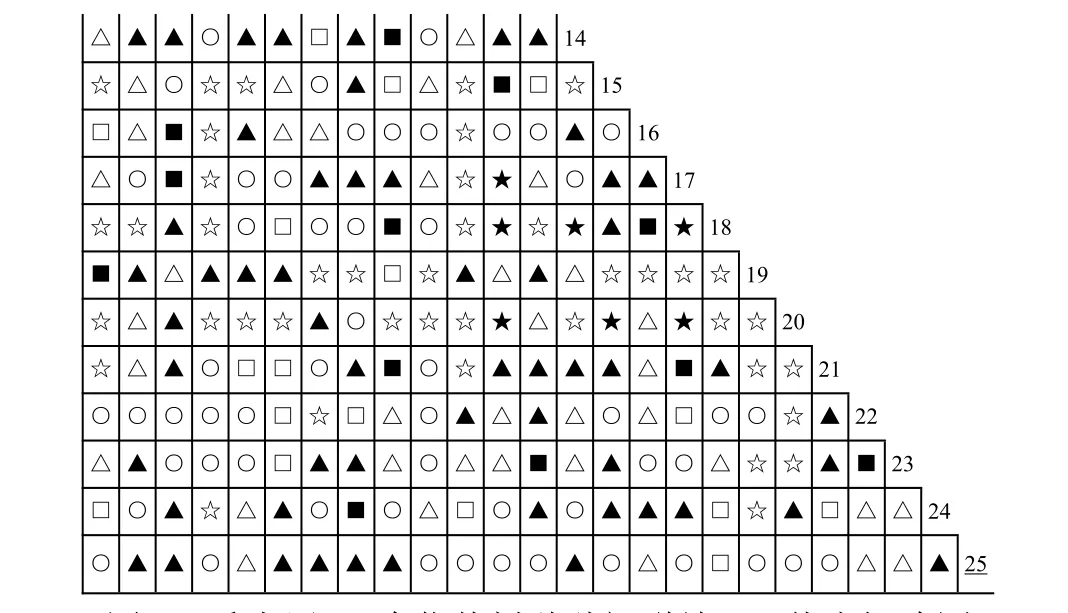
图1 乔木层25 个优势树种种间联结AC 值半矩阵图(种序号同表1,下同)Fig.1 Semi-matrix diagram of AC correlation coefficients for 25 dominant species in the tree layer

图2 乔木层25 个优势树种种间联结JI 值半矩阵图Fig.2 Semi-matrix diagram of JI correlation coefficients for 25 dominant tree species

图3 乔木层25 个优势树种种间联结系数Φ值半矩阵图Fig.3 Semi-matrix diagram of Φ correlation coefficients for 25 dominant tree species

图4 乔木层25 个优势树种x2 测定半矩阵图Fig.4 Semi-matrix diagram of x2 correlation test for 25 dominant tree species
在25 个优势种群的300 个种对组合中,AC 值在-0.4~0.4 之间的弱联结种对有214 对,占总对数的71.3%;JI<0.4 的弱联结种对有244 对,占81.33%;Φ值处于-0.2~0.2 值域的弱联结种对达211对,占70.3%;x2检测有264 个种对,占88%的种对种间联结(x2<3.841)未达到显著程度。以上4项种间关联测度值均显示群落中大多数种对的联结性较弱,种间独立性相对较强。
AC≥0.6 的极显著正联结种对9 对,0.6>AC≥0.4 的显著正联结种对有16 对;AC≤-0.6 极显著负联结的种对43 对,其中AC 值为-1 的完全负联结性种对32 对,这是因为如果两物种没有共同出现的样地(即a=0),AC 则为-1,造成得到极显著负相关的物种对较多,与实际情况不符,因此需结合x2检验的结果来进行分析。Φ值在0.4 以上极显著正联结种对有9 对,φ值在-0.4 以下的极显著负联结种对仅1 对。x2检测极显著正联结x2≥6.635的种对有13 对,显著的正联结(6.635>x2≥3.841)种对10 对;极显著负联结2 对,显著负联结11 对。以上种间关联测度各指标值显示群落显著与极显著联结的正联结种明显对多于负联结种对。
3.3 优势树种种间的联结关系分析
种间关联测度AC值、Jaccard指数、Φ值及x2检测4 项指标计算结果显示:群落优势种绝大多数种对的种间联结关系松散,未达到显著水平,种间独立性相对较强。这是由于在长期的演替过程中,由于种内、种间的竞争,使群落的组成成分趋于稳定,在进一步分化了的生态位中,各个物种都占据有利于自己或适合自己的位置和谐共处,相互依赖和相互竞争大为降低和减弱,所以多数种对联结程度不强。Φ值和x2检测显示25 个优势种群的300 个种对组合中,显著正联结种对多于显著负联结种对,说明表明格氏栲自然保护区常绿阔叶林群落目前属比较稳定、成熟的顶级群落。
植物种对的正联结体现了植物利用资源的相似性和生态位的重叠性[29],其植株高度的差别,体现了生态位空间分异,这是森林群落自然演替的必然结果[22]。综合分析4 项指标值,群落极显著正联结的种对有10 对,如1-19(马尾松-甜槠)、2-6(格氏栲-多毛茜草树)、15-20(闽楠-沉水樟)、17-18(刨花楠-南酸枣)等。形成显著正联结的原因之一在于树种生态学特性及其对环境要求的相似性,例如:1-19(马尾松-甜槠),它们都为阳性树种,常分布在山的中上坡和山脊等阳光充足的地方;15-20(闽楠-沉水樟)则较耐荫喜湿,常分布在海拔较低的山体中下坡湿润的环境;8-24(连蕊茶-梨茶)、9-10(华南木姜子-木荚红豆)、12-17(赤楠-刨花楠)等,由于它们对生境有相同的适应性而呈极显著正联结。正联结的另一原因是生态位空间分异,一个种的生长为另一个种的生存创造了条件,即它们对生境要求的互补有关[30],如2-6(格氏栲-多毛茜草树),格氏栲是群落的建群种,位于乔木层的第一层,为稍耐荫处于乔木层第二层的多毛茜草树提供了有利的环境条件,环境要求的同质性和生态位的空间分离使物种之间相互兼容,导致它们相伴出现的概率大大提高,多毛茜草树相当于是格氏栲的伴生种;类似的种对还有南酸枣-刨花楠、米槠与赤楠、甜槠与乌饭树、闽楠与梨茶等,二者在高度上的空间分异,生态位的差异使它们避免了激烈的竞争而得以在同一个群落中共存[30]。
负联结种对中,AC 值为-1,JI 值为0 的完全负联结性种对有32 对,表明这些种对之间无关联,完全不能出现在同一生境,如18-19(南酸枣-甜槠)、20-21(沉水樟-苦槠)、4-15(中华杜英-闽楠)等。x2值显示极显著负联结2 对,分别是8-19(连蕊茶-甜槠)、19-24(甜槠-梨茶);显著负联结11对,如1-15(马尾松-闽楠)、2-16(格氏栲-观光木)、3-19(木荷-甜槠)、5-15(米槠-闽楠)等。植物种对的负联结是由于空间资源的有限性导致种间竞争排斥,或是由于长期适应不同的微环境,利用不同的资源空间形成生态位分离而表现出负联结。如甜槠与梨茶、连蕊茶表现极显著负联结的原因,是由于甜槠对光照要求较强,而梨茶、连蕊茶喜湿耐荫,由于对生境要求的不同,因此表现出极显著的负联结,类似的有马尾松与闽楠、木荷与甜槠、米槠与闽楠等。有些种对由于生态位相同或相似,对环境资源有共同的要求(如光照、水分、湿度等)而产生强烈的竞争,表现出物种之间的排斥性,如格氏栲与观光木,都位于乔木层相同层次,两者对光照等环境条件要求相似,二者之间产生了强烈的竞争和物种之间的排斥性,从而形成显著负联结。
4 讨论
种间相关性与联结性的精确性在很大程度上取决于取样面积,取样面积过大,理论上出现正联结植物种对的机率将随之增加,而负联结的机率相对降低;而取样面积过小,则又可能形成负联结过多的情况[8]。对中亚热带常绿阔叶林种间联结研究,一般取样面积为100 m2[21-24]。本研究区域位于中亚热带南缘,纬度偏低,因此取样面积定为125 m2。计算结果显示,正联结种对数占总数50.77%,负联结占49.23%,正负联结种对数相差不大,所以该取样面积是适合的,与王伯荪[14]的研究结果一致。
本研究用方差比率法来判断种间的总体联结性,采用AC、JI、Φ值共同测定物种之间的关联度,并通过x2值检验其联结显著性,各指标值的综合分析能较准确全面地反映出种间联结性。格氏栲自然保护区常绿阔叶林群落25 个优势种群间的总体相关性检验VR值为1.541>1,显示整体联结性为显著的正联结;绝大多数种对的AC、JI、Φ值均不高,为不显著联结关系。这是由于在长期的演替过程中,由于种内、种间的竞争,各个物种都占据有利于自己或适合自己的位置和谐共处,相互依赖和相互竞争大为降低和减弱,所以多数种对联结关系松散,独立性较强。x2检测具极显著正联结种对有13对,显著正联结种对10 对,极显著负联结2 对,显著负联结11 对,显著和极显著正联结种对数多于显著和极显著负联结种对数。以上研究数据显示,目前格氏栲自然保护区常绿阔叶林群落已处于和该地区环境条件相适应的稳定顶极阶段,群落内种群间的联结关系符合目前的群落特征。
群落中占绝对优势地位的格氏栲除了与多毛茜草树呈极显著正联结,与观光木呈显著负联结外,与其余树种的联结关系较弱,为不显著联结,这与格氏栲种群多由百年以上的老龄个体组成而形成随机分布空间格局或低聚集度分布有关,因而表现出与多数树种有一定相互独立倾向[8]。格氏栲与其它24 个优势种的联结关系中,不显著联结种对占91.7%,说明调查区绝大部分优势树种易与格氏栲混生,形成较为稳定的群落。
群落乔木层重要值列前5 位(表1)的树种为格氏栲、木荷、米槠、马尾松和甜槠,格氏栲在群落中重要值最高,占据较宽的生态位,在群落中占绝对优势地位。顶极群落在正常情况下虽然不会发生质的变化,但稳定是相对的,量的变化仍贯穿整个生命过程,种内竞争和种间竞争时刻存在着[21]。目前格氏栲在群落中处于绝对优势地位,位于乔木第一亚层,平均树高26 m,平均胸径45 cm。绝大多数格氏栲树龄达百年以上,个别老树200 年以上,有些区域格氏栲存在过熟现象,树干腐心严重,树冠已开始变稀。乔木第二、第三亚层格氏栲幼树数量很少。格氏栲种群自然更新状况值得探究,一般林冠上层树木竞争较弱,竞争主要发生在中下层物种间[18]。野外调查数据统计显示,灌木层中格氏栲、木荷、米槠和甜槠苗在样方中出现的频度分别为26%、47.7%、30.7%和14.8%,灌木层中木荷、米槠幼苗出现的频度和数量均明显多于格氏栲苗,甜槠苗也达到一定的数量规模,这些阔叶树幼苗的耐荫性和生态适应性更强,与格氏栲苗形成了强的竞争关系,威胁到格氏栲种群的天然更新。目前该保护区常绿阔叶林群落除了有大面积的格氏栲林外,还有成片的米槠林、甜槠林、木荷林等,随着时间的推移其它阔叶树种群的扩张导致的种间竞争加剧有可能影响到格氏栲种群现有的规模,格氏栲种群未来发展趋势有待进一步研究。
