滨海防护林木麻黄和花吊丝竹混交林群落的种间关联和生态位分析
2021-10-13郑俊鸣李敏张盟TarinMuhammadWaqqasKhan何天友陈凌艳陈礼光洪小龙郑郁善
郑俊鸣, 李敏, 张盟, Tarin Muhammad Waqqas Khan, 何天友,陈凌艳, 陈礼光, 洪小龙, 郑郁善*
滨海防护林木麻黄和花吊丝竹混交林群落的种间关联和生态位分析
郑俊鸣1a, 李敏1a, 张盟1b, Tarin Muhammad Waqqas Khan1a, 何天友1a,陈凌艳1a, 陈礼光1b, 洪小龙2, 郑郁善1a*
(1. 福建农林大学, a. 园林学院; b. 林学院,福州 350002; 2. 福建省东山赤山国有防护林场,福建 漳州 363400)
为了解东山岛滨海防护林木麻黄()和花吊丝竹(var.)混交林群落的稳定性,采用样方法调查分析群落的种间关联和生态位。结果表明,群落总体的种间联结呈现不显著的正关联(=1.72,=20.625),表明群落处于不稳定的动态演替状态。花吊丝竹和马缨丹()、梵天花()和木麻黄表现出非常紧密的关联性和共现性(PC≥0.7, OI≥0.8, DI≥0.8)。马缨丹等外来植物生态位宽度较高,与花吊丝竹具有较高的生态位重叠,从而与花吊丝竹具有较高的正联结,使得群落处于不稳定状态。滨海防护林生境单一,优势物种习性相似,使花吊丝竹与多数物种形成强的正关联。受到环境资源的限制,强的正关联与物种生态位宽度较大有关。种对间强的正关联以及外来物种潜在的生存优势可能是影响群落稳定性的重要因素。
木麻黄;花吊丝竹;群落;种间联结;生态位;滨海沙地
种间联结是指不同物种在群落中的相互关系,反映了物种在群落中的地位。研究种间联结可揭示物种的共存机制,为群落演替发展、稳定和构建提供参考[1]。种间联结的研究对象通常为天然植被, 如海岛丘陵林地[2]、草原[3]、山地[4]和热带雨林[5]。对人工植被的群落种间联结研究相对较少,部分学者认为人工林的空间分布格局和种间联结主要是人为导向,研究意义不大。随着林分发展过程中林分更新和生态竞争,群落的分布格局逐渐形成较弱的聚集分布,与自然群落分布格局较为相似[6]。现阶段,人们逐渐将研究重点集中在人工防护林上,如通过研究人工林的种间关联以揭示人工植被自然恢复的演替规律[7];为低效人工林的森林可持续经营和养护提供参考[8];发挥公益林最大的综合性生态效益[9]。因此,研究人工林群落种间联结, 对植被的恢复与重建,防止生态退化、森林经营管理等问题有着重要的意义。
福建省海岸带以沙质海岸为主,少部分为基岩和泥质海岸[10]。滨海沙地防护林的主要优势种为木麻黄()、桉树(spp.)和湿地松()等。人工防护林物种多样性低,群落结构单一,林分更新困难,森林生产力弱等问题突出[11],但滨海人工防护林群落的种间联结、生态位和稳定性的研究相对较少。引入物种能丰富滨海防护林的生物多样性,增加群落稳定性[11]。其中,竹类植物具有较强的应用潜力,生长繁殖能力强,竹材纤维长,具有较强的耐旱和抗风能力。20多年前竹类植物被引种到福建东山滨海沙地防护林。有研究表明,混交竹林对群落多样性和均匀度高于木麻黄纯林[12]。同属植物的种间关联能增强竞争力,排斥非亲缘关系的物种以获得更多的资源空间[13],但混交竹林的种间联结和生态位尚未有报道。关注一些特殊物种和极小种群的种间联结和生态位分析有助于理解该物种对生境的特殊要求,为物种保护提供科学依据[14]。花吊丝竹(var.)原产于广西南部的低海拔山谷,引种后的花吊丝竹对沙地的土壤质量具有改良作用[15]。花吊丝竹应对沙地的干旱胁迫在生理和蛋白层面产生相应的适应策略[16],具有较强的适应能力和研究价值。本文旨在研究滨海沙地防护林的木麻黄和花吊丝竹混交林群落种间联结和生态位,探讨群落的稳定性,为今后的滨海防护林建设和物种筛选提供参考。
1 材料和方法
1.1 研究区概况
研究区位于福建省漳州市东山县东山岛,位于23°40ʹ N,118°18ʹ E,属于亚热带海洋性季风气候,干湿季节明显,湿季集中在5-9月,旱季集中在11月-翌年2月。东山岛10月-翌年3月有98 d为大风天,占全年大风天数的80%;风速6~8 m/s,年均风速7.1 m/s;年降水量为1 113.9 mm,蒸发量为2 013.2 mm; 年均温20.8℃,最高温36.6℃,最低温3.8℃;平均径流量为481 mm[11]。台风和干旱是当地主要的自然灾害,台风多发于7-8月。
1.2 样方选取和野外调查
2018年8-10月,在福建省东山岛滨海沙地选取木麻黄和花吊丝竹混交林群落分布的区域,设置16个20 m×20 m的样方。调查记录样方内所有物种的名称、高度和数量等信息。剔除偶见种和草本植物后,计算木本物种在群落中的相对重要性(表1)。参考江焕等[17]的方法计算重要值,提取重要值大于1.0,且频度大于15的优势种进行种间关联、生态位重叠和相似性等分析。
1.3 种间联结分析
总体联结性检验 采用方差比率()法判断植物群落的总体关联性,以统计量检验联结性的显著度[18]。参考江焕等[17]的方法计算和。> 1表示种间联结为正关联,<1则表示负关联。
种间联结关系的检验 运用2检验对总体联结性的显著程度进行检验[19]。选取联结系数(AC)对花吊丝竹的种间联结性进行检验。为了提高指标的精确性,采用能反映物种间正联结强弱程度的共同出现百分率(PC),能精确反映种对间相伴共现的几率和联结性的Ochiai指数(OI)和Dice指数(DI)[5]。参考江焕等[17]的方法计算2检验、AC、PC、OI和DI指数,并评估种间联结的关联性。
群落稳定性分析 运用改良的贡献定律法[17]。将群落优势种的频度换算为相对频度,并依次排列,逐步累积频度;将物种总和取倒数,并按照频度排列数据对应进行逐步累积;以物种倒数累计值为横坐标,以物种累计频度为纵坐标,绘制散点平滑曲线模型和直线方程模型(=100-),两条线的交点即为群落稳定值。交点坐标越接近(20, 80),群落就越稳定[20–21]。
花吊丝竹与伴生物种的生态位分析 1个样方作为1个综合资源位,14个优势种在各资源位的生态位宽度、生态位重叠指数和生态位相似性指数作为评估物种对环境资源的利用情况。采用Shannon-Wiener指数计算生态位宽度[22],生态位相似性指数采用Schoener指数[23],生态位重叠指数采用Pianka生态位重叠指数[24]。
1.4 数据的统计和分析
运用Excel 2013对数据进行统计和整理,运用R 3.6.1的SPAA包完成种间联结分析并绘图。
2 结果和分析
2.1 总体联结性和群落稳定性
花吊丝竹所在的12个群落共有30种植物,其中重要值大于1且频率大于15%的优势木本植物有14种(表1)。其中,禾本科6种、豆科5种、大戟科2种、锦葵科2种、榆科2种。入侵物种有8种,主要有马缨丹()、木麻黄、光荚含羞草()等。相对频度以花吊丝竹最高,达16.67%,马缨丹其次,为13.64%。马缨丹的相对多度最高(30.15%),其次为花吊丝竹(25.23%)。木麻黄的相对显著度最高,达86.89%。木麻黄的重要值最高(37.01),其次为花吊丝竹和马缨丹;黑面神()、女贞()和赛葵()的重要值相对较低。
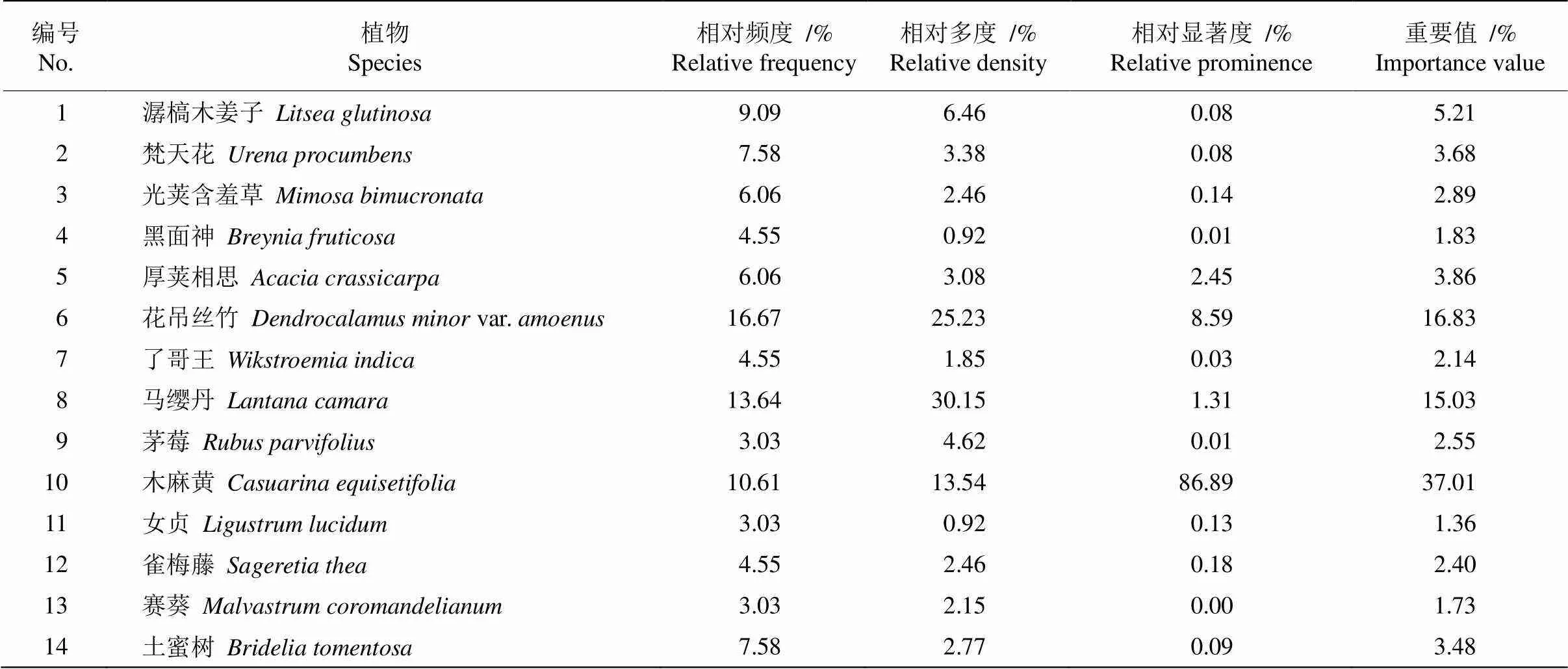
表1 14个优势种的重要值
群落稳定性分析见图1,曲线与直线交点坐标为(39.20, 60.80)。群落内主要优势种的种间呈现不显著的正关联关系(=1.72,=20.625)。
2.2 主要种对间的关联性
14种优势种共组成91个种对(表2)。91个种对的2统计量均小于3.841,表明91个种对间的联结不显著,其中,有19个种对的2统计量为0.00。
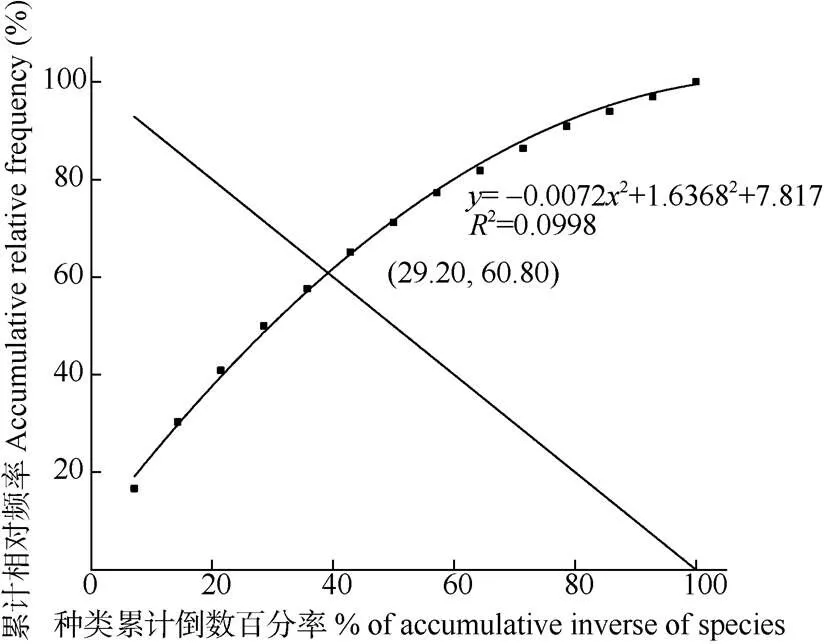
图1 群落的稳定性
种间联结系数(AC)大于0.7的种对有15对, AC小于-0.7的有19对(图2)。花吊丝竹与了哥王()、马缨丹、茅莓()、木麻黄、女贞、雀梅藤()、赛葵、土蜜树()均存在强的正联结性,与光荚含羞草存在强的负联结性,与厚荚相思()的联结性相对较弱。
花吊丝竹和马缨丹、梵天花()和木麻黄表现出非常紧密的关联性,2个种对相伴出现概率很高(PC≥0.7, OI≥0.8, DI≥0.8)。91种对中有7种对表现出相对较高的共现率(0.5≤PC< 0.7), 占总种对数的7.69%;19种对表现出极低的共现率(PC<0.1),占总种对数的20.88%;有20种对表现出极低的共现率和关联性(OI<0.2, DI<0.2)。
2.3 生态位相似性和生态位重叠指数
群落中生态位相似性和生态位重叠指数见表3。花吊丝竹的生态位宽度最高(2.25),赛葵的最低(0.60),马缨丹、木麻黄、潺槁木姜子()、梵天花的相对较高。生态位重叠指数大于0.5的有28对,小于0.1的有21对,几乎不存在竞争关系。生态位相似性指数大于0.5的有16对,小于0.1的有23对。花吊丝竹的生态位重叠指数为0.21~0.78,与茅莓和赛葵的生态位重叠指数和生态位相似性较低。花吊丝竹与马缨丹、木麻黄、土蜜树的生态位重叠指数和相似性指数均大于0.5。
2.4 生态位与种间联结指数的回归分析
物种的生态位宽度与重要值具有显著的正相关关系(2=0.37,<0.05, 图4)。生态位重叠值与AC、DI、PC、OI等指数具有极显著的正相关关系(<0.01)。生态位重值叠与生态位相似性指数具有极显著的正相关关系(2=0.91,<0.01)。
3 结论和讨论
3.1 种间联结关系
物种的生态习性及其对生境需求是影响种间联结的重要因素[25]。本研究的14种优势种中木麻黄和花吊丝竹占据林冠层,其他物种主要生长在林下层,即木麻黄和花吊丝竹在光照资源的获取上具有相对的优势。不同物种在空间上形成垂直结构,在一定程度上减少了生态位竞争。种间联结呈现正联结性,表明种对间的生态习性较为相似,对环境资源的需求较为一致,能够互不干扰,共同生长[26]。花吊丝竹与木麻黄、雀梅藤、土蜜树、黑面神等均存在强的正联结性(AC>0.7)。这说明花吊丝竹与其他物种的生态习性和生境需求较为相似。滨海防护林土壤干旱,营养贫瘠,地势平坦,生境单一。花吊丝竹与木麻黄、雀梅藤、土蜜树、黑面神均为喜阳耐旱植物,能较好地适应滨海防护林的生态环境。有研究表明,种间联结不仅受生态习性等因素的影响,物种多度也是影响种间关联的重要因素[2]。本研究结果支持这一结论,马缨丹、花吊丝竹、木麻黄等具有较高的相对多度(表1),表明在滨海防护林中,种对间能相互促进,物种多度逐渐增加,从而形成较强的正关联。
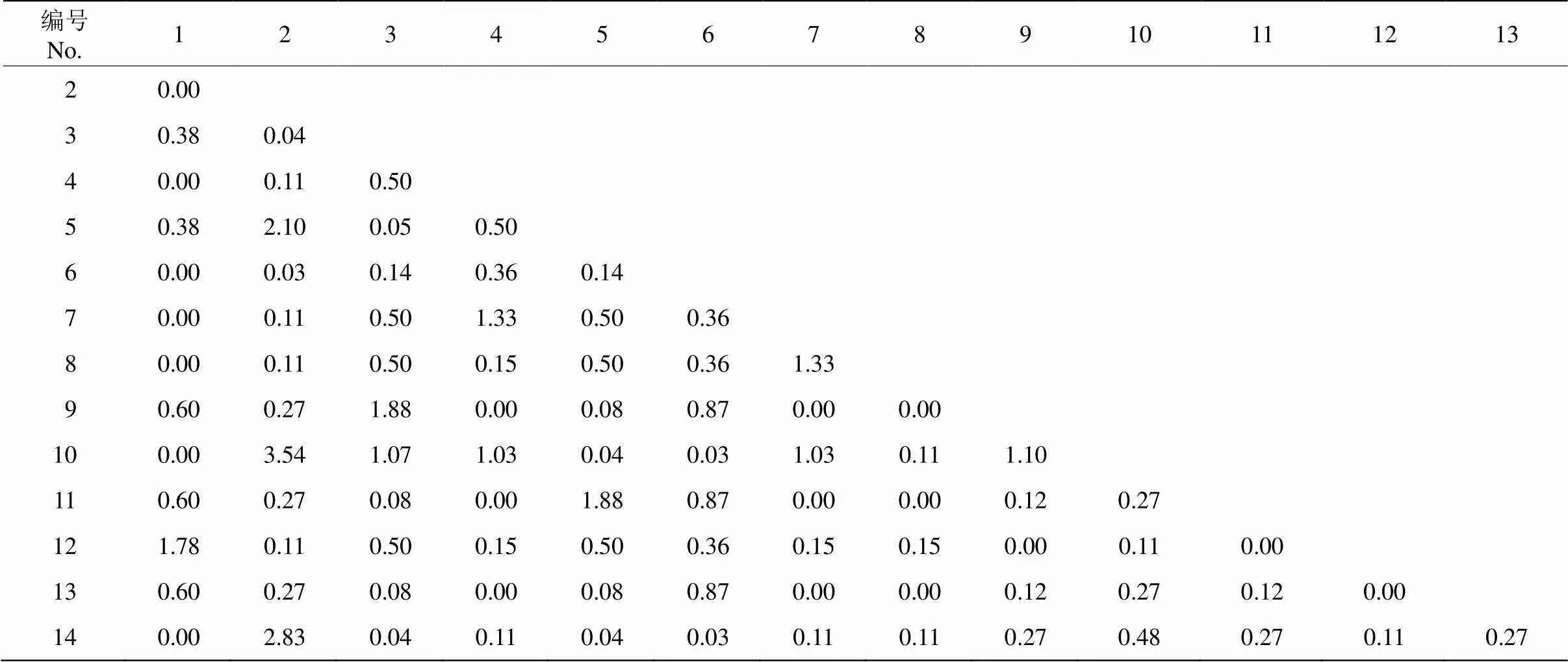
表2 群落优势种间χ2统计量检验
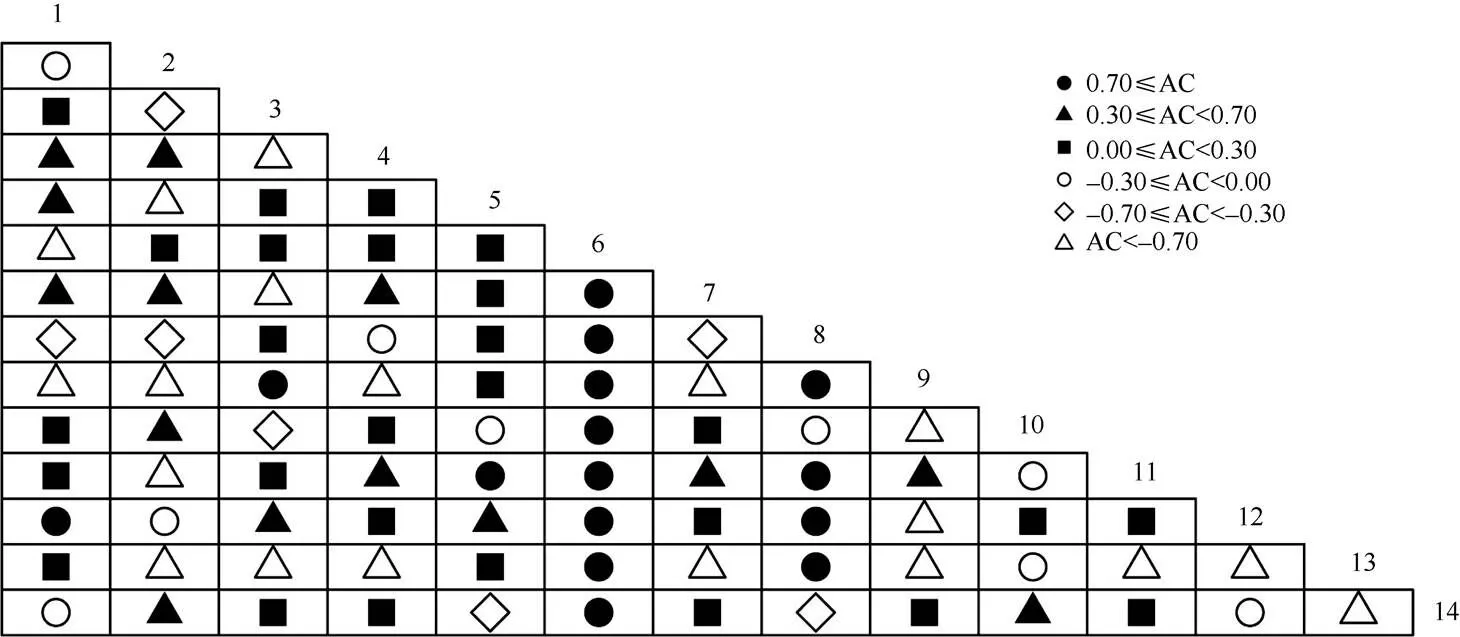
图2 群落主要优势种种间联结系数(AC)半矩阵图。1~14见表1。下图表同。
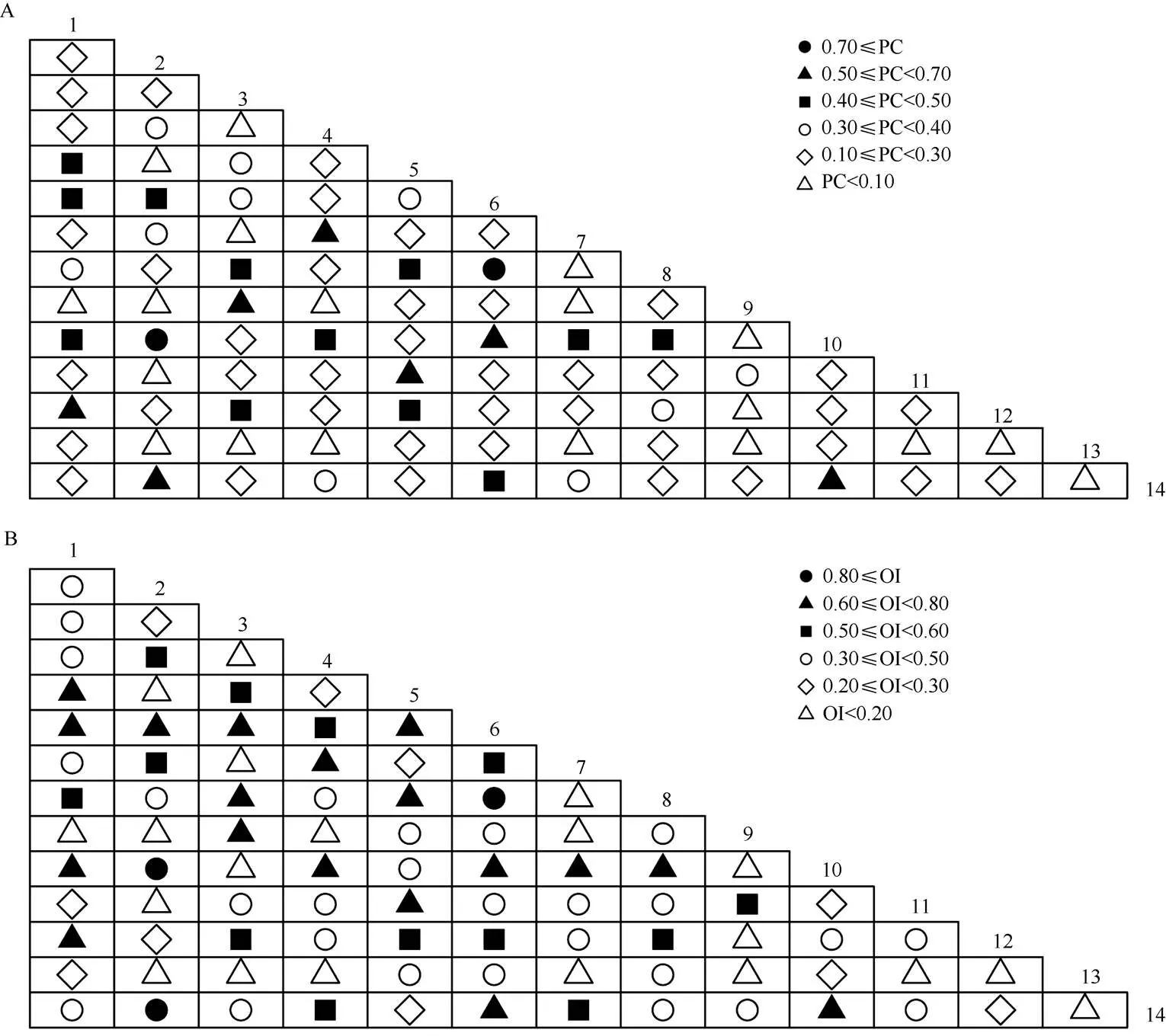
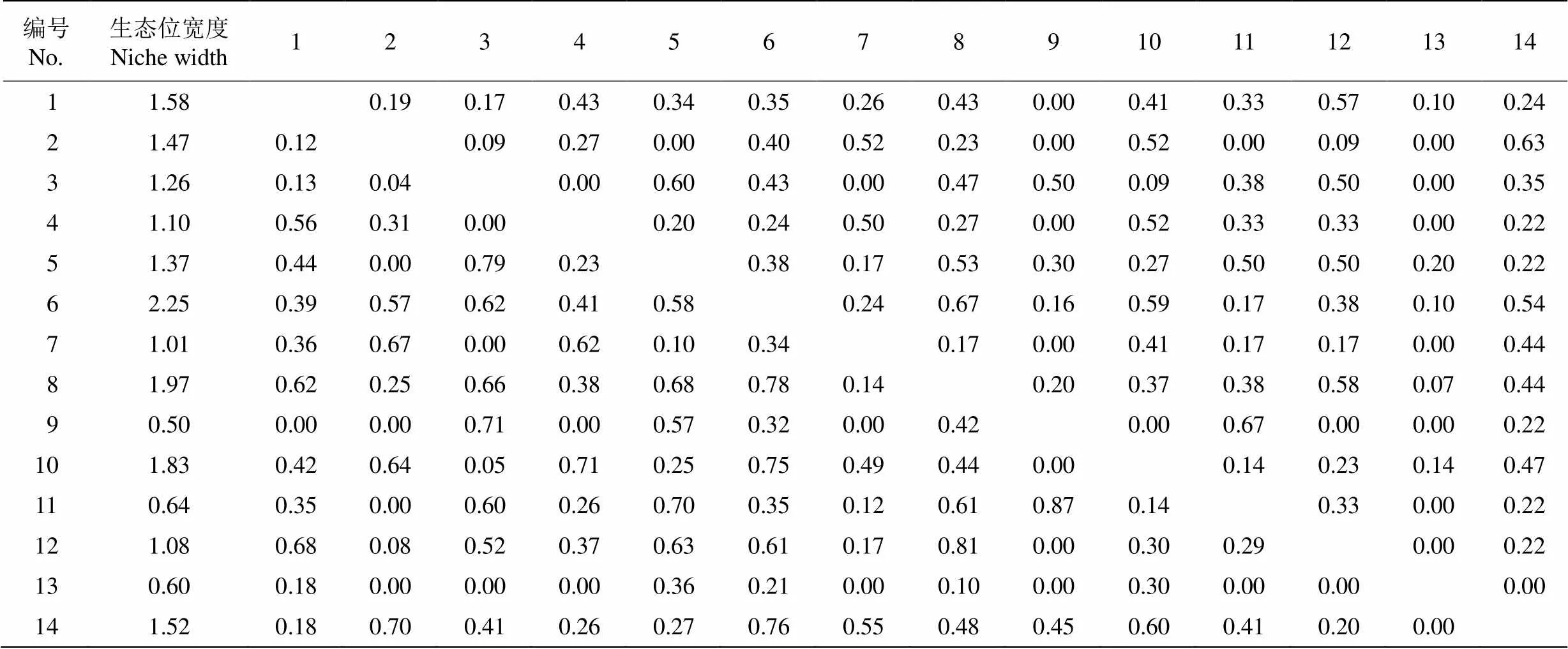
表3 群落主要优势种间的生态位相似性比例(右上)和生态位重叠指数(左下)
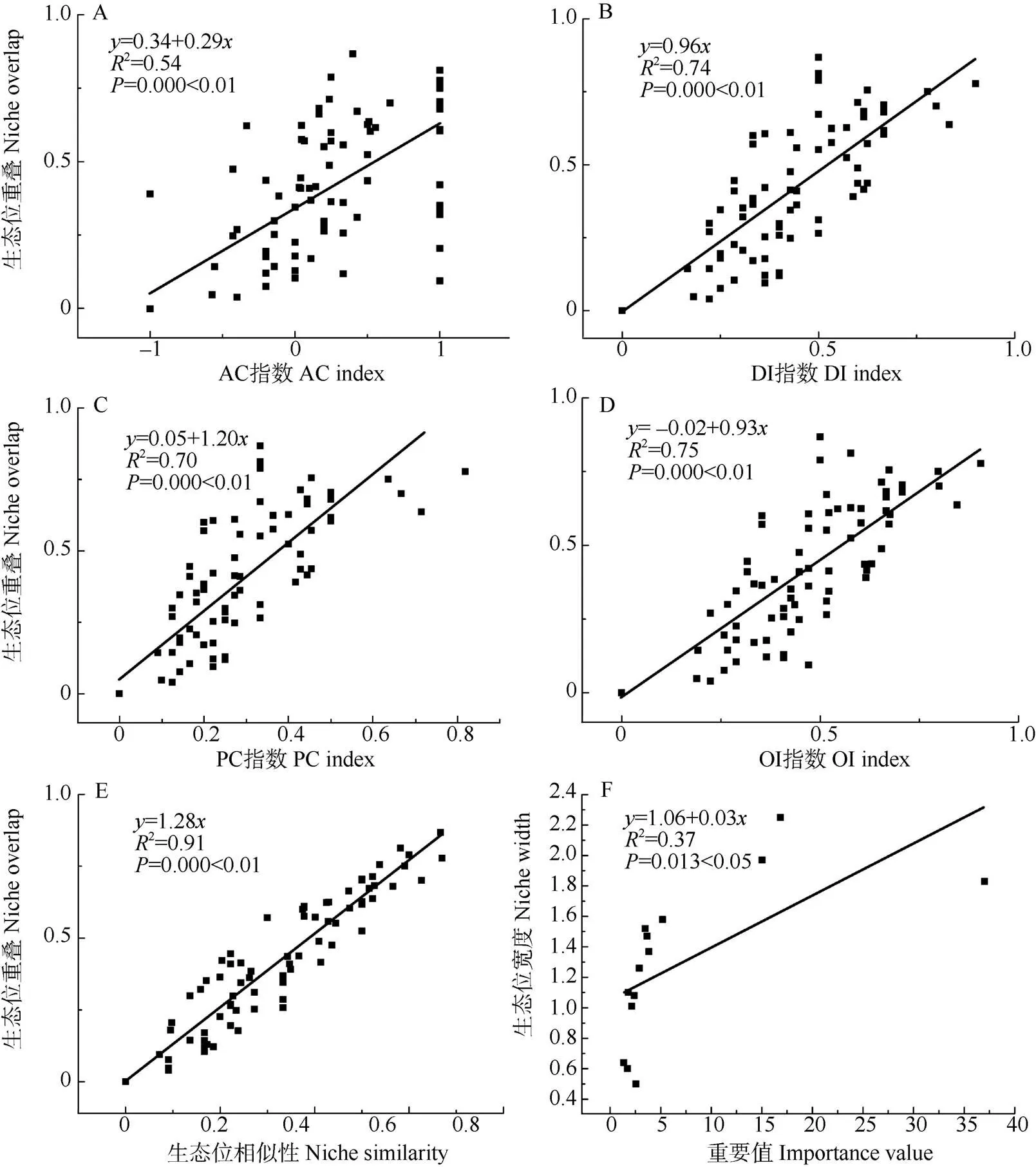
图4 生态位重叠值与种间联结指数的回归分析
3.2 种间联结与生态位的相关性
生态位宽度反映了物种对环境资源的利用情况[19]。花吊丝竹的生态位宽度最高(2.25),在群落中占绝对优势,对环境资源利用更为充分,具有较强适应能力。马缨丹、木麻黄、潺槁木姜子、梵天花的生态位宽度相对较高,对生境资源的竞争较为紧张。花吊丝竹和马缨丹、梵天花和木麻黄表现出非常紧密的关联性,2个种对的相伴出现概率很高。花吊丝竹与马缨丹、木麻黄、土蜜树间的生态位重叠指数和相似性指数大于0.5,即这些种对具有较高的关联性和共现性。由此表明,生态位宽度高的物种,生态位重叠指数也相对较高,这些物种通常具有相似的生境偏好。大尺度上(>20 m)的种对生境偏好与种间联结有很好的一致性[27]。生态位与种间联结存在一定的潜在相关性[28],在荒漠盐碱地植被中,优势植物的生态位重叠指数与种间联结存在显著的正相关[29]。但也有研究表明,生态位宽度与生态位重叠并非呈现绝对的正相关[30]。即便是生态位宽度小的物种也有很高的生态位重叠指数[31]。本文更支持前者的观点,生态位与种间联结存在显著的正相关,生态位重叠指数与AC、DI、PC、OI等指数均呈现极显著的正相关(图4)。在滨海沙地,物种生存受到严酷的环境和短缺的资源影响,生态位宽度越大,物种存活的概率就越高。受到资源的限制,生态位宽度较大的物种必然面临较大的生态位重叠,从而产生较高的共现率和种对间的正联结。
3.3 外来物种对群落稳定性和种间联结的影响
群落内主要优势物种间呈现不显著的正关联关系(=1.72,=20.625),群落稳定性的交点坐标为(39.20, 60.80),离稳定的坐标点(20, 80)有一定距离,即群落总体的种间联结较为松散,群落演替处于不稳定状态,距离顶级群落有一定的距离。种间竞争、环境过滤、人为干扰和外来物种均会对植物群落的稳定性产生影响[1]。群落内的光荚含羞草、马缨丹和木麻黄等植物具有较强的生长优势,表现出一定的入侵趋势和潜力。木麻黄的根系分泌物具有化感作用[32],而马缨丹通过根茎叶和果实释放的化合物对邻近和伴生植物具有明显的化感作用, 表现出一定的入侵潜力[33–34]。花吊丝竹是丛生竹, 相对于散生或混生竹,其扩散能力相对较弱,而外来入侵物种的扩散能力显著较强,导致花吊丝竹对资源的争夺和获取能力大大下降。入侵物种能显著影响群落种间联结及生态位,造成群落处于不稳定的状态[17]。受到外来物种的影响,群落内的生态位宽度低的稀有种、狭域种逐渐被剔除,使得广布的适生物种的相对多度增加,从而与花吊丝竹等形成较强的正关联。花吊丝竹与马缨丹等外来种对间处于较强的正关联,说明了群落演替处于初级演替。但入侵物种对群落的稳定性具有潜在的威胁, 进一步说明了群落演替处于动态的不稳定状态。通过人工的方式清除或抑制马缨丹等入侵物种扩散, 能促进群落的稳定性。引入生态位宽度大,且生态位重叠低的物种也是抑制物种入侵的手段[17],而潺槁木姜子可能是维护群落稳定和演替发展的潜力树种。
[1] XU M H, LIU M, ZHAI D T, et al. A review of contents and methods used to analyze various aspects of plant interspecific associations [J]. Acta Ecol Sin, 2016, 36(24): 8224–8233. doi: 10.5846/stxb20150531 1092.
徐满厚, 刘敏, 翟大彤, 等. 植物种间联结研究内容与方法评述[J]. 生态学报, 2016, 36(24): 8224–8233. doi: 10.5846/stxb201505311092.
[2] LIU X Y, HE D, TIAN W B, et al. Patterns of species associations in woody plants in forest communities of Putuoshan Island, Zhejiang, China [J]. Chin J Plant Ecol, 2017, 41(12): 1219–1227. doi: 10.17521/ cjpe.2017.0170.
刘翔宇, 何东, 田文斌, 等. 浙江普陀山岛森林木本植物的种间关联格局[J]. 植物生态学报, 2017, 41(12): 1219–1227. doi: 10.17521/ cjpe.2017.0170.
[3] DUAN H L, ZHAO A, YAO Z. Species relationship among typical marshland plants in Poyang Lake region on different sample scale [J]. J Trop Subtrop Bot, 2018, 26(5): 449–456. doi: 10.11926/jtsb.3862.
段后浪, 赵安, 姚忠. 不同样方尺度下鄱阳湖湿地典型草洲植物种间关联性[J]. 热带亚热带植物学报, 2018, 26(5): 449–456. doi: 10. 11926/jtsb.3862.
[4] WEI S G, LI L, XU R, et al. Spatial pattern and interspecific relation- ship of dominant species in plant community in Jinggang Mountain [J]. J Trop Subtrop Bot, 2015, 23(1): 74–80. doi: 10.11926/j.issn.1005- 3395.2015.01.011.
魏识广, 李林, 许睿, 等. 井冈山植物群落优势种空间分布格局与种间关联[J]. 热带亚热带植物学报, 2015, 23(1): 74–80. doi: 10. 11926/j.issn.1005-3395.2015.01.011.
[5] CÁRDENAS R E, VALENCIA R, KRAFT N J B, et al. Plant traits predict inter- and intra-specific variation in susceptibility to herbivory in a hyperdiverse Neotropical rain forest tree community [J]. J Ecol, 2014, 102(4): 939–952. doi: 10.1111/1365-2745.12255.
[6] LI X, LIAO L N, YANG H P, et al. Spatial pattern of woody plants in artificial mixed forest ofand[J]. Acta Bot Boreali-Occid Sin, 2019, 39(10): 1842–1850. doi: 10.7606/j.issn.1000-4025.2019.10.1842.
李宪, 廖良宁, 杨海鹏, 等. 杉木与米老排人工混交林群落木本植物空间格局研究[J]. 西北植物学报, 2019, 39(10): 1842–1850. doi: 10.7606/j.issn.1000-4025.2019.10.1842.
[7] LUO J F, ZHENG J M, ZHOU J X, et al. Analysis of the interspecific associations present in an alpine meadow community undergoing reve- getation on the railway-construction affected land of the Qinghai-Tibet Plateau [J]. Acta Ecol Sin, 2016, 36(20): 6528–6537. doi: 10.5846/stxb 201502030276.
罗久富, 郑景明, 周金星, 等. 青藏高原高寒草甸区铁路工程迹地植被恢复过程的种间关联性[J]. 生态学报, 2016, 36(20): 6528– 6537. doi: 10.5846/stxb201502030276.
[8] XUE O, WEI T X. Interspecific association among understory species of the low-efficiency plantation in the Jiufeng National Forest Park [J]. Bull Bot Res, 2016, 36(0): 34–42. doi: 10.7525/j.issn.1673-5102.2016. 01.005.
薛鸥, 魏天兴. 鹫峰国家森林公园低效人工林林下植物种间关联[J]. 植物研究, 2016, 36(1): 34–42. doi: 10.7525/j.issn.1673-5102. 2016.01.005.
[9] SHANG T Q, YE N N, GAO H Q, et al. Community structure analysis of a public welfare forest based on multivariate regression trees [J]. Chin Bull Bot, 2018, 53(2): 238–249. doi: 10.11983/CBB17088.
商天其, 叶诺楠, 高海卿, 等. 基于多元回归树的公益林群落结构解析[J]. 植物学报, 2018, 53(2): 238–249. doi: 10.11983/CBB17088.
[10] Office of Leading Group for Comprehensive Survey of Coastal Zone and Tidar Resources of Fujian Province. Comprehensive Survey Report on Coastal Zone and Tidar Resources of Fujian Province [M]. Beijing: Ocean Press, 1990: 1–575.
福建省海岸带和海涂资源综合调查领导小组办公室. 福建省海岸带和海涂资源综合调查报告[M]. 北京: 海洋出版社, 1990: 1–575.
[11] LIN W X, ZHU W, NIE S, et al. Preliminary study on introduction and application of local tree species in coastal zone of Taiwan island [J]. Chin Agric Sci Bull, 2014, 30(13): 59–65. doi: 10.11924/j.issn.1000- 6850.2013-2903.
林武星, 朱炜, 聂森, 等. 台湾海岸乡土树种引进应用的初步研究[J]. 中国农学通报, 2014, 30(13): 59–65. doi: 10.11924/j.issn.1000- 6850.2013-2903.
[12] LIN A Y. Vegetation characteristics of four types of protection forests on sandy coast in Fujian [J]. J Fujian Agric For Univ (Nat Sci), 2018, 47(1): 66–73. doi: 10.13323/j.cnki.j.fafu(nat.sci.).2018.01.011.
林爱玉. 福建沿海沙地4种防护林类型的群落学特征[J]. 福建农林大学学报(自然科学版), 2018, 47(1): 66–73. doi: 10.13323/j.cnki. j.fafu(nat.sci.).2018.01.011.
[13] YUAN Z L, WEI B L, CHEN Y, et al. How do similarities in spatial distributions and interspecific associations affect the coexistence ofspecies in the Baotianman National Nature Reserve, Henan, China [J]. Ecol Evol, 2018, 8(5): 2580–2593. doi: 10.1002/ece3.3863.
[14] WU L D, LI T T, FU G L, et al. Niche and interspecific relationship of the wild plantwith extremely small population [J]. J Zhejiang Univ (Agric Life Sci), 2020, 46(4): 459–474. doi: 10.3785/ j.issn.1008-9209.2019.08.263.
吴霖东, 李婷婷, 傅国林, 等. 极小种群野生植物浙江安息香的生态位和种间关系[J]. 浙江大学学报(农业与生命科学版), 2020, 46(4): 459–474. doi: 10.3785/j.issn.1008-9209.2019.08.263.
[15] ZHENG J M, CHEN X Y, CHEN L G, et al. Comprehensive evaluation of soil quality at different stand densities ofvar.plantations [J]. Appl Ecol Environ Res, 2020, 18(4): 5985– 5996. doi: 10.15666/aeer/1804_59855996.
[16] HE T Y, FAN L L, TARIN M W K, et al. Physiological and proteomic responses ofvar.(ghost bamboo) under drought stress [J]. Appl Ecol Environ Res, 2020, 18(4): 4817– 4838. doi: 10.15666/aeer/1804_48174838.
[17] JIANG H, ZHANG H, LONG W X, et al. Interspecific associations and niche characteristics of communities invaded by[J]. Biodiv Sci, 2019, 27(4): 388–399. doi: 10.17520/biods. 2019007.
江焕, 张辉, 龙文兴, 等. 金钟藤入侵群落的种间联结及生态位特征[J]. 生物多样性, 2019, 27(4): 388–399. doi: 10.17520/biods.2019007.
[18] SCHLUTER D. A variance test for detecting species associations, with some example applications [J]. Ecology, 1984, 65(3): 998–1005. doi: 10.2307/1938071.
[19] GREIG-SMITH P. Quantitative Plant Ecology [M]. London: Butter- worths, 1957: 1–359.
[20] GODRON M. Some aspects of heterogeneity in grasslands of Cantal [J]. Stat Ecol, 1972, (3): 397–415.
[21] ZHENG Y R. Comparison of methods for studying stability of forest community [J]. Sci Silv Sin, 2000, 36(5): 28–32. doi: 10.3321/j.issn: 1001-7488.2000.05.005.
郑元润. 森林群落稳定性研究方法初探[J]. 林业科学, 2000, 36(5): 28–32. doi: 10.3321/j.issn:1001-7488.2000.05.005.
[22] LEVINS R. Evolution in Changing Environments: Some Theoretical Explorations (MPB-2) [M]. Princeton, NJ: Princeton University Press, 1968.
[23] SCHOENER T W. Resource partitioning in ecological communities [J]. Science, 1974, 185(4145): 27–39. doi: 10.1126/science.185.4145.27.
[24] PIANKA E R. The structure of lizard communities [J]. Annu Rev Ecol Syst, 1973, 4: 53–74. doi: 10.1146/ANNUREV.ES.04.110173.000413.
[25] QUEENBOROUGH S A, BURSLEM D F R P, GARWOOD N C, et al. Habitat niche partitioning by 16 species of Myristicaceae in Amazonian Ecuador [J]. Plant Ecol, 2007, 192(2): 193–207. doi: 10.1007/s11258- 007-9328-3.
[26] WANG B S, PENG S L. Studies on the measuring techniques of interspecific association of lower subtropical evergreen broad-leaved forests: I. The exploration and the revision on the measuring formulas of interspecific association [J]. Chin J Plant Ecol, 1985(4): 274–285.
王伯荪, 彭少麟. 南亚热带常绿阔叶林种间联结测定技术研究: I. 种间联结测式的探讨与修正[J]. 植物生态学与地植物学丛刊, 1985(4): 274–285.
[27] YANG Q S, LIU H M, ZHU T T, et al. Interspecies associations and species-habitat associations in the evergreen broad-leaved forest of Tiantong National Forest Park, Zhejiang [J]. J E China Norm Univ (Nat Sci), 2020(2): 110–119. doi: 10.3969/j.issn.1000-5641.201931004.
杨庆松, 刘何铭, 朱彤彤, 等. 浙江天童国家森林公园常绿阔叶林种间关联和种-生境关联[J]. 华东师范大学学报(自然科学版), 2020(2): 110–119. doi: 10.3969/j.issn.1000-5641.201931004.
[28] CHAI Y F, YUE M. Research advances in plant community assembly mechanisms [J]. Acta Ecol Sin, 2016, 36(15): 4557–4572. doi: 10.5846/ stxb201501140114.
柴永福, 岳明. 植物群落构建机制研究进展[J]. 生态学报, 2016, 36(15): 4557–4572. doi: 10.5846/stxb201501140114.
[29] ZHANG D M, ZHAO W Z, LUO W C. Niche and interspecific association of dominant plant species in saline-alkaline soils of desert steppe zone [J]. Chin J Ecol, 2018, 37(5): 1307–1315. doi: 10.13292/j. 1000-4890.201805.027.
张东梅, 赵文智, 罗维成. 荒漠草原带盐碱地优势植物生态位与种间联结[J]. 生态学杂志, 2018, 37(5): 1307–1315. doi: 10.13292/j. 1000-4890.201805.027.
[30] TIE J, LI Y F, JIN S, et al. Niche characteristics of dominant tree species inhabitat of Lishan National Nature Reserve, Shanxi [J] Chin J Ecol, 2015, 34(3): 634–641. doi: 10.13292/j.1000- 4890.2015.0087.
铁军, 李燕芬, 金山, 等. 山西历山国家级自然保护区猕猴栖息地优势树种生态位特征[J]. 生态学杂志, 2015, 34(3): 634–641. doi: 10.13292/j.1000-4890.2015.0087.
[31] HU Z H, QIAN H Y, YU M J. The niche of dominant species populations inforest in Gutian Mountain National Nature Reserve [J]. Acta Ecol Sin, 2009, 29(7): 3670–3677. doi: 10. 3321/j.issn:1000-0933.2009.07.027.
胡正华, 钱海源, 于明坚. 古田山国家级自然保护区甜槠林优势种群生态位[J]. 生态学报, 2009, 29(7): 3670–3677. doi: 10.3321/j.issn: 1000-0933.2009.07.027.
[32] LI H M, CAO T T, GU M Z, et al. The special bacterial metabolites and allelopathic potentials inwoodland of different stand ages [J]. Chin J Appl Environ Biol, 2016, 22(5): 808–814. doi: 10. 3724/SP.J.1145.2016.01021.
李慧敏, 曹婷婷, 顾美子, 等. 不同林龄木麻黄林地特有细菌代谢产物及其化感潜力[J]. 应用与环境生物学报, 2016, 22(5): 808–814. doi: 10.3724/SP.J.1145.2016.01021.
[33] RANJAN R. Deriving double dividends through linking payments for ecosystem services to environmental entrepreneurship: The case of the invasive weed[J]. Ecol Econ, 2019, 164: 106380. doi: 10.1016/J.ECOLECON.2019.106380.
[34] MISHRA A. Allelopathic properties of[J]. Int Res J Basic Clin Stud, 2015(3): 13–28. doi: 10.14303/irjbcs.2014.048.
Interspecific Association and Niche of Mixed Forest Communities ofandvar.in the Windbreak of Sandy Coast
ZHENG Junming1a, LI Min1a, ZHANG Meng1b, TARIN Muhammad Waqqas Khan1a, HE Tianyou1a, CHEN Lingyan1a, CHEN Liguang1b, HONG Xiaolong2, ZHENG Yushan1a*
(1a. College of Landscape architecture, 1b. College of Forestry, Fujian Agriculture and Forestry University,Fuzhou 350002, China; 2. Chishan Forest farm, Dongshan Country in Fujian Province,Zhangzhou 363400, Fujian, China)
In order to understand the stability of mixed forest community ofandvar.in the coastal windbreak of Dongshan Island, China, the inter-specific association and niche of the community was analyzed by using quadrat method. The results showed that there was no significant positive correlation among inter-specific associations as a whole (=1.72,=20.625), indicating that the community was in an unstable dynamic succession state. Two species pairs;var.and,andhad high association and co-occurrence (PC≥0.7, OI≥0.8, DI≥0.8). The niche width of invasive plantwas high, and the niche overlap withvar.was high, showing high positive correlation, making the population in an unstable state. The habitats of the coastal windbreak was simple, and the habits of dominant species were similar, so there was a strong positive interspecific association betweenvar.and other most species. Restricted by environmental resources, the strong positive association was related to the large niche width of species. The strong positive interspecific association of species pairs and the potential competitive advantage of alien species might be an important factor affecting the stability of the community.
;var.;Population; Inter-specific association; Niche; Sandy coast
10.11926/jtsb.4346
2020-12-03
2021-01-13
福建省区域发展项目(2015N3015); 福建省科技创新团队项目(2018[49]); 福建农林大学科技创新发展基金项目(CXZX2017089); 福建省科技重大专项(2013NZ0001)资助
This work was supported by the Project for Regional Development in Fujian (Grant No. 2015N3015); the Project for Science and Technology Innovation Team in Fujian (Grant No. 2018[49]); the Project for Science and Technology Innovation Development of Fujian Agriculture and Forestry University (Grant No. CXZX2017089); and the Special Project for Key Science and Technology in Fujian (Grant No. 2013NZ0001).
郑俊鸣(1991~ ),男,博士研究生. 研究方向:园林植物应用。E-mail: 2181775005@fafu.edu.cn
. E-mail:zys1960@163.com
