籽粒镉低积累水稻地上部镉高积累遗传特性分析
2021-10-11杨安琪余海英黄化刚张锡洲李廷轩
杨安琪,余海英,黄化刚,张锡洲,李廷轩
四川农业大学资源学院,成都 611130
镉(Cd)是一种毒性极强的重金属元素,其生物有效性高、迁移性强[1]。随着工业化和城镇化进程的加快,农田土壤Cd污染日益严重,且易随食物链进入人体、危害人类健康[2-3]。水稻是人类膳食Cd的主要来源,其平均Cd含量显著高于其他谷类食物[4]。因此,安全利用Cd污染农田土壤、降低糙米Cd含量成为亟待解决的粮食安全问题。选育籽粒Cd低积累水稻能保障Cd污染农田的安全生产,同时一些生物量大、地上部Cd积累能力较强的水稻品种表现出较好的Cd修复潜力[5-6],因而选育地上部Cd高积累、籽粒Cd低积累的水稻品种利于实现Cd污染农田的修复和安全生产。明晰水稻Cd积累的遗传机制是选育籽粒Cd低积累或地上部Cd高积累水稻的重要前提。不同水稻品种Cd积累存在明显的基因型差异,利于寻找相关功能基因[7-8]。研究发现,水稻Cd积累相关性状属数量性状,遗传机制复杂,同时受环境因素影响,因此不能仅通过表型对基因型进行选择[9]。QTL定位通过建立数量性状表型值与DNA分子标记间的关系,可以确定各个QTL位点在染色体上的位置、效应及其相关作用[10-11],使复杂数量性状的遗传改良和分子操纵成为可能,具有较大的应用前景[12-13]。近年来,利用Cd积累能力差异显著的水稻品种构建的F2、RIL、CSSLs等群体进行QTL定位,在水稻12条染色体上发现了控制水稻籽粒和地上部Cd积累相关的QTLs[14-16]。Xue等[17]利用DH群体,在第7号染色体上得到控制地上部Cd含量的QTLqCDS7。Ueno等[18]和Tezuka等[19]利用不同亲本构建的F2群体,也在第7号染色体上得到控制地上部Cd积累量的QTL,其中QTLqCdT7包含的基因OsHMA3发生碱基突变,以该基因作为分子标记,筛选出了Cd污染农田修复效果显著的水稻品种Akita 110[19-22]。除第7号染色体外,Ueno等[18]和Yan等[23]分别还在第2、5、11和10号染色体上定位到控制水稻地上部Cd积累量的QTL。此外,还有研究发现水稻Cd积累相关QTL如qCd-2、qCd-7、GCC7、qCd1-3、CAL1等影响籽粒、叶片、根等部位Cd含量[14-15,24-25]。可见,通过QTL定位挖掘水稻Cd积累的关键基因有助于阐明水稻Cd积累的遗传机制,为利用分子标记辅助选择育种奠定基础。相同土壤环境条件下,相较于其他籽粒Cd低积累水稻品种,雅恢2816具有更强的地上部Cd积累能力,可用于Cd污染农田边修复边生产。前期已对其籽粒Cd低积累特性进行了探讨[26-28],但其地上部Cd积累的遗传稳定性和遗传机制尚不清楚。利用雅恢2816杂交后代F1和F2,分析其地上部Cd积累相关性状的杂种优势,挖掘控制地上部Cd积累相关性状的QTL,明确其地上部Cd积累机制,为分子标记辅助选择籽粒Cd低积累且地上部Cd高积累材料提供理论依据。
1 材料与方法
1.1 供试材料
水稻雅恢2816(R)、泸98A(S1)、5406A(S2)、C268A(S3)、泸98A/雅恢2816(H1)、5406A/雅恢2816(H2)、C268A/雅恢2816(H3)、F1群体(包含雅恢2816与3个不育系材料(泸98A、5406A、C268A)分别杂交得到的第一代群体H1、H2和H3)、F2群体(共120个株系,为杂交组合H3的自交后代)。由四川农业大学农学院提供。雅恢2816是四川省常用杂交生产的籼稻品种,具有地上部Cd高积累和籽粒Cd低积累特性;泸98A(S1)、5406A(S2)和C268A(S3)均为不育系水稻材料,其地上部Cd含量与积累量在盆栽和大田试验中均显著低于雅恢2816[26,28]。
1.2 试验设计与处理
第一期试验共设21个试验小区(1 m×2 m),种植亲本及F1,每个材料设3次重复。第二期试验共设6个试验小区和1个试验大区(1 m×4 m),分别种植亲本(雅恢2816、C268A)和F2。各小区随机区组排列,各区间设20 cm缓冲带。每小区种植水稻32穴,每大区种植水稻120穴,每穴1株,株行距为20 cm×25 cm。三叶期栽插,采用旱地育秧,于当年4月播种,5月移栽,按当地习惯进行水肥管理和病虫害防治。
试验区位于四川省成都平原某市的Cd污染农田,属中纬度亚热带湿润气候区,海拔507 m,年均温15.9—16.1 ℃,年均降水量900—1000 mm。试验土壤基本理化性质为pH 6.60、有机质34.61 g/kg、全氮2.43 g/kg、碱解氮100.31 mg/kg、有效磷19.03 mg/kg、速效钾69.25 mg/kg、全Cd 1.92 mg/kg、有效Cd 0.63 mg/kg。
1.3 样品采集与制备
于水稻抽穗期,分单株采集雅恢2816、C268A和F2幼叶鲜样于2mL灭菌灭酶冻存管中,经液氮冷冻后储存于-75℃超低温冰箱中,用于DNA提取。于水稻成熟期采样,将样品用自来水冲洗干净,根部用20 mmol/L Na2-EDTA溶液浸泡15 min去除根表附着的Cd2+,再用去离子水洗净,最后用吸水纸擦干。将植株分为根、茎、叶、穗,根、茎、叶于105 ℃下杀青30 min后,75 ℃烘干至恒重,称重后粉碎备用,穗部经风干称重后,脱粒制成糙米备用。
1.4 测定项目及方法
土壤基本理化性质采用常规分析方法[29];土壤全Cd含量采用HNO3-HClO4-HF(5∶1∶1,体积分数)消化,有效态Cd含量采用DTPA提取,植株Cd含量采用HNO3-HClO4(5∶1,体积分数)消化,火焰原子吸收分光光度计(AA400P,Analytikjena,Germany)测定。
1.5 基因型分析
采用Takara试剂盒提取叶片DNA。利用简化基因组测序技术(Genotyping by sequencing)进行基因型鉴定。内切酶组合为EcorI/NIaIII(New England Biolabs,Ipswich,MA)。
1.6 QTL定位
利用已构建的遗传连锁图谱和表型数据[27],采用WinQTL Cartographer v2.5 程序包的复合区间作图法(Composite Interval Mapping,CIM)进行QTL定位分析。选择0.5 cM的扫描区间,在整个遗传图谱上逐步扫描寻找QTL。同时,选择距离待检测区间至少10 cM,与性状相关性显著性最强的前15个标记作为协变量,校正背景QTL效应对目标区间QTL分析的影响。以LOD值2.5作为阈值来判断QTL是否存在,若标记区间中LOD>2.5,则认为该区间LOD最高处所对应的位点为一个QTL,并估计各个QTL对表型变异的贡献率。QTL的命名基本按照McCouch等[30]的方法并稍作改动:q+目标性状英文缩写名称(首字母大写)+“-”+连锁群的序号+“-”+同一连锁群上QTL数,本文中QTL名称均用斜体表示。qSB指控制指地上部生物量的QTL;qSCdA指控制指地上部Cd积累量的QTL。
1.7 数据处理
地上部Cd积累量=茎生物量×茎Cd含量+叶生物量×叶Cd含量,超亲优势=(F1表型值-高值亲本表型值)/高值亲本表型值×100%,中亲优势=(F1表型值-双亲表型平均值)/双亲平均值×100%。
统计分析在SPSS 24.0中进行,多重比较选择LSD法;图表制作采用Origin 8.0和Excel 2013。
2 结果与分析
2.1 F1地上部Cd积累相关性状的杂种优势
亲本及其F1地上部生物量、Cd含量和Cd积累量差异显著(表1,表2)。总体而言,F1地上部生物量显著高于雅恢2816,除H2,杂交组合H1和H3与其不育系母本无显著差别。H1、H2和H3地上部生物量分别为雅恢2816的1.43、1.35和1.43倍。所有杂交组合地上部Cd含量显著低于雅恢2816,显著高于不育系母本。所有杂交组合地上部Cd积累量显著高于各不育系母本,与雅恢2816无显著差别。H1、H2和H3地上部Cd积累量分别为雅恢2816的1.17、1.16和1.16倍。F1地上部Cd积累相关性状表现出较好的中/超亲优势,Cd积累相关性状杂种优势受遗传效应的影响,C268A/雅恢2816后代表现较好,可利用其F2进行遗传特性分析。
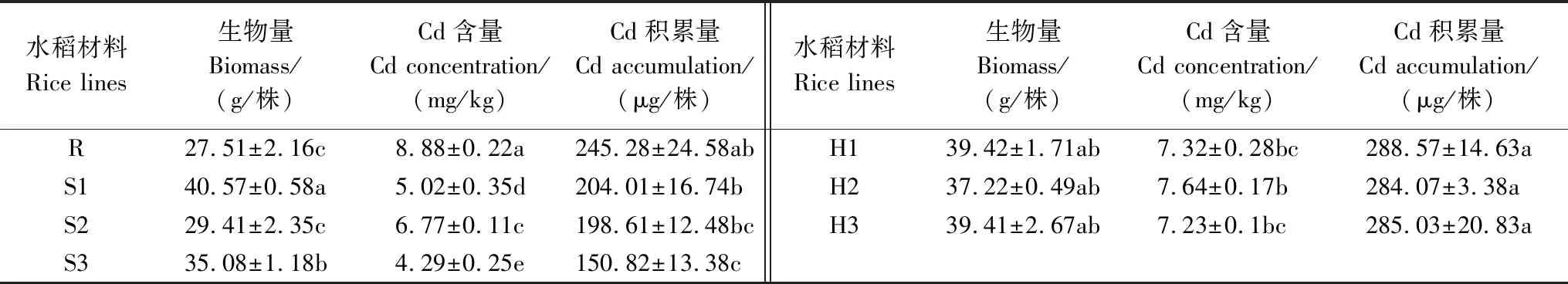
表1 亲本及其F1地上部Cd积累相关性状Table 1 Comparison of differences of traits related to shoot Cd accumulation between rice parents and their F1 hybrids
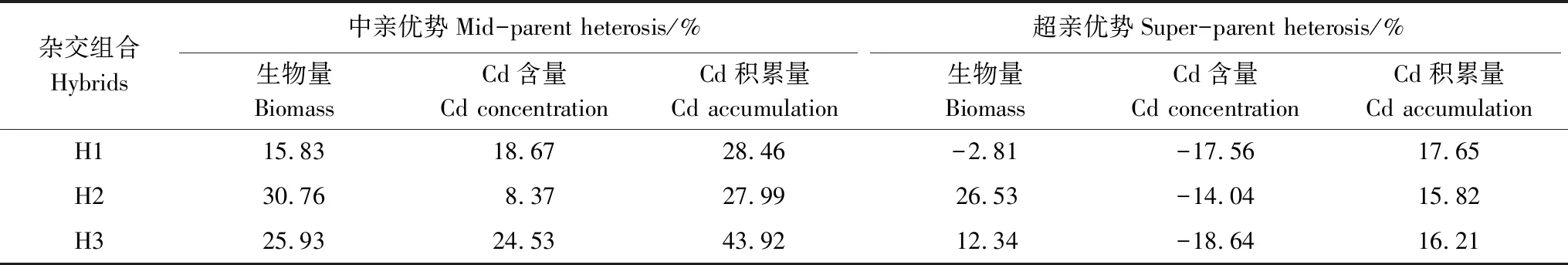
表2 F1地上部Cd积累相关性状的中/超亲优势Table 2 Mid-/Super-parent heterosis of traits related to shoot Cd accumulation in F1 hybrids
2.2 F2地上部Cd积累相关性状的遗传特性分析
雅恢2816地上部Cd含量、Cd积累量显著高于C268A,生物量无显著差异(表3)。雅恢2816地上部Cd含量和Cd积累量分别为C268A的2.17和2.03倍。F2地上部生物量、Cd含量和Cd积累量表现为连续分布和中/超亲分离,变异系数分别为51.14%、15.18%和57.79%,其中Cd积累量变异较大。说明地上部Cd积累相关性状均为数量性状,适合进行QTL定位。
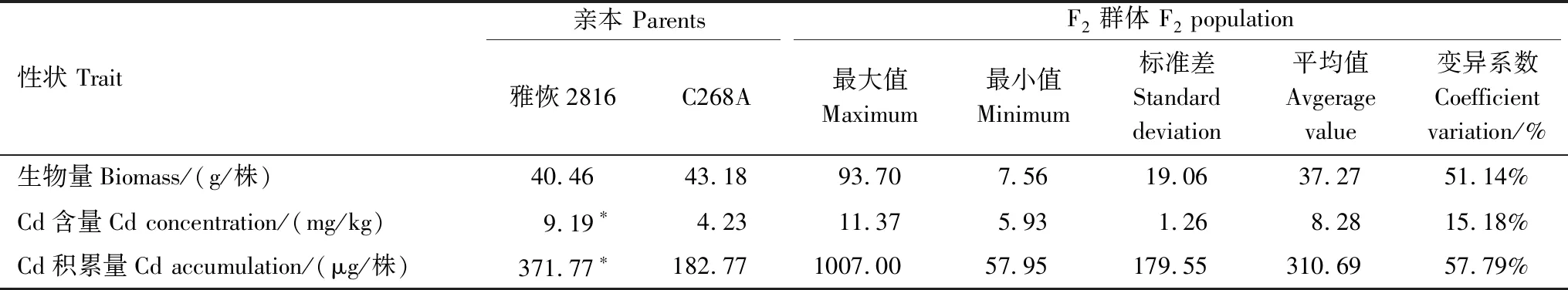
表3 亲本与F2地上部Cd积累相关性状的表型变异Table 3 Phenotypic variation of traits related to shoot Cd accumulation in rice parents and F2 population
对F2地上部Cd积累量与各性状间进行相关性分析(表4),除根和糙米Cd含量外,地上部Cd积累量与地上部生物量、Cd含量,根、糙米的生物量、Cd积累量和根-地上部转移系数呈极显著正相关,与地上部-籽粒转移系数呈极显著负相关。说明水稻地上部Cd积累量与地上部生物量、Cd含量,根、糙米的生物量、Cd积累量和根-地上部转移系数、变化趋势相同,而与根-地上部转移系数变化趋势相反。
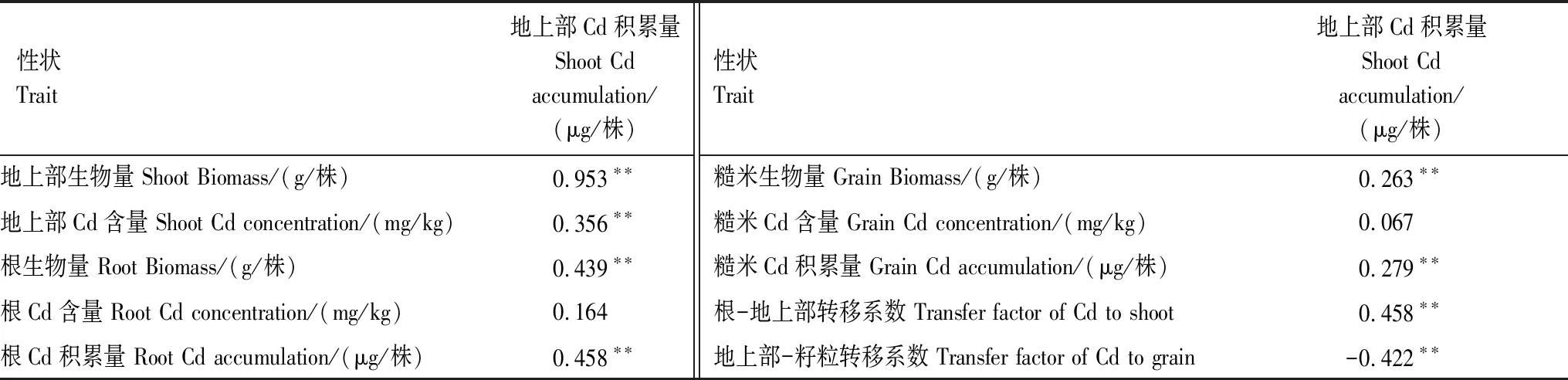
表4 F2地上部Cd积累量与各器官Cd积累相关性状之间的相关性分析Table 4 Correlation analysis between shoot Cd accumulation and traits related to organs Cd accumulation in F2 population
2.3 F2地上部Cd积累相关性状QTL定位
利用C268A与雅恢2816构建的F2作图群体,共定位到4个与地上部生物量和Cd积累量紧密连锁的主效QTL位点(表5,图1)。其中,地上部生物量紧密连锁的QTLqSB-6位于第6号染色体,物理区间大小为4.28cM,表型贡献率为14.4%。3个控制地上部Cd积累量的QTLqSCdA-4、qSCdA-6-1和qSCdA-6-2分别位于第4、6和6号染色体,其物理区间大小分别是9.64、13.5和2.43cM,表型贡献率分别为11.7%、14.4%和10.6%。4个QTL的加性效应均为负,表明控制增加地上部生物量及Cd积累量的基因均来自父本雅恢2816。在第6号染色体上,区间marker04171-marker04197同时控制地上部生物量与Cd积累量,其物理区间大小为3.28cM。

表5 F2地上部Cd积累相关性状QTL定位Table 5 QTL mapping of traits related to shoot Cd accumulation in F2 population

图1 F2地上部Cd积累相关性状QTL的染色体定位Fig.1 Chromosome location of QTL related to shoot Cd accumulation in F2 populationQTL qT-r-s-6-2、qGB-4和qRCdA-6来源于前期研究结果[27,31]
3 讨论
3.1 水稻地上部Cd积累的杂种优势及遗传机理
Cd胁迫下,植物的正常生长发育会受到影响,首先体现在根、茎、叶生物量的变化[32]。本研究中,杂交组合泸98A/雅恢2816、5406A/雅恢2816和C268A/雅恢2816的地上部生物量显著高于父本雅恢2816,表现出中/超亲优势,表明利用杂交育种,可以明显提高水稻的生物量。同时,各杂交组合地上部Cd含量和Cd积累量显著高于不育系母本,表现出中/超亲优势。前期发现,在Cd污染大田和不同Cd处理的盆栽试验中,各杂交组合籽粒Cd含量显著低于双亲,表现出明显的负向超亲优势[31,33]。研究表明,杂交水稻对Cd的积累主要受双亲遗传背景影响[34]。雅恢2816具有籽粒Cd低积累、地上部Cd高积累特性,可稳定遗传给后代。因此,以上杂交组合可应用于Cd污染农田的边生产边修复。具有杂种优势的性状一般属数量性状,受多基因控制,亲本材料的遗传背景差异可能导致杂交组合Cd积累量存在较大差异[34-35],从分子层面深入挖掘水稻Cd积累的遗传机制可提高杂种优势利用效率。本研究利用地上部Cd积累特性差异显著的亲本材料,在第4和6号染色体上得到控制地上部Cd积累量的QTLqSCdA-4、qSCdA-6-1和qSCdA-6-2,与前人结果比较,是新的控制水稻地上部Cd积累量的主效QTL。
3.2 水稻Cd积累相关性状遗传关系分析
了解水稻地上部与籽粒Cd积累的遗传关系对于实现Cd污染农田边生产边修复目标至关重要。Wang等[36]利用RIL群体,在7号染色体上定位到同时控制水稻地上部Cd含量和糙米Cd含量的QTLqCd7.1。Ishikawa等[37]利用BILs群体,在第2号染色体上控制籽粒Cd含量的QTLqGCd2与控制地上部Cd含量的QTLqSCd2紧密连锁,在第7号染色体上控制籽粒Cd含量QTLqGCd7和地上部Cd含量的QTLqSCd7完全重叠。Hu等[38]与Abe等[39]在3号染色体上分别定位到控制糙米Cd含量的QTLqCCBR3和控制地上部Cd含量的QTLqlGCd3,二者存在重叠区域。Ueno等[18]和Liu等[15]利用不同的作图群体,以水稻地上部Cd积累量与糙米Cd含量为指标进行QTL定位,在第2号染色体上,两个QTL存在重叠部分。本研究中控制水稻地上部Cd积累量的QTL与前期得到的4个控制糙米Cd含量的QTL位点位于不同的染色体[27],与Wang等[36]和Ishikawa等[37]的研究结果不同,且与其他学者比较,也并无重叠区域,与Hu等[38]和Abe等[39]、Ueno等[18]和Liu等[15]结果不同。这可能与不同研究的双亲遗传差异和群体类型不同有关[40]。本研究中,控制糙米Cd含量和地上部Cd积累量的QTL增效等位基因来源不同,且定位于不同染色体,表明源于雅恢2816的等位基因不会同时增加地上部Cd积累量和糙米Cd含量。
水稻地上部Cd积累量与各器官生物量、Cd积累量、转移系数之间存在明显的相关性,且在4号和6号染色体出现QTL集簇区Cl4-1、Cl6-1、Cl6-2和Cl6-3。在4号染色体上,集簇区Cl4-1发生qGB-4与qSCdA-4重叠,且加性效应一致[31],表明Cl4-1能增加地上部Cd积累量并提高稻米产量。经基因预测,在集簇区Cl4-1内含有基因Os04g0613000(OsZIP3)。研究发现,OsZIP3在水稻节点表达较高,负责扩大维管束的木质部中Zn的卸载,从而调控Zn在地上部的分配[41]。Cd和Zn具有相似的化学性质,一些Zn转运蛋白同时负责Cd的转运[42],因而推测OsZIP3可能参与了Cd的分配,进一步研究Os04g0613000的功能,有助于解释雅恢2816地上部Cd积累机制。亲本雅恢2816具有较高的根-地上部转移系数,且地上部-籽粒转移系数较低,使其具有籽粒Cd低积累、地上部Cd高积累特性。相关研究表明,OsHMA2和OsHMA3在Cd从根向地上部的转运过程中起到重要作用,OsHMA2参与Cd向木质部的装载[43],而OsHMA3能将Cd区隔化在根细胞的液泡中[20,44]。但在本研究中同时控制根-地上部转移和地上部Cd积累量的集簇区Cl6-1不包含OsHMA2和OsHMA3,有望在集簇区Cl6-1挖掘到类似功能的新基因。集簇区Cl6-2发生qSCdA-6-1、qSB-6、qRCdA-6重叠,Cl6-3 发生qSCdA-6-1、qRCdA-6重叠[31],F2地上部Cd积累量与地上部生物量、根Cd积累量呈极显著正相关,可见增加地上部生物量和根系Cd积累量,能有效提高水稻对Cd的提取效率。控制数量性状的QTL在染色体上成簇分布的现象可见于绝大多数研究中,这可能是同一基因控制不同性状的表达,也可能是控制多个性状表达的基因紧密连锁的结果[39,45],后续可针对重叠区段进行深入研究,以期挖掘控制地上部Cd积累的关键基因。
4 结论
籽粒Cd低积累亲本材料雅恢2816具有地上部Cd高积累特性,且能稳定遗传。地上部Cd积累相关性状由多基因控制,F2代中/超亲优势明显。QTLqSB-6控制地上部生物量,QTLqSCdA-4、qSCdA-6-1和qSCdA-6-2控制地上部Cd积累量。区间marker04171-marker04197同时关联着水稻地上部生物量(qSB-6)与地上部Cd积累量(qSCdA-6-1),且与控制籽粒Cd含量的QTL未发生重叠,该区间为后期利用分子标记辅助育种同时实现籽粒Cd低积累、地上部Cd高积累提供可能。
