入侵性红耳滑龟与土著乌龟幼体游泳、翻身及心率表现的差异
2021-10-11张辰欢康春全黄心怡陆洪良
张辰欢,康春全,黄心怡,陆洪良
杭州师范大学生命与环境科学学院,生态系统保护与恢复杭州市重点实验室,杭州 311121
人为地将一些具入侵性的外来物种引入到适宜其生活且缺乏天敌的新环境中时,它们可能会在新栖息地建立种群,破坏当地自然生态系统,从而产生导致被侵入地土著物种的灭绝、生物多样性的下降等系列环境问题[1-2]。外来种为何能在其分布区之外的环境中成功入侵是研究者最关心的问题。在用于解释外来种入侵机制的多种假说中,入侵种在生活史、生理、行为等表型特征上比土著物种具内禀优势应是认同度较高的一个观点[3]。此外,入侵种的这些表型特征的可塑性程度较高对其在新环境快速适应亦可能起到了关键作用[4-5]。动物运动能力、对极端环境的耐受能力是与其适合度密切相关联的指标,在土著-入侵种对比研究中同样被频繁使用。多数的相关研究证实入侵种较亲缘关系相近的土著种常常表现出较强的运动和抗逆耐受能力。例如,入侵美国沿海的地中海贻贝(Mytilusgalloprovincialis)热耐受能力显著高于同属的海湾贻贝(M.trossulus)[6];入侵法国岛屿的一种壁虎-疣尾蜥虎(Hemidactylusfrenatus)的运动速度显著快于当地土著壁虎-哀鳞趾虎(Lepidodactyluslugubris)[7]。外温动物的生理与功能表现常显示出热依赖性,某一特定生理或功能表现随温度变化的趋势可用热功能曲线表示[8]。热功能曲线在不同物种或种群可存在差异。一些研究预测:入侵种比土著种应更具广温性,其生理或功能表现的热敏感性应较低,热功能曲线应相对平缓[6,9]。然而,一些研究所得的结果与该预测存在着偏差。
原产于美国中南部的红耳滑龟(Trachemysscriptaelegans)已被列为全球最危险的100个入侵种之一,在世界范围内许多地区已成功入侵[10]。一些与土著龟类的对比研究显示:入侵当地的红耳滑龟通常具有较好的功能与生理表现。例如,成功入侵欧洲的红耳滑龟比当地土著龟类-地中海拟水龟(Mauremysleprosa)具更快速的翻身表现及较强的低温耐受能力[11-12]。红耳滑龟在我国南方各地的入侵形势也同样严峻,然而与我国土著龟类相互作用的研究报道仍非常有限[13]。仅有的少量研究结果显示:红耳滑龟比我国土著龟类(中华花龟M.sinensis、乌龟M.reevesii)具较快的游泳速度及较强的对环境污染物的耐受能力[14-15]。本研究将采自我国海南、浙江不同养殖群体的红耳滑龟和乌龟卵,在相同温湿度条件孵化,孵出幼体在多个温度下测定运动表现(游泳速度、翻身时间)和心率,并构建热功能曲线。通过比较两种龟的功能表现及其热敏感性,以期解答以下几个预测:(1)若入侵龟在运动能力、代谢水平等相关特征上较土著龟表现出其优势性,那么红耳滑龟幼体的游泳速度、翻身反应及心率应快于乌龟幼体;(2)若入侵龟较土著龟更具广温性,能在相对大的温度范围内维持其功能表现,那么红耳滑龟幼体的运动及心率表现的热功能曲线比乌龟幼体应显示较低热敏感性(较大的热表现宽度);(3)若入侵龟的功能表现较土著龟具更大的可塑性,那么来自不同养殖群体的红耳滑龟幼体运动及心率表现受温度变化的影响幅度应大于乌龟幼体。
1 材料与方法
1.1 龟卵收集与孵化
实验用160枚红耳滑龟和乌龟卵于2017年6月中旬购自海南海口和浙江海宁的龟类养殖场(两种龟每个养殖场各40枚)。龟卵为产出后1—2 d收集,随后所有卵被运至杭州师范大学实验室。卵称重、编号后随机分配于不同的孵化盒,每个孵化盒20枚卵(来自相同养殖群的同种龟卵放入同一孵化盒)。孵化盒内含潮湿基质(-220 kPa,蛭石∶水=1∶1[16]),卵白斑朝上半埋于基质。所有孵化盒均置于一个温度预先设置为29 ℃的人工气候箱(宁波莱福科技有限公司),每3 d补充基质散失的水分以保持基质湿度恒定,并按事先设置好的顺序移动孵化盒在气候箱中的位置以减少箱内热梯度所造成的影响。
1.2 幼体运动表现和心率的测定
待有幼体开始出壳,增加检查孵化盒的频次,及时将新生幼体移出孵化盒,随即测定体质量。2个月后,大部分龟卵成功孵化。孵出幼体暂养于水深约3 cm的60 cm × 45 cm × 30 cm饲养箱(每个饲养箱内幼龟不超过5只),所有饲养箱放置于温度预先设为(30.0±0.5)℃的恒温室内。孵出幼体中选取无畸形的个体(每种龟每个养殖群20只)用于测定其功能表现(游泳和翻身反应)。所有测试在恒温室内进行。恒温室预先设置好的测试温度7个(16、20、24、28、32、36、40 ℃),测试温度顺序随机排列。一个测试温度结束后,幼龟均放回原来饲养箱,隔1 d之后再进行另一个温度。实验开始前,幼龟置于底部铺有湿纱布的饲养箱(每饲养箱幼龟不超过5只)内,在预先设置好测试温度的恒温室内适应2 h,以确保动物达到相应体温。测定游泳表现前,在长约1 m的带刻度直线泳道中注入与恒温室温度一致的水,泳道内水深约10 cm,将单个幼龟在泳道一端放入,一人用毛笔头轻触幼龟尾部驱使其向前游动,另一人手持一松下HDC-HS900数码摄像机拍摄幼龟在泳道内一个来回的游泳过程。泳道水温用点温计实时监控。若水温偏差较大时,注入适量热水(或冷水)搅匀以使水温变动不超过0.5 ℃。测定翻身反应时,将幼龟腹面朝上放在25 cm × 20 cm × 4 cm塑料盒的中央位置,同时用塑料盒正上方(约1.5 m)垂直向下放置的HDC-HS900数码摄像机拍摄动物的翻身过程。游泳和翻身表现拍摄结束后,所有视频片段用MGI VideoWave III分析软件(MGI Software Co.,Canada)读取幼龟在泳道内游过30 cm的最快游泳速度和成功翻身所需的时间间隔。翻身测试的幼龟若在10 min内未能成功翻身,则对应数据将不用于统计分析。
心率测定在上述相同温度下进行,个体则是从运动测试中随机选取的一半(每种龟每个养殖群10只)。测定开始前,用透气海绵固定幼龟以防止其活动,轻放于Buddy system红外胚胎心率监测仪(Avian Biotech International,USA),待显示心率稳定后读取相应数值(用每分钟心跳次数表示,Beat per minute,bpm)。
1.3 数据处理与统计分析
用SPSS 20.0统计软件包进行统计分析。参数统计分析前,用Kolmogorov-Smirnov检验和Levene检验分别检验数据的正态性和方差同质性。用两因子方差分析检测孵出幼体大小的养殖群及物种间的差异;巢式重复测量方差分析检测测试温度、物种及养殖群对游泳速度、翻身反应及心率的影响。用Table Curve 2D 5.01曲线拟合程序(Systat Software Inc.,USA)对不同测试温度的游泳速度、翻身时间倒数和心率的平均值(以引自Geng等[17]的临界高低温平均值作为游泳速度、翻身时间倒数和心率为0时的对应体温)以指数修正的高斯模型(Exponentially modified Gaussian model)构建热功能曲线,并从每条曲线中提出最大功能值和热表现宽度(实现最大功能表现80%水平以上的温度范围)。文中描述性统计值用平均值±标准误表示,显著性水平设置为α=0.05。
2 结果
测试用红耳滑龟孵出幼体体质量大于乌龟幼体(F1,76=206.93,P<0.001),而来自不同养殖群的幼体大小并没有显著差异(F1,76=3.05,P=0.085)(表1)。

表1 来自不同养殖群的乌龟和红耳滑龟孵出幼体的体质量Table 1 Mean body mass (Mean±SE)of Mauremys reevesii and Trachemys scripta elegans hatchlings from two different cohorts
幼龟的游泳速度、翻身时间和心率受测试温度、物种及养殖群差异的影响均显著(表2)。在16—40 ℃的测试温度范围内,两种龟幼体心率均随着温度的升高而显著增加;但游泳速度通常在32 ℃或36 ℃时达到最大(除来自海宁养殖群的乌龟游速在40 ℃达到最大)、翻身时间达到最短。总体上讲,红耳滑龟幼体游泳速度、心率大于乌龟幼体,而翻身时间则长于乌龟幼体。来自海口养殖群的两种龟幼体运动表现好于海宁养殖群幼体,心率也较快(图1)。

图1 不同测试温度下乌龟和红耳滑龟孵出幼体的运动表现(游泳速度和翻身时间)和心率Fig.1 Locomotor performances (i.e., swimming speed and righting time) and heart rate (Mean±SE) of Mauremys reevesii and Trachemys scripta elegans hatchlings
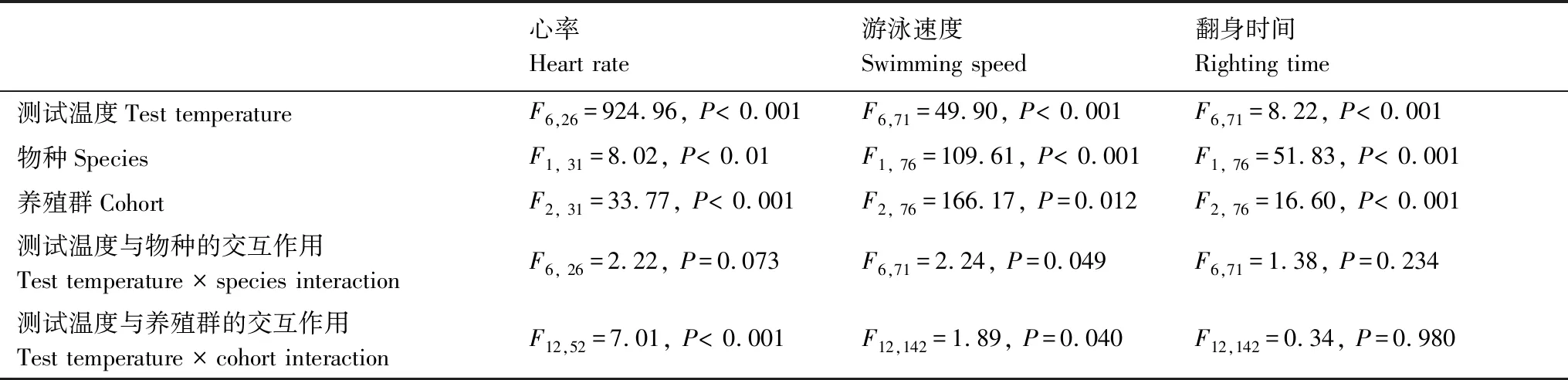
表2 不同测试温度下乌龟和红耳滑龟孵出幼体的运动表现(游泳速度和翻身时间)和心率的巢式重复测量方差分析结果Table 2 Results of nested ANOVAs with repeated measures on locomotor performances (i.e.,swimming speed and righting time) and heart rate of Mauremys reevesii and Trachemys scripta elegans hatchlings
利用Geng等[17]的临界高低温数据构建热功能曲线(图2)显示:来自海口和海宁养殖群的红耳滑龟幼体最大心率分别为128.8、107.8 bpm,最大游泳速度为18.8、16.1 cm/s,最短翻身时间为48.4 s和102.8 s;而乌龟幼体最大心率为107.8 bpm、102.0 bpm,最大游泳速度为12.4 cm/s、10.1 cm/s,最短翻身时间为26.2 s和51.9 s。来自不同养殖群的红耳滑龟幼体(海口10.1 ℃,海宁12.4 ℃)游泳速度的热表现宽度大于乌龟幼体(海口7.6 ℃,海宁8.9 ℃)相应值;而红耳滑龟幼体(海口10.8 ℃,海宁12.7 ℃)翻身时间的热表现宽度小于乌龟幼体(海口12.0 ℃,海宁14.2 ℃)相应值;红耳滑龟幼体(海口10.0 ℃,海宁7.8 ℃)心率的热表现宽度与乌龟幼体(海口9.4 ℃,海宁8.1 ℃)相应值差异不大。
3 讨论
与先前针对龟鳖类动物一些研究结果一致[15,18-19],温度显著影响乌龟、红耳滑龟幼体的运动和心率表现。在16—40 ℃测试温度范围内,两种龟幼体心率随温度升高逐渐加快;而运动能力(游泳速度和翻身反应)先随温度升高而逐渐加强,但温度过高时随之快速下降。所检测的运动和心率表现同时显示出不同物种间以及地理养殖群间的差异。特别是红耳滑龟幼体的游泳速度和心率显著快于乌龟幼体,可能反映了入侵龟在运动表现以及相关联的代谢水平上较土著龟显示出的一些竞争优势。
动物的运动能力(速度与耐力)在一定程度上可决定其在自然环境中活动的范围大小、与其它个体竞争取胜、以及长期存活的机率[20]。较好的运动能力有助于外来入侵性物种在进入新环境后能快速扩大其活动范围,并使其成功入侵这类生境成为可能[21-22]。与第一个预测相符,红耳滑龟幼体在不同测试温度获得的游泳速度实测值以及热功能曲线中提取的游泳速度最大值均显著大于乌龟幼体。类似结果也见于其它入侵-土著物种的对比研究。例如,与土著的亲缘关系较近物种相比,入侵澳大利亚的甘蔗蟾蜍(Rhinellamarina)可显示出较强的运动耐力[21];入侵法国的疣尾蜥虎(H.frenatus)显示出较快的运动速度[7];入侵智利的非洲爪蟾(Xenopuslaevis)显示出较快的翻身表现[22]。相反地,与预测不一致的是红耳滑龟幼体的翻身速度则慢于乌龟幼体。另一项与地中海拟水龟(M.leprosa)对比研究显示,入侵西班牙的红耳滑龟显示出较快的翻身速度[12]。这几种龟类对比研究存在不一致的结果可能反映了龟类翻身表现受形态差异的影响较大有关。扁平体型使龟鳖翻身的困难程度增加[23]。地中海拟水龟的体型比红耳滑龟更扁平(较小的背甲高:背甲宽比值),需较长的时间实现成功翻身与上述预测相吻合。同样,本研究中的红耳滑龟幼体体型与乌龟幼体相比则显得较扁平,那么红耳滑龟幼体需花较长时间实现翻身也是可预见的。这两项对比研究显示相反的结果表明以翻身表现作为入侵龟运动能力的潜在优势指标时应需考虑形态上的影响。
红耳滑龟幼体游泳速度的热表现宽度大于乌龟幼体,这与我们第二个预测相符,表明红耳滑龟比乌龟能在较宽的温度范围内较好地表达其运动水平,这也被认为是入侵种所具有的优势性特征[22,24-25]。同样,这种优势性特征在不同功能指标间存在差别。例如,红耳滑龟幼体翻身时间的热表现宽度反而小于乌龟幼体。虽然翻身表现常被用来作为评价龟类适合度,但该指标可能较适合用于种内或形态差别不大的种间比较研究。对体型差异较大但同属半水栖型龟类的红耳滑龟与乌龟运动表现对比研究时,游泳、爬行速度及耐力等运动指标比翻身表现可能更好些。入侵种运动表现的不同指标(如上述速度、热表现宽度)一致性地好于土著种。例如,入侵性非洲爪蟾翻身速度显著快于土著盖氏智利蟾(Calyptocephalellagayi),但其翻身时间的热表现宽度反而小于后者[22]。
对比研究也显示两种龟幼体所测量的运动表现以及其热依赖性在不同养殖群体间存在差异。来自海口养殖群的两种龟幼体的运动表现均好于海宁养殖群;海口养殖群个体游泳速度和翻身时间的热表现宽度均小于海宁养殖群个体。不同养殖群间的差异可能反映了动物在各自生活环境长期驯化的结果[24]。与纬度相对较高的海宁地区,纬度较低的海口地区年平均温度应较高,但温度变异较小。较高温度条件下驯化的动物通常比较低温度驯化的动物具有较好的运动表现;而经历较宽温度范围驯化可能会使动物在较大温度范围内表达相对较好的运动能力[8]。海口养殖群个体运动能力好于(但热表现宽度小于)海宁养殖群个体与上述预测是相符合的。此外,结果显示测试温度与养殖群交互作用对游泳速度具有显著的影响,表明游泳速度的热依赖性在不同养殖群中存在差异。红耳滑龟游泳速度热表现宽度两养殖群间的差值(2.3 ℃)大于乌龟相应值(1.3 ℃),可能反映了红耳滑龟游泳表现的较大可塑性,这与我们第三个预测相符。热生理及功能表现的较大可塑性被认为有助于易化入侵种的分布扩散[24,26]。入侵种一些生理表现(例如,甘蔗蟾蜍的低温耐受性和代谢率)的可塑性大于土著种已得到证实[24,27-28]。
心率是一个可反映动物代谢水平的重要生理指标[29]。与其它龟类(欧洲泽龟Emysorbicularis、北美拟鳄龟Chelydraserpentina)[30]研究结果类似,红耳滑龟和乌龟幼体心率随测试温度升高而显著增加。该结果也反映了高体温条件下动物表现出较高能量代谢水平。与运动表现的两个指标不同,在高测试温度下两种龟幼体心率并没有显示出下降的趋势,预测可能只有接近致死高温因心脏供血不足时才会出现快速下滑。心率在不同种间和养殖群体间也具有显著差异。通常情况下,动物心率有随着体型增加而下降的趋势。红耳滑龟幼体体质量大于乌龟幼体,心率反而显著高于乌龟幼体。该结果反映了红耳滑龟所具有的较高代谢水平,这可能与其在自然环境中入侵种较高活动水平与竞争能力有关联。虽然目前缺乏红耳滑龟与乌龟野外互动情况的数据,但另一项与地中海拟水龟的比较研究可反映出入侵性红耳滑龟与土著龟互动过程中展现出明显优势性。在与土著龟混养的环境下,红耳滑龟表现出更具侵略性、以及较强的争夺调温热资源和食物资源的行为表现[31-32]。此外,海口养殖群个体心率高于海宁养殖群个体,这可能因较低纬度地区相对较高温环境中长期驯化所导致的。有意思的是,一些研究显示短期的较高温驯化会导致龟类心率下降[30]。较高温下长短期驯化导致龟类心率变化趋势是否相反仍需在后续研究中加以确认。
