土壤酸胁迫对黑麦草生长生理及镉富集特征的影响
2021-10-11戴学斌许瑜兴马佳珠陈莉莉穆兴民
戴学斌,许瑜兴,邓 义,马佳珠,陈莉莉,穆兴民
(1.玉溪师范学院 地理与国土工程学院,云南 玉溪 653100;2.中国科学院 水利部 水土保持研究所,陕西 杨凌 712100)
土壤重金属镉(Cd)污染问题已经引起国内外的广泛关注。Cd是毒性非常强的一种重金属,易被作物吸收后通过食物链危害到人类健康。我国南方地区Cd污染程度比北方地区高[1],加之南方地区土壤酸化尤为严重[2],更加剧了土壤Cd污染[3]。因此,解决我国南方土壤酸化条件下Cd污染问题尤为迫切。植物修复技术是利用修复植物对土壤中的重金属进行固定,通过收获植物降低土壤中的重金属含量,从而达到修复目的[4]。与传统的物理修复和化学修复方法相比,植物修复以其环境友好及成本低等优势成为重金属污染治理研究领域的热点,其中修复植物的选择是植物修复最关注的内容[5-6]。目前,已有文献报道的Cd富集植物有宝山堇菜(Violabaoshanensis)[7]、商陆(Phytolaccaamericana)[8]、龙葵(Solanumnigrum)[9]、三叶鬼针草(Bidenspilosa)[10]、滇苦菜(Picrisdivaricata)[11]、东南景天(Sedumalfredii)[12]和忍冬(Lonicerajaponica)等[13],但这些植物因其生物量较小,或因环境适应性较差等因素,未被大面积地推广及应用,不能满足对土壤Cd污染治理的需求。且目前用于Cd污染修复的植物多是一年生植物,其地上部分在生长季结束后发生脱落,不能连年覆盖于地表,因此起不到很好的植物修复效果[14]。当前有关Cd富集植物的研究多集中在杂草[10]、蔬菜[15]和农作物[16],通过草本花卉进行土壤修复的研究刚刚兴起。草本花卉作为修复植物,在进行Cd污染土壤修复的同时,还可以增加绿化量以及美化环境。
多年生黑麦草(Loliumperenne)为禾本科黑麦草属多年生草本花卉植物,对Cd有很强的耐性和富集能力,其分布广、易栽培、生长速度快、根系发达、耐性强、可多次刈割并再生,因此在土壤Cd污染植物修复方面具有重要的研究及应用价值[17]。关于Cd对黑麦草的胁迫已有许多的文献报道[14,17-18],但目前对黑麦草在酸性土壤中的种植研究还相对较少,对黑麦草在酸胁迫下的适应策略尚不太清楚。因此,本研究是以Cd富集植物黑麦草为研究对象,通过盆栽试验研究不同强度酸胁迫对黑麦草种子萌发、植物生长生理及Cd富集特征的影响,旨在探讨黑麦草对土壤酸胁迫的耐性和对Cd污染土壤修复的可行性。为我国南方土壤酸化条件下Cd污染土壤的植物修复提供理论依据。
1 材料与方法
1.1 试验材料
供试植物为在南方酸性红壤地区分布广泛且生长良好的Cd富集植物多年生黑麦草,黑麦草种子购于玉溪花鸟市场。供试土壤采自玉溪地区未受Cd污染的0—20 cm表层混合土壤,除去石块树枝等杂质,将土样自然风干、磨细和过筛(5 mm),供盆栽试验使用(土壤pH值4.5,有机质9.55 g/kg,全氮1.09 g/kg,全磷0.40 g/kg,Cd背景值0.14 mg/kg)。试验过程中所用的试剂硫酸、氢氧化钠、氯化镉等均为分析纯。
1.2 试验设计
将制备好的土壤装入到塑料花盆(盆口直径32 cm×底径20 cm×高22 cm),每盆装土4 kg。设置4种酸胁迫水平:以溶液形式往供试土壤中添加硫酸和氢氧化钠,使土壤pH值为4.0,4.5,5.0,6.0,混匀后,静置10 d。Cd浓度设置100,0 mg/kg两个处理,方法是将一定量CdCl2溶液加入土壤中,混匀,在室内静置30 d后进行盆栽试验,期间添加纯净水4次,使土壤能够保持湿润。试验一共8个处理,每个处理3个重复。试验土壤基本理化性质见表1,Cd0和Cd100土壤中的有机质、全氮和全磷含量在土壤pH值为4.0时均较低;随着土壤pH值升高,土壤有机质和全氮含量呈现先升高后降低的趋势,且在土壤pH值5.0时最高。

表1 试验土壤基本理化性质
选取饱满的黑麦草种子,经浸种后进行均匀播种,每盆播种40粒,在温室内培养。在播种的第15天进行间苗,每盆留20株长势良好及大小一致的幼苗。黑麦草生长期间,根据盆中土壤的干湿状况进行浇水,使土壤含水量保持在田间持水量的80%,水中未检出Cd。黑麦草生长期间采用自然光照,昼夜温度为25℃/20℃,相对湿度66%。每7 d添加1次Hoagland营养液,每次每盆200 ml,同时注意防治病虫害,拔除盆中杂草。黑麦草在生长90 d时收获,取样测定相关的指标。
1.3 指标测定
(1)种子萌发指标的测定。每天观察并记录黑麦草种子的发芽情况,待种子萌发结束后,计算种子发芽势、发芽率和发芽指数,其计算公式具体如下所示:
发芽势=5 d内正常发芽种子数/供试种子数×100%
发芽率=10 d内正常发芽种子数/供试种子数×100%
发芽指数GI=∑(Gt/Dt)
式中:GI为发芽指数;Gt为第t日的发芽种子数;Dt为发芽天数。
(2)生长生理指标的测定。将植株保持完整取出后分为地上和根部两部分,用蒸馏水洗净沥干水分后测量记录株高和根长。叶绿素含量采用紫外分光光度法进行测定[17];根系活力采用氯化三苯基四氮唑(TTC)还原法进行测定[19];叶片和根系丙二醛含量采用硫代巴比妥酸比色法进行测定[19-20]。将黑麦草样品放入烘箱中,在105℃杀青15 min,在65℃烘干至恒重,用万分位电子天平称重,计算地上生物量、地下生物量和总生物量。根冠比通过地下生物量与地上生物量的比值计算。将采集的新鲜根样品,迅速投进5%戊二醛的固定液中,抽气使材料沉底,在4℃冰箱里固定24 h。固定的材料用双蒸水冲洗,经乙醇系列脱水,用环氧树脂包埋,用超薄切片机切片,切片用醋酸双氧铀和柠檬酸铅染色后,用透射电镜对黑麦草根细胞的细胞壁、细胞基质、线粒体进行观察及拍片[21-22]。
(3)Cd富集指标的测定。将烘干的黑麦草样品磨碎过尼龙筛(0.25 mm),采用HNO3-HClO4法消解;将风干土壤样品磨碎过0.15 mm筛,采用HNO3-HCl-H2O2消解。消解液中的Cd含量采用原子吸收分光光度法进行测定。黑麦草的Cd富集系数和转运系数根据如下公式计算:
黑麦草地上部富集系数=地上部Cd含量/土壤中Cd含量
黑麦草根部富集系数=根部Cd含量/土壤中Cd含量
黑麦草转运系数=地上部Cd含量/根部Cd含量
1.4 数据处理
使用R4.0.2对数据进行分析。采用双因素方差分析检验土壤酸胁迫、Cd浓度及二者交互作用对黑麦草种子萌发和生长生理指标的影响,采用Duncan多重比较检验法进行不同处理间的差异显著性检验。使用SigmaPlot 14.0作图。图表中的数据为平均值±标准误。
2 结果与分析
2.1 土壤酸胁迫对黑麦草种子萌发的影响
从表2可以看出,土壤酸胁迫及其与Cd浓度处理的交互作用对黑麦草种子的发芽势和发芽率有极显著影响(p<0.01),对种子发芽指数无显著影响(p>0.05)。由图1可知,当土壤pH值为4.0(Cd0)时,黑麦草种子的发芽势和发芽率明显受到抑制(p<0.05);随着土壤pH值升高,种子发芽势和发芽率表现先升高后降低的趋势。在Cd100土壤中,黑麦草种子的发芽势在pH值6.0时显著高于pH值4.0和pH值4.5(p<0.05);种子发芽率在pH值5.0时达89.17%,显著高于其余3种酸处理(p<0.05)(图1)。
2.2 土壤酸胁迫对黑麦草生长生理的影响
由表2可知,土壤酸胁迫对黑麦草的株高、根长和根冠比有极显著影响(p<0.01),土壤酸胁迫及其与Cd浓度处理的交互作用对黑麦草的地上生物量、地下生物量和总生物量存在极显著影响(p<0.01)。从图2可以看出,在Cd0土壤中,黑麦草的株高、根长、地上生物量、地下生物量和总生物量随着土壤pH值的升高呈先升高后降低的趋势,且在土壤pH值5.0时达到最大,显著高于pH值4.0(p<0.05)。土壤pH值为5.0(Cd0)时黑麦草的株高、根长、地上生物量、地下生物量和总生物量分别为pH值4.0(Cd0)的1.76倍、2.20倍、1.91倍、2.20倍和2.04倍。在Cd100土壤中,黑麦草的株高、根长、地上生物量、地下生物量和总生物量在pH值4.0时显著低于其余3种酸处理(p<0.05)(图2)。表明较高浓度的土壤酸胁迫抑制黑麦草生长。

图2 土壤酸胁迫对黑麦草生长的影响
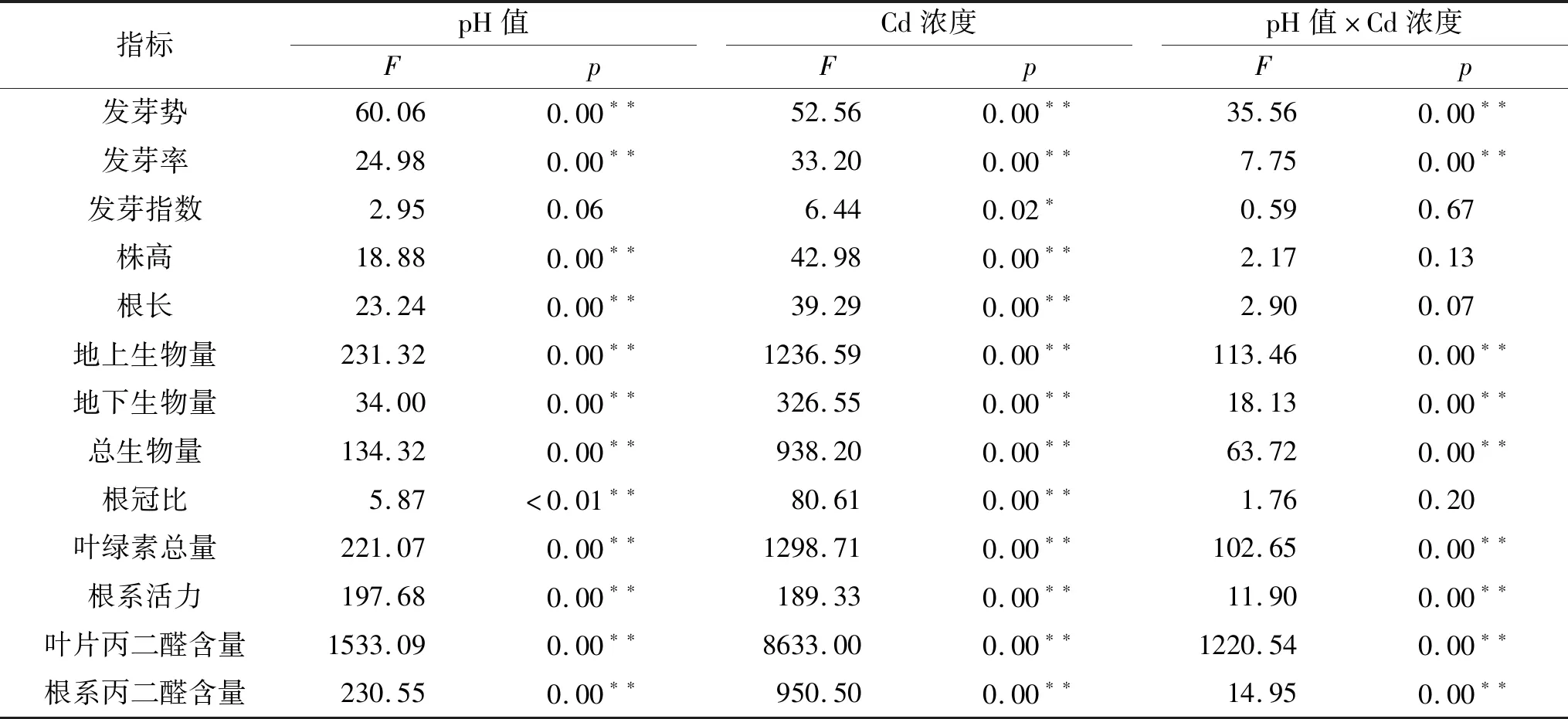
表2 土壤酸胁迫、Cd浓度及交互作用对黑麦草种子萌发和生长生理的影响
注:Cd0和Cd100分别表示Cd浓度为0,100mg/kg;不同字母表示处理间差异显著(p<0.05),下同。
土壤酸胁迫及其与Cd浓度处理的交互作用对黑麦草的叶绿素总量、根系活力、叶片丙二醛含量和根系丙二醛含量有极显著影响(p<0.01)(表2)。由图3可知,当土壤pH值为4.0时,在Cd0和Cd100土壤中生长的黑麦草的叶绿素总量和根系活力明显受到抑制(p<0.05);随着土壤pH值升高,叶绿素总量和根系活力显著增加(p<0.05)。相反的,黑麦草叶片和根系的丙二醛含量随着土壤pH值升高而显著降低(p<0.05)(图3)。
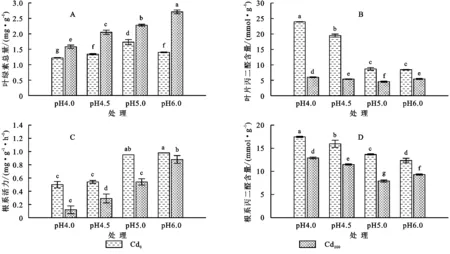
图3 土壤酸胁迫对黑麦草生理特性的影响
土壤酸胁迫破坏黑麦草的根细胞超微结构。透射电镜下观察黑麦草根细胞,当土壤pH值为6.0(Cd0)时,细胞壁结构紧密,厚度均匀,细胞膜完整,细胞基质呈凝胶态,线粒体为圆形或椭圆形,线粒体双层膜结构明显,线粒体内嵴结构清晰(图4A—B);当土壤pH值降低至4.0(Cd0)时,细胞壁粗细不均,部分细胞壁溶解断裂,细胞壁内侧出现絮状物,细胞膜结构破坏明显,细胞基质由凝胶态向分散碎块变化,线粒体解体为絮状物难以辨别(图4C—D),表明高浓度的酸胁迫会造成黑麦草根细胞壁和线粒体结构的破坏。
2.3 土壤酸胁迫对黑麦草Cd富集的影响
土壤酸胁迫对黑麦草Cd富集的影响见图5。在Cd100土壤中,不同土壤酸胁迫下的黑麦草地上部Cd含量为93.74~208.76 mg/kg,根部Cd含量为369.95~827.59 mg/kg,地上部富集系数为1.20~2.41,根部富集系数为3.78~9.55,可见植物体内不同部位Cd含量和对Cd的富集系数不同。黑麦草地上部和根部对Cd的富集系数均大于1,且根部对Cd的富集系数高于地上部。黑麦草地上部Cd含量、根部Cd含量、地上部富集系数和根部富集系数均随着土壤pH值升高表现出先升高后降低,其在pH值4.5和pH值5.0时显著高于pH值4.0和pH值6.0(p<0.05)。黑麦草的转运系数为0.19~0.32,随着土壤pH值升高显著降低(p<0.05)。

图5 土壤酸胁迫对黑麦草Cd富集的影响
3 讨 论
发芽势反映了种子出苗速度的快慢,发芽率能够反映种子生命力的强弱[20]。本研究中黑麦草种子萌发随土壤pH值的变化特征表明,较高浓度的土壤酸胁迫(pH值4.0)对黑麦草种子的发芽势和发芽率具有明显抑制作用,而较低浓度的土壤酸胁迫(pH值4.5~6.0)对种子发芽势和发芽率具有一定的促进作用。即黑麦草种子在弱酸性土壤条件下能够正常萌发,具有一定的耐酸性,但是在强酸性土壤条件下,种子的萌发受到了显著抑制,这与魏祯祯[23]、何影[24]等的研究结果相似。酸胁迫下黑麦草种子萌发受到抑制的原因可能是低浓度的酸胁迫可改善胚的生理活性,促进种子萌发;而高浓度酸胁迫降低种子细胞内保护酶活性,抑制种子内储藏物质分解与呼吸代谢,从而影响种子萌发所需的物质与能量,对胚根及胚芽等造成损伤[25]。

注:Cw,Cm和M分别表示细胞壁、细胞基质和线粒体。
植物生长对土壤酸碱度有一定要求,酸胁迫对植物生长的影响可通过株高、根长、生物量和根冠比等形态指标直观地表现出来。在本研究中,黑麦草的株高、根长、地上生物量、地下生物量和总生物量在较高浓度土壤酸胁迫(pH值4.0)下受到明显的抑制作用。根系是植物吸收养分的主要器官,黑麦草根系的生长抑制和生物量的降低意味着该植物在较高浓度土壤酸胁迫下吸收养分的能力下降,进而对整个植株的生长造成影响。王一鸣等[19]通过对土壤酸胁迫下景天三七生长进行研究发现,植物的株高和根长随土壤酸化程度的加剧受到明显的抑制,这与本研究结果一致。而项敬银等[3]对秃瓣杜英生长的研究表明,酸胁迫显著促进了秃瓣杜英总生物量的积累。试验结果表现差异的原因可能是由于不同植物对酸胁迫敏感性的不同。生物量的大小是判定植物耐性的指标之一[26]。本研究发现,黑麦草的根长、地下生物量和根冠比在较低浓度土壤酸胁迫(pH值4.5~6.0)下无显著差异,且生长良好,说明该植物的根系对土壤酸胁迫具有较强耐性。蒙程等[2]研究了土壤酸胁迫对紫花苜蓿生长的影响,结果表明较低浓度的酸胁迫对植物的株高和地上生物量影响不显著,与本研究结果相似。
土壤酸化严重时会使叶绿素含量减少,光合作用减弱,影响植物正常生长。在本研究中,黑麦草的叶绿素总量在较高浓度土壤酸胁迫(pH值4.0)时最低,随着土壤pH值的升高而显著增加(p<0.05)。陶巧静等[27]研究发现,酸胁迫对植物叶绿素的合成表现出高浓度抑制而低浓度促进的影响,与本研究结果一致。
但也有研究表明,在植物能够正常生长的情况下,较高浓度的酸胁迫对植物叶绿素含量具有一定的促进作用[28]。胁迫对植物叶绿素的影响表现在使叶绿素的合成受阻和降解加速两方面[29]。根系活力的高低直接影响植物的生长。黑麦草的根系活力随土壤pH值升高的变化趋势与叶绿素总量的相一致。黑麦草根系活力在较低浓度土壤酸胁迫下较强,这与王一鸣等[19]对景天三七根系活力的研究结果相同。丙二醛含量的多少可反映植物体细胞膜受损情况[26]。黑麦草叶片和根系的丙二醛含量在土壤pH值4.0时最高,随着土壤pH值升高呈显著降低,在pH值5.0时达到最低。说明较高浓度的土壤酸胁迫会使植物体内产生大量丙二醛,细胞膜受损严重,影响植物正常生长。陈雪娇等[30]的研究表明,类芦丙二醛含量在土壤pH值3.5时显著增加,与本研究结果相似。酸胁迫对黑麦草叶绿素总量、根系活力、叶片和根系丙二醛含量的研究表明,该植物对土壤酸胁迫的耐性较强。
膜体系的损伤被认为是造成植物体受损害的根本机制[31]。与较低浓度的土壤酸胁迫(pH值6.0)相比,较高浓度的土壤酸胁迫(pH值4.0)对黑麦草的根细胞超微结构具有破坏作用。当土壤pH值降低至4.0时,细胞壁粗细不均,部分细胞壁发生溶解断裂,细胞壁内侧出现絮状物,细胞膜结构破坏明显,细胞基质由凝胶态向分散碎块变化,线粒体解体为絮状物难以辨别,说明高浓度的酸胁迫对黑麦草根细胞壁和线粒体结构造成损伤,这种损伤是不可逆的,可使植物根系活力和生物量下降,最终将导致植物死亡。钱莲文等[22]的研究也发现,常绿杨在高浓度铝胁迫下的根冠细胞壁、细胞基质、淀粉质体和淀粉粒明显受损,与本研究结果相似。王一鸣等[32]研究发现,景天三七在轻度(pH值5.5)和中度(pH值4.6)土壤酸化下叶绿体超微结构完整,在重度(pH值3.4)土壤酸化下叶绿体的结构受到严重破坏。
植物对Cd富集能力的大小是进行Cd污染土壤修复时筛选植物的重要指标之一,植物对Cd富集能力的大小受到土壤pH值的影响。土壤酸化严重时会影响着植物根系的生长,而根系形态结构的变化反过来会影响根系对Cd吸收,从而影响Cd在植物体内的转运和积累。本研究中,不同土壤酸胁迫下的黑麦草地上部Cd含量高于100 mg/kg(仅pH值6.0除外),根部Cd含量高达369.95 mg/kg以上,可见该植物在土壤酸胁迫下仍对Cd表现出较强的积累能力。项敬银等[3]研究表明,秃瓣杜英在酸胁迫下对Cd也具有较强的积累能力。富集系数被作为评价植物对Cd富集能力的重要指标,富集系数越高,说明植物对Cd的吸收积累能力就越强,越是有利于植物对Cd污染土壤的修复[33-34]。本研究结果显示,不同土壤酸胁迫下的黑麦草地上部和根部对Cd的富集系数都大于1,表明该植物在土壤酸胁迫下仍对Cd具有较强的富集能力。黑麦草地上部Cd含量、根部Cd含量、地上部富集系数和根部富集系数在不同土壤酸胁迫下差异显著,其在pH值4.5和pH值5.0时要显著高于pH值4.0和pH值6.0(p<0.05),说明土壤pH值在适度弱酸性条件下更有利于黑麦草对Cd的富集。黑麦草根部的Cd含量和富集系数明显高于地上部,导致转运系数小于1,这也表明该植物吸收的Cd主要富集在根部,其向植物地上部转运Cd的能力较弱,与其他研究者的结果相同[3,14,17,21,34-35]。
通过以上分析可发现,植物对酸胁迫的耐受性存在一定的范围。研究表明,景天三七能够适应土壤pH值4.2~6.2的土壤环境[19],类芦可在土壤pH值3.5~5.6条件下正常生长[30],刨花楠在土壤pH值2.5时仍然能够生长[29],可见不同植物生长对酸胁迫的耐受范围不同。通常,在强酸性土壤条件下仍能够存活的植物属于耐酸性植物。黑麦草在土壤pH值4.5~6.0时种子萌发、植物生长生理及Cd富集能力的各项指标都表现出该植物对土壤酸胁迫具有较强的耐性,在酸性土壤条件下的土壤Cd污染植物修复中具有较大利用潜力。
4 结 论
(1)黑麦草在土壤pH值4.5~6.0时生长和生理状况良好,但当土壤pH值降低至4.0时黑麦草种子发芽势和发芽率减小,植物株高、根长、生物量、叶绿素总量和根系活力都降低,根细胞超微结构受损,Cd富集能力受到明显的抑制。
(2)黑麦草在不同浓度土壤酸胁迫下对Cd的富集能力存在明显差异,从黑麦草地上部与根部的Cd含量和富集系数来看,土壤pH值在适度弱酸性(pH值4.5和pH值5.0)条件下更有利于黑麦草对Cd的富集。黑麦草根部对Cd的富集能力大于地上部。
