‘凤丹’种子发育及其营养物质含量和相关酶活性的动态变化
2021-10-10邹雨婷朱铭玮李永荣翟金庭李淑娴
邹雨婷,朱铭玮, 李永荣, 翟金庭,李淑娴*
(1. 南京林业大学林学院,南方现代林业协同创新中心,江苏 南京 210037;2. 南京绿宙薄壳山核桃科技有限公司,江苏 南京 210014;3.盐城林场,江苏 盐城 224136)
‘凤丹’(Paeoniaostii‘Feng Dan’)曾被称为“铁杆庄稼”[1],具有结实率高、适应能力强和可抵御风蚀等特点[2],其种子还具有含油率高、油品质好的特点。牡丹籽油中不饱和脂肪酸质量分数高达92.60%[3],富含人体必需脂肪酸——亚油酸和α-亚麻酸[4];不仅适于食用,还是一种重要的工业原材料[5]。此外,‘凤丹’不仅可作为经济作物进行种植,还可应用于园林造景[6]。因此,‘凤丹’是一种中国所特有的集生态、经济和营养价值为一体的优良木本油料作物。目前,不少学者主要在牡丹(P.suffruticosa)籽油的提纯方法、油脂成分分析、籽油抗氧化活性、籽粒储藏等方面开展了系列研究[7-10],在‘凤丹’的育种、栽植管理和种子休眠等方面研究也进展迅速[11-13],对‘凤丹’种子发育方面的研究主要集中在脂肪酸积累方面,但缺乏主要营养物质间转化关系的相关研究[14-16]。陈虹等[17]在核桃(Juglansregia)种子发育的研究中发现油脂的累积与蛋白质含量的增加呈正相关关系,可能会存在能源竞争的现象,但油茶(Camelliaoleifera)[18]种子中油脂与蛋白质的积累却呈现负相关关系。由此可以看出,不同植物中,油脂积累与蛋白质合成也存在不同的关系。
本研究以油用牡丹中的‘凤丹’为材料,在观察其果荚、种子发育过程中形态变化的同时,研究此过程中主要营养物质及其相关酶活性的变化,初步探究油脂合成与碳水化合物之间的关系,进一步揭示油脂累积的规律,以期找到影响‘凤丹’种子油脂合成的关键时期,为‘凤丹’的科学栽培及籽油的开发利用提供理论依据。
1 材料与方法
1.1 试验材料与设计
试验所用果荚及种子采自南京绿宙油用牡丹研究所(118°34′~119°03′E,32°11′~32°27′N)6年生‘凤丹’结实母树,于盛花期选取生长正常、株型一致(每株高度基本一致且均含5个分枝)、花朵大小一致且同天开放的植株挂牌,做好标记。‘凤丹’通常在4月初开始开花,果期在4月下旬至8月上旬,花后0~45 d为授粉成功、胚珠形成阶段[19]。本研究主要在‘凤丹’蓇葖果及种子进入快速生长期即花后45、65、85、95、105和115 d后进行采样,随机选取一定数量的新鲜蓇葖果和种子用于形态结构的观察、测定和含水率测定。其余种子密封于自封塑料袋中,贮藏在-80 ℃冰箱中以供营养物质含量以及相关酶活性测定,其中,除粗脂肪含量测定以冷冻干燥后的种子为材料外,其余均采用贮藏于-80 ℃冰箱中的新鲜蓇葖果和种子为材料。
1.2 指标测定
1.2.1 发育过程中蓇葖果形态观察
随机选取各发育阶段中具典型特征的完整聚合蓇葖果,观察其颜色的变化。
1.2.2 种子内部结构变化观察
随机选取各发育阶段的种子10粒,纵向切开,用OLYMPUS SZX16体视显微镜(奥林帕斯公司,日本)对其内部结构进行形态观察。
1.2.3 蓇葖果和种子横、纵径测定
于每个发育阶段随机取5个新鲜采集的聚合蓇葖果,生物学重复3次,分别用游标卡尺测量单个蓇葖果的横径及纵径,精确到0.01 mm。剥开果荚,得到种子,随机取30粒种子,生物学重复3次,用游标卡尺分别测量种子的横径及纵径,精确到0.01 mm。
1.2.4 含水率测定
随机选取每个发育阶段的种子20粒,重复3次,将其切成小于2 mm的小块,称其鲜质量,精确至0.001 g。将切成小块的种子置于-80 ℃冰箱中预冷2 h后取出,再将其置于初始温度为-76 ℃,压力为0.001 MPa的Alpha 1-4 LOC-1冷冻干燥机(Martin Christ,德国)中干燥直至样品达到质量恒定。最后将样品放置在装有硅胶的干燥器中。待样品恢复至室温后再次称质量,精确到0.001 g,以确定其干质量。按下式计算每个发育阶段种子的含水率。

1.2.5 营养物质含量测定
采用索氏提取法[20]测定粗脂肪含量,具体为:将各发育阶段干燥后的样品研磨成粉末,称取1.0 g粉末(M0),用脱脂滤纸(已于105 ℃下烘至质量恒定)包裹后称质量,记为M1(精确到0.001 g)。用石油醚(30~60 ℃)对各发育阶段的种子样品采用索式抽提法提取籽油。抽提完成后,待石油醚完全挥发,再次称质量,记为M2(精确到0.001 g),生物学重复3次。用相对于1.0 g初始干质量的质量(M0)损失百分数表示样品的粗脂肪质量分数,计算公式如下:
可溶性糖含量测定采用蒽酮比色法[21],粗淀粉含量测定采用高氯酸水解-蒽酮比色法[21],可溶性蛋白含量测定采用考马斯亮蓝比色法[22]。以鲜质量为单位计算各指标的含量。
1.2.6 营养物质相关酶活性测定
分别利用乙酰辅酶A羧化酶(ACCase)分光光度法试剂盒(苏州科铭生物技术有限公司,中国)和植物磷脂酸磷酸酯酶(PPase)ELISA 试剂盒(苏州科铭生物技术有限公司,中国)测定不同发育阶段种子中油脂合成ACCase和PPase相关酶的活性。采用3,5-二硝基水杨酸比色法[23]和福林-酚比色法[24]分别测定淀粉水解相关酶-总淀粉酶和蛋白质分解相关酶-总蛋白酶的活性,以鲜质量为单位计算各酶的活性。
1.3 数据分析
运用Excel 2010进行数据统计及图表制作,同时利用IBM SPSS Statistics 22.0进行单因素方差分析(ANOVA)及Duncan多重比较。
2 结果与分析
2.1 ‘凤丹’发育过程中形态的变化
2.1.1 蓇葖果及种子发育过程中外观形态及大小的变化
花后45~85 d时,‘凤丹’蓇葖果果皮由青绿色逐渐转变为黄绿色,也称绿熟期。随着蓇葖果的不断成熟,果皮由黄绿色逐渐转变为黄褐色(褐熟期)。除此之外,花后95 d时,少量果荚尖端沿着腹缝线开始开裂,直至花后115 d时,蓇葖果完全成熟,果荚尖端呈现灰褐色,且向下弯曲,并沿着腹缝线向基部开裂,露出黑褐色的种子。
每个发育阶段的蓇葖果及种子横、纵径的变化如表1所示。花后45~85 d期间(绿熟期),蓇葖果进入快速膨大期,横径和纵径快速增长,分别显著增长了12.7%和19.9%;而花后95~115 d期间(褐熟期),蓇葖果横径间的变化差异不显著,但纵径却呈减小趋势,花后105、115 d时的纵径相同,两个阶段均较花后95 d时显著减少了3.1%。

表1 ‘凤丹’蓇葖果及种子发育过程中的横、纵径变化Table 1 Changes of horizontal and vertical diameters during the development of Paeonia ostii ‘Feng Dan’ follicles and seeds
种子的横、纵径变化趋势与蓇葖果大致相同,于花后95 d时,分别达到此次测定最大值8.4 mm和11.1 mm,较45 d时分别显著增加了33.3%和32.1%。此后横径变化不显著,而花后115 d时的纵径较95 d时显著减少了10.9%。
2.1.2 ‘凤丹’种子发育过程中内部结构观察
花后45 d(图1A)的‘凤丹’种子,外种皮较薄,呈淡黄色,与珠被组织较易剥离;种子内部主要为珠被组织和胚乳组织。其中与种皮相邻的珠被组织较幼嫩,呈半液态;中央的胚乳组织则为透明状的液体,汁液丰富(图1A中蓝色圈内所示)。此时,珠孔端的胚仅为极小的一个点。花后65~85 d(图1B、1C)时,外种皮变厚,种皮、种仁变为乳白色;珠被组织逐渐变薄;点状胚开始初具雏形;合点端出现了一小块黑色区域(图1C黑色箭头所示);花后95~115 d(图1D、1E、1F)时,种皮逐渐向黑棕色转变,珠被组织消失,胚乳逐渐变硬,并在中央处出现裂缝;胚根极短,子叶呈卵圆形,位于胚乳中央细长线条处。

A-F均为‘凤丹’种子的纵切面,观测时间分别为花后45、65、85、95、105、115 d。 A-F are the longitudinal anatomies of Paeonia ostii seeds, and the observation time is 45, 65, 85, 95, 105, 115 days after flowering. I. 珠被组织 integument;En. 胚乳endosperm;Em. 胚 embryo;T. 种皮 testa;Ch. 合点chalaza。黑色箭头表示合点端出现了一小块黑色区域,蓝色圈出的部分表示胚乳部分。The black arrow indicates a black area at the chalaza and the part circled in blue represents the endosperm.图1 ‘凤丹’种子发育过程中的体视显微镜观察Fig.1 Stereoscopic microscope images of Paeonia ostii ‘Feng Dan’ seeds development
2.2 ‘凤丹’种子发育过程中含水率的变化
‘凤丹’种子的含水率在整个发育阶段处于持续失水趋势(图2),花后95 d时,含水率较45 d时显著下降了27.8%;花后95~115 d期间,含水率下降速度减缓,花后115 d时的含水率较95 d时仅下降了4.9%。但即使在完全成熟(花后115 d)时,‘凤丹’种子中的含水率仍然较高,为41.6%。
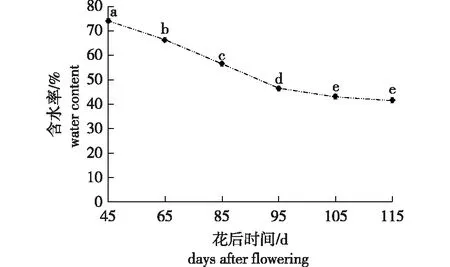
图2 ‘凤丹’种子发育过程中含水率的变化Fig.2 Changes of water content during Paeonia ostii Feng Dan’ seeds development
2.3 种子发育过程中营养物质含量的动态变化
不同发育阶段‘凤丹’种子中可溶性糖、粗淀粉、可溶性蛋白和粗脂肪含量的变化规律不尽相同(图3)。种子发育初期(花后45~85 d)可溶性糖含量快速下降,并于花后85 d达到本次测定的最小值,较45 d时显著下降了44.5%;发育中期(花后85~95 d)可溶性糖快速积累,随后再次下降并趋于稳定。粗淀粉含量变化趋势与可溶性糖大致相同,也是在花后45~85 d时呈下降趋势,花后85 d时的粗淀粉含量较45 d时显著下降了73.1%;种子发育中后期(花后85~105 d),粗淀粉含量呈快速上升趋势,花后105 d时的含量较85 d显著增加了187.2%,发育末期(花后105~115 d)时其含量达至稳定状态。可溶性蛋白含量的变化趋势和前两种物质有所不同,花后45~85 d时,其含量显著增加,花后85 d时较45 d大幅增加了294.1%;而在发育中期,可溶性蛋白含量快速下降;花后105~115 d时,可溶性蛋白含量波动较小。

图3 ‘凤丹’种子发育过程中营养物质的动态变化Fig.3 The dynamic changes of nutrients during Paeonia ostii ‘Feng Dan’ seeds development
粗脂肪质量分数在整个种子发育阶段呈不断上升趋势,变化趋势大概分为两个阶段:花后45~85 d,种子含油率迅速增加,从本次测定最低值13.8%显著上升到30.1%;随后,含油率增加趋势变缓,花后105 d时的含油率较85 d仅上升了1.3%;花后105~115 d期间,种子含油率的增加速度又略有上升,最终于花后115 d时,达到本次测定最大值33.1%。
2.4 种子发育过程中营养物质相关酶活性的变化
总淀粉酶和总蛋白酶分别具有水解淀粉和蛋白的作用。‘凤丹’种子发育过程中,这两种酶的活性处于不断变化中(图4A)。花后45~65 d即种子发育初期,总淀粉酶活性显著上升,从本次测定最低值53.2 μg/(mg· min)上升至196.4 μg/(mg· min),显著增加了269.2%;花后65~85 d时总淀粉酶活性略有上升,但变化不显著;种子发育的中后期(85~105 d),总淀粉酶活性呈显著下降的趋势,花后105~115 d期间,其活性差异不显著。
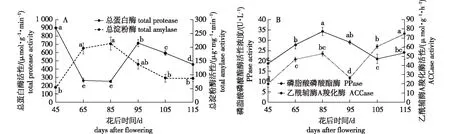
图4 ‘凤丹’种子发育过程中营养物质相关酶活性的变化Fig.4 Changes of nutrient-related enzymes activities during Paeonia ostii ‘Feng Dan’ seeds development
种子内总蛋白酶活性的变化趋势与淀粉酶的变化趋势大致相反(图4A)。花后45~65 d期间,种子总蛋白酶活性呈现下降趋势,花后65 d时的总蛋白酶活性较45 d时显著下降了70.4%;花后65~85 d期间,酶活性差异不显著;花后85~95 d时,总蛋白酶活性则又重新呈显著上升的趋势,花后95 d时酶的活性较85 d显著增加了185.3%;随后(花后95~115 d)酶活性呈线性下降趋势,且各时间点间差异显著,花后115 d时的总蛋白酶活性较95 d下降了36.1%。
‘凤丹’种子发育过程中ACCase和PPase活性的变化趋势大致相似(图4B)。花后45~85 d期间,ACCase和PPase的活性均呈上升趋势,85 d时酶活性较45 d分别显著上升了164.0%和82.4%;花后85~95 d,ACCase活性显著下降,从52.8 μmol/(g·h)下降至26.3 μmol/(g·h);PPase在85~105 d期间显著下降,活性由34.3 U/L显著下降至21.0 U/L;后期这两个酶的活性再次上升,特别是ACCase,115 d时活性较95 d上升了183.7%;PPase活性则在种子发育末期(花后105~115 d)略有上升,但差异不显著。
2.5 种子发育过程中营养物质含量及其酶活性的相关性分析
‘凤丹’种子发育过程中营养物质含量及其相关酶活性之间存在一定的相关性(表2)。其中,可溶性糖与粗淀粉含量呈显著正相关(P< 0.05),而可溶性蛋白与粗淀粉含量呈显著负相关关系(P< 0.05),与可溶性糖含量之间则呈极显著负相关关系(P< 0.01),与粗脂肪含量之间也呈正相关关系且达到极显著程度(P< 0.01);总淀粉酶活性与总蛋白酶活性之间呈显著负相关关系(P< 0.05),可溶性糖含量与总蛋白酶活性之间存在显著正相关关系(P< 0.05);磷脂酸磷酸酯酶活性与粗淀粉之间存在极显著的负相关关系(P< 0.01),与总淀粉酶活性则呈现极显著正相关关系(P< 0.01)。据统计分析,粗脂肪与含水率之间也呈负相关关系且达到极显著程度(P< 0.01),相关系数为-0.93。

表2 ‘凤丹’种子发育过程中营养物质含量及其酶活性的相关性分析Table 2 Correlation analysis of nutrients content and nutrient-related enzymes activities during Paeonia ostii‘Feng Dan’ seed development
3 讨 论
3.1 ‘凤丹’种子生长发育特性与栽培管理的关系
‘凤丹’种子营养物质的积累、转化与植株的光合作用及其栽培管理存在着密不可分的关系。花后45~65 d时,通过体视显微镜观察到,半液态化珠被组织的面积不断缩小,胚乳组织则由液态逐渐转变为固态,并且面积在不断增加,这与董兆磊[19]观察到的‘凤丹’种子胚乳的发育以及紫斑牡丹(P.rockii)[25]胚乳发育观察到的结果基本相同,说明胚乳从珠被等母体细胞中吸收的可溶性糖和粗淀粉等营养物质主要应用于自身形态建成,此时期也是种子胚乳固态化的重要时期。花后45~65 d,蓇葖果和种子的横、纵径处于大幅增长状态,是蓇葖果和种子的膨大期。此时,‘凤丹’植株不仅对水分的需求量较大,还对土壤中磷元素需求较高,应尽量保持水肥供应,有助于种子中光合产物的运输以及碳水化合物的代谢,保证种子的充分发育,这也从侧面反映了‘凤丹’生产中多在花后半个月左右开展施用磷肥管理的必要性和科学性。
花后65~85 d时,种子内的点状胚逐渐明显,珠被组织继续退化直至消失,胚乳逐渐硬质化。此阶段中,蓇葖果和种子的横、纵径仍在不断增加,种子内的可溶性糖和粗淀粉含量下降显著,而油脂和可溶性蛋白处于显著上升阶段,此时需适量施用氮肥促进植株内叶绿素的形成,从而促进光合作用的进行,以达到“以氮增碳”的作用。与此同时,适当减少磷肥的施用、增施钾肥,可提高糖类的积累、转化及运输。此时植株对水分要求相对敏感,以浅浇灌为最佳,为蓇葖果的持续膨大以及种子内部构建提供正常的水分供给。
花后85~115 d时,随着极短的胚根以及卵圆形子叶的不断发育完全,胚乳与种皮的接触愈加紧密,中央位置也逐渐开始形成一道长条形的空隙,其作用主要是便于种子萌发时子叶的伸展[26],种子内部结构逐步构建完成。在此阶段中,蓇葖果尖端逐渐褐化、开裂。蓇葖果和种子的横、纵径基本保持稳定甚至出现下降趋势,其中蓇葖果纵径在花后85 d时就开始稍有下降,花后105 d时已较最大值下降显著,明显早于其他测定指标,而造成这一变化的原因可能是,随着蓇葖果的成熟以及脱水现象的产生,使得果皮皱缩,导致蓇葖果尖端向下弯曲。因此,此阶段可以适当减少水分供给,使土壤处于较干燥状态,与此同时,花后85~95 d时,可溶性糖和粗淀粉含量显著上升,可再次追施少量磷肥,促进糖分和淀粉的代谢,为油脂和蛋白的积累提供充足的能源,利于提高种子的品质。
3.2 ‘凤丹’种子发育过程中营养物质含量间的转化
3.2.1 ‘凤丹’种子发育过程中主要营养物质含量与其相关酶活性的关系
种子发育初期,光合作用产生的物质以可溶性糖和粗淀粉等碳水化合物形式储藏在种子中。随着种子的不断发育,碳水化合物进行代谢及转化,种子中的蛋白和油脂不断积累,而淀粉、蛋白质和粗脂肪在代谢过程中都必须有酶的参与[27]。其中,总淀粉酶主要是将淀粉水解成葡萄糖等物质,为其他物质的合成提供原料;当总淀粉酶活性增大时,将更多的粗淀粉水解,从而使粗淀粉的含量呈下降趋势;反之,当总淀粉酶活性降低时,粗淀粉含量则趋于上升变化趋势。同样,总蛋白酶的主要作用是将蛋白质分解为可溶性含氮化合物,为其他物质的合成提供氮源[28-29]。PPase和ACCase则为油脂合成中的主要调节酶,其活性高低可能影响油脂的合成[30]。本研究发现,粗淀粉与可溶性蛋白含量的变化呈显著负相关,并且总蛋白酶也与总淀粉酶活性呈现显著负相关关系,这与前人研究基本一致[31]。但与向日葵(Helianthusannuus)、拟南芥(Arabidopsisthaliana)、油菜(Brassicanapus)和芝麻(Sesamumindicum)等大多数油料种子不同的是,它们种子油脂的快速积累主要集中在种子发育后期[32],而‘凤丹’种子油脂积累的旺盛时期主要集中在发育前期(45~85 d)。这一阶段种子中的ACCase和PPase酶活性不断上升,发育后期,这两个酶的活性迅速下降,含油率的增长也变得较为缓慢,这与山桐子(Idesiapolycarpa)种子油脂的积累规律[33]较为相似。
虽然ACCase和PPase酶活性的变化趋势以及花后85 d和花后115 d两次峰值的结果与油脂合成速率基本一致。但油脂的合成过程是多种生理机制共同作用的结果,除ACCase和PPase两种酶,还存在其他多种酶及营养物质的作用,他们共同对油脂的合成进行调节。本研究只是初步分析了主要贮藏物与其相关酶活性的关系,更多调节机制以及作用机理还需要进一步探究。
3.2.2 ‘凤丹’种子发育过程中油脂合成与碳水化合物的关系
种子发育过程中,油脂及蛋白的合成原料归根结底来源于糖类物质的分解与转化[34]。King等[35]也发现,种子发育初期所产生的贮藏物质大多来源于光合作用形成的糖类物质,这些物质主要以蔗糖的形式进入种子,然后分解为己糖。己糖磷酸化后,碳的分配就开始了[36],己糖既可生成淀粉等物质,也可进入糖分解途径转化为丙酮酸、磷酸丙糖等,用于油脂以及蛋白质等物质的合成[35]。由于种子中的淀粉和油脂分别处于糖氧化途径的上游和下游,碳源分配将最终引起这两类贮藏物质间含量的变化[37]。Lin等[38]在拟南芥突变体中通过“向上调控”,提高淀粉合成速率,最终由于碳源向油脂合成途径的流动被“切断”,导致种子油含量降低。赵翠格[39]研究发现,文冠果(Xanthocerassorbifolium)种子在油脂积累初期,体内的淀粉含量不断增加,而可溶性糖的含量却不断减少。本研究发现‘凤丹’种子在油脂快速累积期(花后45~85 d),淀粉和可溶性糖含量均呈快速下降趋势,其原因可能是,油脂大量合成时主要是由可溶性糖提供原料,后期由于可溶性糖无法供应充足的原料,才导致淀粉大量分解,以满足油脂合成对原料的需求;花后85~105 d时,油脂累积基本趋于平缓,而淀粉的含量则显著增加,说明此时碳源主要分配至淀粉处,从而使淀粉积累;种子最后的成熟阶段(花后105~115 d),油脂累积速度又开始缓慢增加,可溶性糖和淀粉的含量也再次突然下降。结合此发育过程中,淀粉酶活性与油脂合成关键酶之一——PPase活性呈极显著正相关关系,在‘凤丹’种子发育过程中,淀粉与油脂之间可能存在着“碳源分配”问题,但此结论仍需要进一步验证。
本研究还发现,‘凤丹’种子发育过程中,可溶性蛋白含量与可溶性糖含量呈极显著负相关,与粗淀粉含量之间也呈显著负相关,虽然其与粗脂肪之间还存在极显著正相关关系,但在发育后期(花后85~115 d)时,可溶性蛋白的含量与粗脂肪含量的变化趋势相反。因此,蛋白是否也参与油脂合成的碳源竞争仍需进一步研究验证。
