茜草科二型花柱植物滇丁香传粉精确性研究
2021-10-09王顺雨游贤松胡德美姚仁秀汤晓辛王晓月
陈 燕,王顺雨,游贤松,胡德美,姚仁秀,汤晓辛,王晓月,*
1 贵州师范大学生命科学学院,贵阳 550025 2 贵州师范大学国家林业局西南喀斯特山地生物多样性保护重点实验室,贵阳 550025 3 贵州师范大学贵州省植物生理与发育调控重点实验室,贵阳 550025
Darwin在《同种植物的不同花型》中首次提出异型花柱(heterostyly)植物在于促进型间授粉,自此异型花柱植物受到生态学家和进化生物学家的广泛关注[1],关于异型花柱的进化及适应意义主要有三种假说:(1)促进型间传粉[2- 4],(2)通过减少型内花粉的落置以阻止花粉浪费[5],(3)避免同型花雌、雄蕊间的相互干扰[5- 7],其中最核心在于促进型间花粉传递,从而促进异交提高传粉精确性。异型花柱是由遗传控制的花型多态性现象,分二型花柱(distyly)和三型花柱(tristyly),而二型花柱分长柱型(long-styled morph)和短柱型(short-styled morph)两种花型[8-9]。二型花柱目前已在28个被子植物科199个属中报道,并且在茜草科中报道最多[10]。交互式雌雄异位(reciprocal herkogamy)是二型花柱植物最典型的特征,两种花柱型的性器官在空间上存在交互式对应关系[8],并且在其他花部结构通常表现出多态性,如柱头裂片长度[11]、花粉粒外壁纹饰[12-13]、花粉数量[6]和花粉粒大小[14-15]等方面。二型花柱植物通常伴随着自交不亲和,仅花型间授粉才能结实[16]。自交不亲和,促进型间授粉的原因可能在于柱头上型间花粉的花粉管可以伸长进入子房。Baker[4]对Mirabilis的研究表明型间花粉能在柱头上萌发,顺利通过花柱进入到子房,而型内花粉不能长出柱头表面或者在花柱道中终止生长。Wu等[17]发现Mussaendamacrophylla型间授粉的花粉管生长比型内授粉和自花授粉都要快。邵剑文等[18]表明长柱型为母本,型间授粉的花粉管的生长速度明显比型内花粉的快,而在短柱型为母本,型间授粉的萌发率最高。
二型花柱植物促进异交,提高传粉精确性,要求植物与传粉者之间有更严格的机械匹配[19](mechanical fit, 即植物花与动物的传粉器官在形态结构上的匹配,如植物的花冠管长和动物的吻长)。例如Primulasecundifora和Primulapoissonii是管状花,体型大的红腹熊蜂(Bombusfriseanus)的个体与报春花结构更适应,将其吻深入花冠管基部吸取花蜜时,吻部会沾上花粉,而其在访问其他花时吻部的花粉会触碰柱头,从而实现花粉的移出和落置,在2011年,2012年主要的访花者是蜜蜂和体型小的红腹熊蜂(个头小,体型与报春花属特征不匹配),导致柱头上型间花粉的比例低,而2013年这两种报春上主要的访花者是体型大的B.friseanus,提高了柱头上型间花粉的比例[20]。茜草科滇丁香属滇丁香(Luculiapinceana)是典型的二型花柱植物,白色,细长管状花,具有浓郁的香气。万友名等[21]研究发现滇丁香的有效传粉者是小蜜蜂(ApisfloraeFabricius),其观察访花的时间是从8点到18点,忽略了晚上潜在的传粉者;并且根据植物花部特征与虫体特征机械匹配以及传粉综合征(pollination syndrome,即与吸引特定类型传粉者相关的花部特征组, 例如管状花冠、白色或者乳白色,具有浓郁的气味,花蜜浓度相对较低等是典型的蛾类传粉综合征),A.florae可能不能为滇丁香进行有效的传粉,更不能促进异交。二型花柱植物滇丁香是否能促进异交,并且是哪种动物进行有效的传粉,需要进一步研究。本文主要探究以下问题:(1)比较自然居群中滇丁香长、短柱型植株的数量以及花部特征是否存在显著性差异。(2)自然状态下长、短柱型柱头上的型间花粉、型内花粉的数量及比例是否有显著性差异。(3)型内和型间授粉处理,比较二者花粉的萌发率和花粉管的长度是否有显著性差异。(4)人工控制授粉检验滇丁香是否型内亲和。(5)进行访花观察,确定滇丁香有效传粉昆虫的类型,测定传粉昆虫的体型特征,并分析比较它是否与滇丁香的花部特征相匹配。
1 材料与方法
1.1 材料
滇丁香(Luculiapinceana)隶属茜草科(Rubiaceae)滇丁香属,是一种多年生常绿乔木或灌木。滇丁香主要分布于亚洲东南部,其中云南省的西南、西北、东南等地分布最为广泛,生于海拔600—3000 m处的山坡、山谷、溪边、林边以及灌丛中。花期较长,一般在6—12月,花果期重叠。伞房状的聚伞花序簇生枝顶,花序大而密集,株型优美,枝叶翠绿,花开繁茂艳丽具芳香,具有一定的药用价值,是一种极具观赏价值和推广前景的野生花卉资源。本研究供试材料为云南省文山州麻栗坡大锡厂老山省级自然保护区(104°49′ 62" E,23°94′ 8" N)的野生滇丁香。该地区属于常年湿润季风气候,多雾天气,雨量充沛,季节分布不均,冬无严寒,夏无酷暑。
1.2 方法
1.2.1比较自然居群长、短柱型的植株数量与花部特征
为比较自然居群中滇丁香长、短柱型的植物数量是否有显著性差异,我们将老山省级自然保护区山顶至山脚分5个居群(居群间隔约2 km),并统计每个居群长、短柱型的植株数量。
为比较滇丁香长柱型、短柱型花部特征,随机选取长柱型与短柱型植株各30株,每株随机选择1朵花,用游标卡尺测量(精确到0.01 mm) 叶片长与宽,萼片长与宽,花长与宽,花瓣长与宽,筒深,筒直径,开口直径;然后将花纵向剖开以便于测量花冠管内的花部特征:雌蕊长,雄蕊长,花药长、宽与厚度,柱头长、宽与厚度。为了比较长、短柱型花粉的大小,每个花柱型随机选择30株植株,每株用尖头镊子摘取1个成熟的花药并固定在装有75%酒精的2 mL离心管中。每个花粉液体样本随机取一滴置于载玻片上,分别用荧光显微镜拍照,每张图片随机选择3粒花粉,用图像分析软件(Digimizer Version 4.6.0)测量花粉的长度(Length, 简写L)和宽度(Width, 简写W),那么花粉的长度为(L1+L2+L3)/3,花粉的宽度为(W1+W2+W3)/3。为了比较长、短柱型的单花花粉数量和胚珠数,随机选取长、短柱型的植株各30个,每个植株选取一个花苞,保存在装有75%酒精的离心管中。在实验室内,用镊子将单花花苞的花药和子房分开,分装在不同的离心管中,对应标记。用镊子充分捣碎花药并定容至2 mL,充分震荡摇匀后,移液枪取5 μL液体滴于载玻片,在光学显微镜下统计花粉的数量。每个样本滴取三滴进行统计,花粉数分别记为a1,a2,a3,花苞中花粉的总数量 =(a1+a2+a3)/3 × 稀释倍数。在解剖显微镜下观察花苞中的胚珠,并统计胚珠数量。花粉胚珠比(P/O)= 单花花粉数量/单花胚珠数量。为比较长、短柱型的单花期,随机选择两柱型植株各30株,每个植株上随机选择1个花苞进行标记,花苞开放记为第1 d,每隔1 d观察花的状态,直至花朵凋落,记为单花期。
1.2.2比较长、短柱型的花粉活力和柱头活性
为了比较长、短柱型的花粉活力和柱头活性,两种花型分别随机选择30个植株,每株随机选择1个次日开放的花苞进行标记,次日收集开放花朵的花粉与柱头,用MTT染色法(3 -(4, 5-dimethylthiazol- 2-yl)- 2, 5-diphenyl tetrazolium bromide)进行测定花粉的活力(有活力的花粉被染成深紫色)[22]。用过氧化氢液体测定柱头活性(若有活性,柱头周围会产生气泡,气泡产生越多表明活性越强)[23]。
1.2.3自然种群植株柱头上花粉的落置
选择晴朗的天气,随机选择长柱型、短柱型滇丁香各30株,每个植株选择一朵花,用镊子摘取其柱头并装入含有FAA溶液(formalin-acetic acid- 70% alcohol,体积比是5:5:90)的1.5 mL离心管中,带回实验室,用8 mol/L的NaOH溶液软化4 h,然后镊子充分捣碎柱头呈均匀的悬浊液,置于高速(8000 r/min)离心机下离心5 min,弃上清液,定容至0.5 mL,用移液枪吸取离心管中的全部样本溶液滴于载玻片上,置于光学显微镜下(Nikon E100)统计型间和型内的花粉数量。
1.2.4不同花型花粉的萌发以及花粉管的生长
为了比较型内授粉,型间授粉和混合授粉(自然种群植株柱头上同时落置长、短柱型花粉,该授粉处理是模拟自然状态)是否会影响花粉的萌发以及花粉管的伸长,我们随机选择长、短柱型植株各30株,每个植株随机标记3个花苞(绑以不同颜色的尼龙线加以区分),进行去雄套袋处理,直至柱头成熟。选择晴朗的天气,从距离受体植株至少30 m以外的其他植株上选择新鲜开裂的花药,分别收集长、短柱型花粉作为花粉源。用小毛笔进行授粉,每个植株上进行3 种授粉处理:(1)型内授粉,(2)型间授粉,(3)混合授粉(长短柱型花粉混合),每个成熟的柱头涂满花粉,然后重新套袋。6 h后收集柱头,储存在装有FAA固定液的离心管中。在实验室中,柱头首先在蒸馏水中清洗3次,然后于室温在8 mol/L 的NaOH 溶液中软化4 h,再置于蒸馏水中漂洗3 min,再在苯胺蓝溶液(0.1% 质量浓度)中染色3 h。用荧光显微镜观察柱头,统计柱头上花粉的总数量,萌发的花粉数量;对于花粉管的长度,先用荧光显微镜自带的软件拍照,然后再用图像分析软件(Digimizer Version 4.6.0)测量花粉管的长度[22]。
1.2.5人工控制授粉实验
为检验滇丁香的繁育系统类型以及是否型内亲和,长柱型和短柱型植株分别作为母本(花粉受体)进行3种不同授粉(花粉供体)处理。随机选择长柱型、短柱型各30个植株,将每个植株上3朵形态发育一致的花苞进行标记,分别进行以下3种处理:(1)自然对照;(2)去雄,套袋,等到柱头成熟时授异株同型花粉,检验型内花粉是否亲和;(3)去雄,套袋,授异株异型花粉,检验型间花粉是否亲和。授粉处理选择在晴朗的天气进行。待果实成熟后野外收集果实带回室内,在解剖显微镜下统计结实情况。计算公式:结实率=单果种子饱满数/单果种子总数 × 100%。
1.2.6传粉昆虫及其传粉行为
野外随机选择5个居群,每个居群随机选择大约100朵开放的长、短柱型的滇丁香单花并标记(两种柱型分别大约50朵)。在晴朗的天气进行访花观察,观察时间从早上7点到晚上10:30,每半小时为一个时间段,观察并拍照记录访花昆虫的种类和访花者行为,记录昆虫访问的花朵数,统计每个居群中标记的花朵数。若访花昆虫将喙伸入到花冠筒中吸取花蜜,并触碰到花药和柱头,则属于有效传粉者。访花观察的时间为2019年6月17号到7月15号,2020年7月12到8月12号。访问频率等于访问次数/每朵花/h。
2020年7月选择夜间晴朗天气,利用高压汞灯(450W,E40螺口,上海亚明1923自镇流)在实验地点诱捕滇丁香的主要传粉昆虫30只,固定并制作为昆虫标本,用游标卡尺测量其体长、喙长、头宽、触角长、头宽、胸部长、胸部宽、胸部厚、腹部长、翅展。
1.3 数据处理与分析
不同的数据类型符合不同的函数分布,根据数据的类型,选择相应的分布函数进行分析。采用拟合优度(G-test of goodness-of-fit)检验自然居群中长、短柱型滇丁香的植株数量是否偏离1∶1。为了比较长、短柱型滇丁香的花部特征,不同的授粉处理花粉管的长度,不同访花昆虫的访问频率是否存在显著性差异,采用广义线性模型(generalized linear model, GLM)中正态分布(normal distribution)恒等关联函数(identity-link function)进行分析,花部特征,花粉管的长度,访问频率分别作为因变量,长、短柱型,不同的花粉来源、不同母本,不同的访花昆虫分别作为自变量。采用泊松分布(poisson distribution)对数关联函数(loglinear-link function)比较长、短柱型的滇丁香的花苞中的总花粉数、胚珠数,其中花粉数和胚珠数作为因变量, 两种花柱型作为自变量。为了比较自然状态下柱头上落置的型间花粉和型内花粉是否存在显著性差异,用泊松分布-对数关联函数进行分析,其中花粉数量作为因变量,不同的花粉来源作为自变量。采用二元Logistic关联函数分析比较不同授粉处理柱头上花粉的萌发率和不同授粉处理的结实率,柱头上萌发的花粉数作为因变量, 柱头落置的总花粉数作为一次实验中总的变量, 不同的花粉来源、不同的母本作为自变量;饱满种子数为因变量,种子总数为一次试验中总的变量,不同授粉处理和不同柱型为自变量。本研究试验数据用IBM SPSS Statistics 20进行分析,Origin 2018制图。
2 结果
2.1 花部特征的比较
在自然居群中,滇丁香长柱型的柱头裸露于花冠管外,柱头所占位置明显高过花药;短柱型的柱头隐藏于花冠管之中,花药位置高于柱头(图1)。5个自然居群中滇丁香长、短柱型植株的数量没有偏离1∶1(表1)。

图1 滇丁香两种表型的单花Fig.1 Single flower of two morphs of Luculia pinceanaL-morph:长柱型;S-morph:短柱型;标尺上边是短柱型单花的解剖图,标尺下边是长柱型单花的解剖图
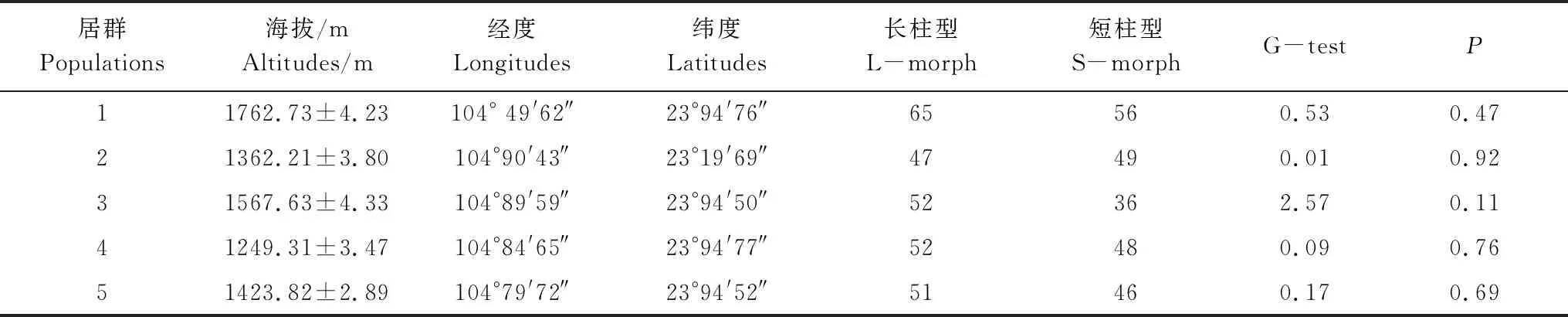
表1 比较不同居群滇丁香长、短柱型植株数量(拟合优度检验)
广义线性模型(GLM)对不同花型花部特征进行比较分析结果表明:长柱型的筒深、筒直径、开口直径、雄蕊长、花粉粒体积、柱头长显著小于短柱型(P<0.01),而长柱型的叶片长、叶片宽、雌蕊长(花柱长)、柱头厚均显著大于短柱型(P<0.01)(表2)。长柱型和短柱型的萼片长、萼片宽、单花期、花长、花宽、花瓣长、花瓣宽、花药体积、柱头宽、花粉数、胚珠数、花粉/胚珠比没有显著性差异(P> 0.05)。两种柱型的花粉/胚珠比(P/O)介于244.70~2588.00之间,根据Cruden[24]的标准,滇丁香属兼性异交(Facultative xenogamy)类型。长柱型的雌蕊长和短柱型的雄蕊长没有显著性差异(Waldχ2=0.80,df=1,P=0.37),长柱型的雄蕊长和短柱型的雌蕊长没有显著差异(Waldχ2=2.70,df=1,P=0.10),表现出交互式雌雄异位的花部特征(表2)。
开花当天长柱型的花粉活性(85.95±2.16)%和短柱花粉活性(88.34±2.16)%没有显著性差异(Waldχ2=0.61,df=1,P=0.43)。开花当天长柱型、短柱型植株的柱头呈黄绿色,柱头二叉分开,均有可授性。
2.2 自然种群植株柱头上花粉的落置
对自然种群植株柱头上花粉的类型和数量分析发现,柱头上型间花粉的数量(1198.38±329.10)显著低于(Waldχ2=38.13,df=1,P<0.001)型内花粉的数量(3663.37±188.23),型间花粉所占的比例(24.65±1.59)%显著低于(Waldχ2=340.97,df=1,P<0.001)型内花粉所占的比例(75.35±1.59)%。其中,长柱型柱头上型间花粉的数量(1001.77±206.47)显著低于(Waldχ2=23.10,df=1,P<0.001)型内花粉的数量(3895.07±425.65)。短柱型柱头上型间花粉的数量(1395.00±290.82)显著低于(Waldχ2=13.26,df=1,P<0.001)型内花粉的数量(3431.67±456.13)。长柱型柱头上型间花粉所占的比例(20.46±1.94)%显著低于(Waldχ2=258.45,df=1,P<0.001)型内花粉所占的比例(79.54±1.94)%。短柱型柱头上型间花粉的比例(28.90±2.29)%显著低于(Waldχ2=130.57,df=1,P<0.001)型内花粉所占的比例(71.10±2.29)%。

表2 长柱型、短柱型滇丁香花部特征(平均值±标准误)的比较(广义线性模型)
2.3 不同花型花粉萌发以及花粉管生长的比较
对不同授粉处理花粉萌发率以及花粉管生长比较发现,短柱型花粉在柱头的萌发率(76.35±2.10)%显著高于混合花粉(68.72±2.52)%和长柱型花粉(63.97±2.20)%的萌发率,其中混合花粉和长柱型花粉的萌发率没有显著性差异(Waldχ2=1.62,df=1,P=0.20)(图2)。长柱型为母本的花粉萌发率(65.42±1.78)%显著低于短柱型为母本的花粉萌发率(75.15±1.93)%,且花粉供体和花粉受体对滇丁香的花粉萌发率的影响有交互作用(表3)。不同花粉供体对滇丁香花粉萌发率的影响在不同的花粉受体间不同,长柱型为母本,授长柱型花粉的萌发率显著低于授短柱型花粉和混合型花粉的萌发率(Wald χ2= 21.71,df=2,P<0.001),授短柱型花粉与授混合型花粉的萌发率没有显著差异(Waldχ2=2.43,df=1,P=0.12);短柱型为母本,授长柱型花粉和授短柱型花粉的花粉萌发率显著大于(Waldχ2=8.78,df=2,P=0.01)授混合花粉的萌发率,而授长柱型和授短柱型的萌发率没有显著性差异(Waldχ2=0.09,df=1,P=0.77)(图2)。
对花粉管长度分析发现,长柱型花粉(120.15±3.25)μm、短柱型花粉(117.13±3.25)μm、混合花粉(119.69±3.25)μm在柱头上的花粉管长度没有显著性差异。长柱型为母本的花粉管长度(130.34±2.36)μm显著高于短柱型为母本的花粉管长度(107.64±2.36)μm(Waldχ2= 46.13,df=1,P<0.001)(表3,图2),且花粉供体和花粉受体对滇丁香的花粉管长度的影响没有交互作用(表3,图2)。长柱型为母本,授短柱型花粉跟混合授粉的花粉管长度没有显著性差异(Waldχ2=7.05,df=1,P=0.01),且都显著高于(Waldχ2=25.26,df=1,P<0.001)授长柱型花粉的花粉管长度。短柱型为母本,授长柱型花粉的花粉管长度显著高于(Waldχ2=36.93,df=2,P<0.001)授短柱型花粉和混合授粉的花粉管长度。
总体上看,在不同花粉受体上,型间授粉的花粉萌发率(76.76±2.07)%显著性大于(Waldχ2=18.00,df=2,P<0.001)型内授粉(63.63±2.19)%和混合授粉(68.72±2.50)%的花粉萌发率。型间授粉的花粉管长度(132.25±2.92)μm显著性大于(Waldχ2=44.41,df=2,P<0.001)型内授粉(104.94±2.92)μm和混合授粉(119.68±2.91)μm的花粉管的长度(图2)。

图2 不同处理滇丁香的花粉萌发率和花粉管长度的比较。长柱型花粉、短柱型花粉、混合型花粉分别为花粉供体, 长柱型和短柱型滇丁香分别作为花粉受体Fig.2 Comparisons of pollen germination and pollen tube length of pollen donors from L-morph, S-morph and Mix, with the L-morph and S-morph in Luculia pinceana as pollen recipients in vivo不同字母表示不同的授粉处理有显著差异
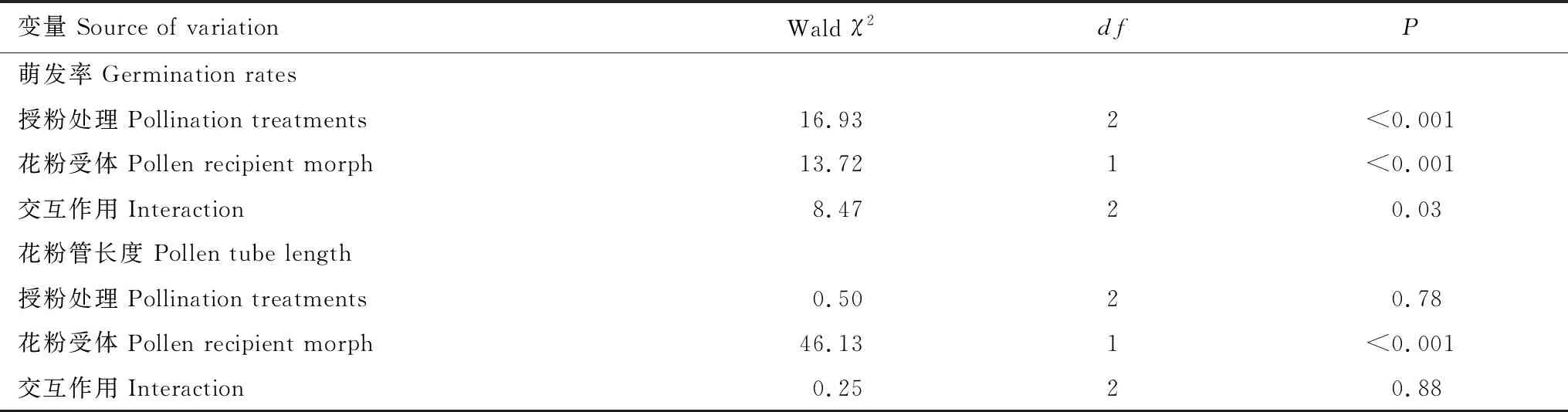
表3 授粉处理(长柱型、短柱型和混合型的花粉),花粉受体(长柱型和短柱型分别为母本)及其交互作用对滇丁香柱头花粉的萌发率、花粉管长度的影响(广义线性模型)

图3 不同授粉处理滇丁香结实率的比较 Fig.3 Comparison of seed sets of Luculia pinceana with different pollination treatments不同字母表示不同的授粉处理有显著性差异
2.4 人工控制授粉试验
滇丁香型间异交的结实率(83.13±4.75)%显著高于(Waldχ2=31.52,df=1,P<0.001)型内异交(28.41±4.63)%的结实率,与自然对照的结实率没有显著性差异(69.26±5.98)%(Waldχ2=2.97,df=1,P=0.09)。长柱型为母本,型间异交和自然对照的结实率没有显著性差异(Wald χ2= 1.46,df=1,P=0.23),均显著性高于(Waldχ2=8.77,df=2,P=0.01)型内异交的结实率。短柱型为母本,型间异交和自然对照的结实率没有显著性差异(Waldχ2=1.18,df=1,P=0.28),均显著性高于(Waldχ2=32.11,df=2,P<0.001)型内异交的结实率。长、短柱型自然状态下结实率没有显著性差异(Waldχ2=0.45,df=1,P=0.51)(图3)。
2.5 传粉昆虫及其传粉行为
访问滇丁香的昆虫主要有三类:长喙条背天蛾(Cechenenalineosa,鳞翅目天蛾科条背天蛾属),熊蜂(Bombussp.,膜翅目蜜蜂科熊蜂属),蜜蜂(Apis,蜜蜂科蜜蜂属)(图4)。熊蜂和蜜蜂体型相对条背天蛾较小。三者的访问行为也完全不同,条背天蛾访花时,其飞至花冠口,长喙深入花冠管并取食花冠筒基部的花蜜,访花过程中吻部会沾上花粉。熊蜂访花时落至花冠管基部,直接进行盗蜜。蜜蜂大多时间只访问短柱型花而较少访问长柱型花,通常两对胸足钩住花药,一对后足搭在花冠上维持平衡,用口器取食和盗取花粉。条背天蛾的访问频率为(0.04±0.06)次/花/h,熊蜂为(0.47±0.06)次/花/h,蜜蜂为(0.29±0.09)次/花/h。条背天蛾的虫体特征见表4,其中吻长显著大于长柱型滇丁香的筒深(Waldχ2=87.86,df=1,P<0.001),也显著大于短柱型的花筒深(Waldχ2=6.86,df=1,P=0.01)(花筒深的数据见表2)。

图4 滇丁香的访花昆虫Fig.4 Visiting insects of Luculia pinceana
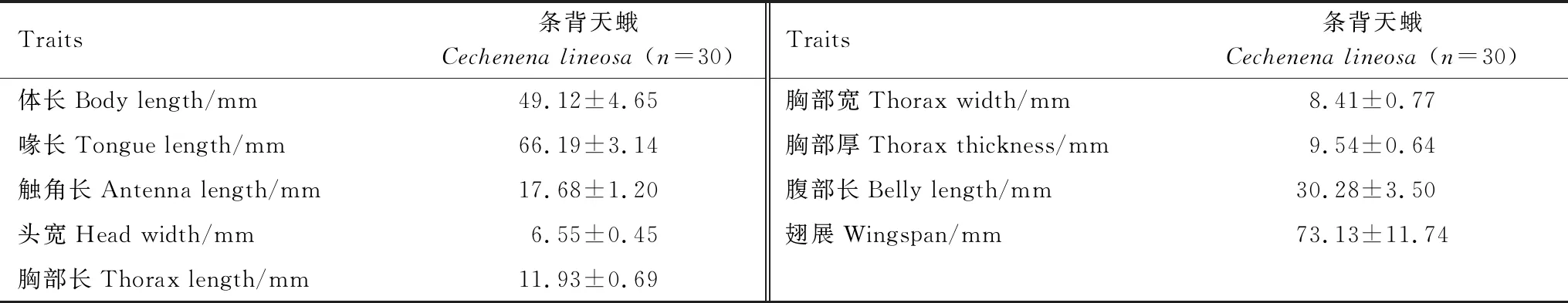
表4 条背天蛾体型特征的测量(平均值±标准误)
3 结论与讨论
3.1 花部特征的多态性
二型花柱植物的花部形态特征具有多态性[2,6,13- 14],例如荞麦属(Fagopyrum)、紫草属(Lithospermum)的花瓣大小在长、短柱型之间存在差异[2]。艳丽耳草(Hedyotispulcherrima)长柱型和短柱型的柱头裂片长度、花粉大小和淀粉含量等具有多态性[11]。本研究在检测花粉数量上表明,长、短柱型的花粉数量没有显著性差异,这种特征与许多二型花柱植物的特征不同[11,25],但与HedyotisacutangulaChamp的报道相似[15]。一般认为,二型花柱植物短柱型的花粉粒体积大于长柱型[11,14- 15,25-27],而我们的研究结果同样支持这一结论。研究发现,短柱型的柱头表面积大于长柱型,这种型间不对称性可能是由于短柱型的柱头隐藏在花冠之中,位置较低,相比长柱型更难接收到花粉,因而短柱型通过增大柱头表面积来提高花粉的接触面积,从而弥补花粉流的不对称性。滇丁香的叶片长、叶片宽、筒深、筒直径、开口直径,柱头厚在长、短柱型间也表现出多态性,是植物保证繁衍顺利而采取的一种生殖策略[28]。此外,我们发现自然状态下滇丁香长、短柱型植株的数量没有偏离1∶1,这在异型花柱植物中是一种普遍现象,能避免雄性资源和雌性资源的浪费,有利于维持居群长期的稳定[29]。
3.2 自然种群植株柱头花粉的落置
Armbruster等[30]提出的不精确指数(inaccuracy index)报道了花粉流中相互作用关系,提出了精确的花器官交互式结构更有利于型间花粉在柱头上的落置[14]。在自交不亲和的二型花柱植物中,型间花粉在柱头上的比例直接反应了型间授粉的成功率。本研究发现滇丁香虽有精确的交互式异位结构特征,但自然种群植株柱头上型间花粉的比例都低于30%,其他二型花柱植物中也有类似报道[17],这一结果似乎与前人提出的异型花柱交互雌雄异位结构促进型间花粉精确传递的假说不一致。
自然植株种群柱头上落置大量的型内花粉可能是自花花粉或者同株异花花粉的干扰。有研究表明,没有去雄情况下,报春花属等柱头上落置大量的型内花粉[20]。但是,对荞麦属甜荞(Fagopyrumesculentum)进行单花去雄(排除自花花粉的干扰)和整株去雄(排除同株异花花粉的干扰)处理后,发现柱头上31.75%为型间花粉,17%为型内花粉,柱头上亲和花粉比例增加,表明二型花柱植物可促进型间异交,提高传粉精确性[31]。
3.3 花粉管萌发和繁育系统
二型花柱植物花粉的萌发及花粉管的伸长可能影响植物的繁殖成功[24-26,32-33]。Chen等[32]发现Mussaendashikokiana自交和异交的花粉管生长率差异不显著。Nishihiro等[33]发现报春科自花花粉的萌发率明显低于型间授粉,自花花粉和型间花粉的花粉管生长速率没有差异。Klein[34]提出P.nuda型内和型间花粉的萌发率及花粉管的生长是相似的,而张大爱等[35]研究表明commonbuckwheat型内授粉的花粉管在柱头或花柱中则是被抑制使结实率降低。本研究表明短柱型为母本,有利于花粉的萌发,而以长柱型为母本,有利于花粉管的生长,自然状态下长、短柱型的结实率没有显著性差异。在不同的花粉受体上,型间授粉的花粉萌发率和花粉管长度显著大于型内授粉,人工型间授粉的结实率相应也显著高于型内授粉,综合人工授粉结实和P/O比的结果,滇丁香是一种兼性异交植物。异型花柱植物具有典型的型内(自交)不亲和性[11,16],但也有一些二型花柱植物具有型内亲和[2],而一定的型内亲和也是对植物的一种生殖保障[36]。
3.4 植物花部特征与传粉昆虫虫体的相互适应
植物的花部特征反映了对某一类群传粉者的适应[19,37]。马达加斯加的兰科植物彗星兰 (Angraecumsesquipedale)的花距长30—40 cm,依靠吻长约30 cm的天蛾为其传粉[38]。石蒜科水仙(Narcissuspapyraceus)是管状花,适应于长吻的昆虫为其传粉,当长吻昆虫伸入花冠管基部吸蜜时,长、短柱型花粉就落置到昆虫吻长的不同部位,随后昆虫在长、短柱型间互访时就能成功的完成型间传粉[39]。茜草科绣球茜(Dunniasinensis)的交互式雌雄异位结构与其传粉者(蜂类和蝶类)的体位相匹配,使得传粉者头部携带的是长柱型花的花粉,而腹部携带的是短柱型花的花粉,最终从而实现植物间的型间传粉[25]。不过在二型花柱植物樱草(Primulasieboldii)中发现,狭窄的花冠筒结构严格限制了的传粉者的吻,使得植物与传粉者无法完全的匹配,导致短柱柱头上亲和花粉的落置和结实率受到影响[40]。滇丁香主要是通过与其花冠管长度相匹配的长吻条背天蛾实现有效的传粉,而小蜜蜂主要是收集花粉,并不能为滇丁香进行有效的传粉。昆虫的不同访花行为对二型花柱植物适合度的影响是多样和复杂的。长喙天蛾的传粉效率还需要探究。熊蜂从花筒基部盗蜜和蜜蜂直接收集花粉是如何影响滇丁香的适合度以及传粉精确性还需要进一步研究。
二型花柱植物滇丁香长、短柱型植物数量在自然居群中没有显著性差异,花特征表现为典型的交互式雌雄异位,自然状态下柱头上落置的型间花粉的数量显著低于型内花粉的数量。滇丁香通过有利于型间花粉的萌发和花粉管的伸长从而提高型间授粉的结实,型内授粉也有一定的结实。长喙条背天蛾的虫体特征与滇丁香的花特征相适应,是其有效的传粉者。前人对二型花柱植物提高传粉精确性的研究大多比较了长、短柱型的花部特征,并检验是否自交亲和,也有部分研究统计自然状态下柱头上花粉落置情况,以及比较型内、型间花粉的萌发率和花粉管长度。对于滇丁香来说,本研究首次详实地比较长、短柱型的花特征,包括单花的花粉数量、大小、花粉/胚珠等;通过不同的授粉处理,发现滇丁香在是有利于型间花粉的萌发和花粉管的伸长而促进了型间授粉的结实;并首次比较条背天蛾、熊蜂、蜜蜂等昆虫的访花行为,确定条背天蛾是其有效的传粉者。该研究为二型花柱植物可通过促进型间花粉的萌发和花粉管的伸长而促进异交、植物花部特征与传粉者虫体特征机械匹配提供了新的例证。
致谢:感谢广西植物所刘长秋副研究员、云南省文山州麻栗坡县郭喜兵先生、贵州师范大学生命科学学院相孟达同学对野外实验的协助。
