杉木第3代种子园自由授粉子代测定及选择
2021-09-29吴炜
吴 炜
(福建省将乐国有林场,福建 将乐 353300)
杉木(Cunninghamialanceolata(Lamb.) Hook.)是我国南方16个省(区)广泛栽培的重要用材针叶树种[1],也是国家“十三五”期间重要的发展规划树种之一[2]。自20世纪50年代开始,福建省持续开展了近70 a的杉木遗传育种及应用研究[3]。特别是第3代育种效果显著,嫁接营建的杉木第3代种子园种子产量可达42.70 kg·hm-2[4-5],且第3代良种生长优势明显,5年生时树高、胸径、单株立木材积年均生长量分别达到1.42 m、2.19 cm、0.00798 m3,明显优于杉木第2代种子园良种[5-9]。随着杉木育种研究逐步深入,福建省已启动杉木第4代育种研究,巩固第3代育种成果的阶段[3]。因此,对建成的杉木第3代种子园开展子代测定势在必行,因为根据子代测定结果对种子园去劣疏伐,后向优良家系作为第3.5代种子园建园亲本,是进一步提高杉木第3代种子园遗传增益的必要途径。同时,在子代测定林中前向选择优良单株充实杉木第4代育种群体,选择无性繁殖原株拓宽杉木优良无性系选择基础是杉木第4代育种和优良无性系繁育不可或缺的途径。
1 材料与方法
1.1 试验地概况
试验地位于福建省将乐国有林场明头山工区061林班11大班010小班,为杉木中心产区[10],地理坐标117°26′10″E、26°43′5″N。属中亚热带季风气候,年均气温18.7 ℃,年平均相对湿度82%,年无霜期287 d,年均降水量1669 mm。低山丘陵,海拔190~283 m,坡度25°~31°,坡向西北。土壤为山地红壤,Ⅱ类立地。前荐为杉木人工林纯林。
1.2 试验材料与方法
参试材料为福建省洋口国有林场杉木第3代种子园自由授粉子代,2010年春育苗,2011年1月20日营建自由授粉子代测定林。参试家系30个,以福建省洋口国有林场杉木第2代种子园混合种(洋口2代,CK1)、福建省光泽华桥国有林场杉木第3代种子园混合种(光泽3代,CK2)、福建省沙县官庄国有林场杉木第3代种子园混合种(官庄3代,CK3)、福建省将乐国有林场杉木第3代种子园混合种(将乐3代,CK4)、福建省良种审定无性系“闽S-SC-CL-010-2018”(闽杉1号,审定前名为2C无性系,CK5)组培苗,广西融水种源(广西融水,CK6)、福建省沙县官庄国有林场红心杉木种子园混合种(官庄红心杉,CK7)7个良种为对照。田间试验设计为完全随机区组设计(RCB),37个处理,9次重复,5株单列小区。株行距1.7 m×2.0 m,种植穴规格50 cm×40 cm×30 cm。
根据《杉木速生丰产林栽培技术规程》[10]管理试验林,但未进行修枝和间伐。试验林1~3 a林龄时每木调查树高,4~6 a林龄时每木调查树高、胸径,9 a林龄时每木调查树高、胸径,并选取每个小区中的平均木在树干1.3 m高处采集木芯,生长锥内径5 mm,木芯通过髓心,通透树干,无节疵。
1.3 统计分析方法
根据树高、胸径估算单株立木材积[11]:V=0.00005877042D1.9699831H0.89646157。木材红心率:rr(%)=(木芯红心长/去皮木芯长)2×100%[12]。木材基本密度:wd(g·cm-3)=1/[(M-M1)/M1+1/DW],式中:M为木芯水饱和时的质量(g);M1为木芯烘干时(105 ℃)的恒重(g);DW为构成细胞壁的木材物质的比重,取平均值1.53[12]。
2 结果与分析
2.1 表型变异分析
试验林平均树高、胸径、单株立木材积累年生长量及其在家系间的表型变异系数见图1~图3。9 a时,试验林平均树高、胸径、单株立木材积分别为10.16 m、13.48 cm、0.08405 m3,年均生长量分别为1.13 m、1.5 cm、0.00934 m3,家系间表型变异系数分别为3.82%、6.03%、13.54%。造林当年(1 a)家系间树高表型差异较大,第2年(2 a)后表型差异趋于平缓,这主要是由于造林当年植株根系处于恢复期,树高生长受影响所致。4 a以后,树高、胸径、单株立木材积在家系间均有稳定的表型变异,其中单株立木材积变异最丰富,表型变异系数达13.00%以上。
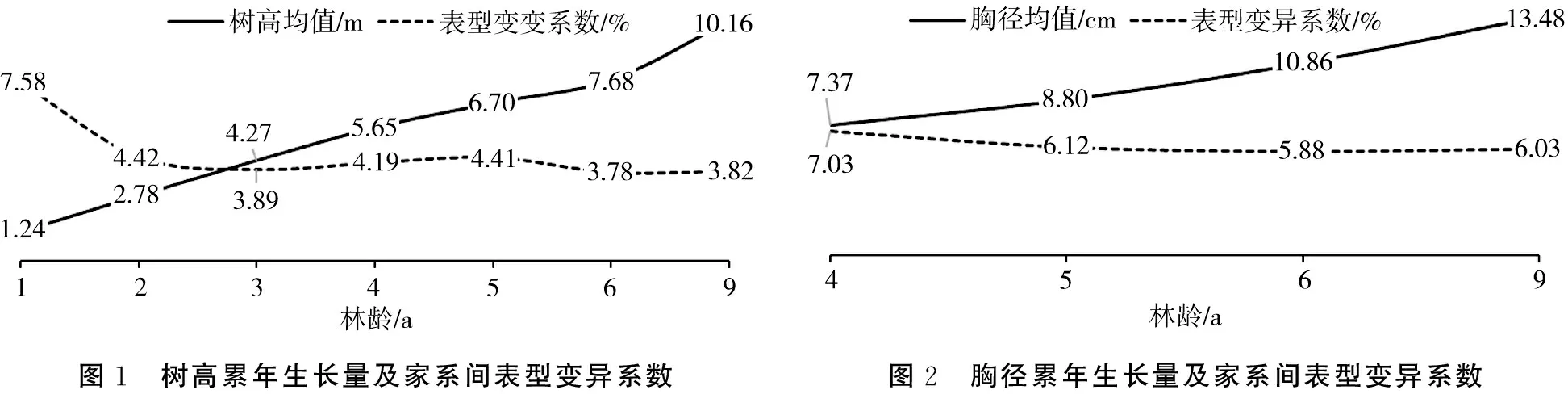
图1 树高累年生长量及家系间表型变异系数图2 胸径累年生长量及家系间表型变异系数

图3 单株立木材积累年生长量及家系间表型变异系数
参试的30个家系树高、胸径、单株立木材积均值与7个对照均值的单样本T检验差值见表1~表3。从表1可知,参试家系平均树高在各林龄时均显著高于洋口第2代种子园混合种和沙县官庄第3代种子园混合种;但与光泽华桥第3代种子园混合种相比,造林后前3 a具有明显的高生长优势;与将乐第3代种子园混合种相比,造林前3 a高生长处于劣势,但从第4年开始表现出了高生长优势。参试家系树高与2C无性系组培苗相比,造林后前5 a具有明显高生长优势,但从第6年开始转变为明显的劣势;与广西融水种源和官庄红心杉种子园混合种相比,造林前期差异不显著,从第5年开始具有明显的高生长优势。从表2、表3可知,参试家系胸径和单株立木材积均值与实生良种相比,表现出跟树高一致的生长差异,但与2C无性系组培苗相比,具有明显的胸径和材积生长优势。总体而言,参试家系生长性状均值在造林后6 a时,确立了其与对照的生长差异特征:树高显著高于实生良种,但低于2C无性系组培苗;胸径和单株立木材积显著高于实生良种和2C无性系组培苗。
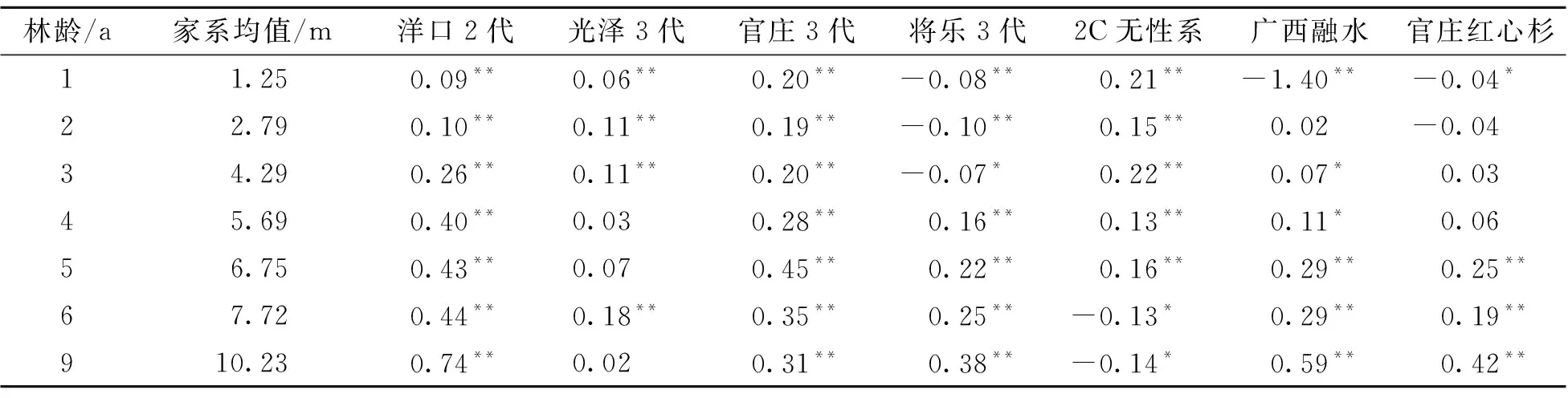
表1 参试家系均值与对照树高单样本T检验差值

表2 参试家系均值与对照胸径单样本T检验差值

表3 参试家系均值与对照单株立木材积单样本T检验差值
9 a林龄时参试家系各性状均值与对照的单样本T检验差值见表4,并将差值换算成百分比(家系均值与对照的差除以家系均值)后绘制了图4。参试家系与7个对照相比,具有明显的生长优势,但红心材率明显低于官庄红心杉种子园5.32%,而木材基本密度除了与洋口第2代混合种和沙县官庄第3碟混合种差异不显著外,明显低于其它对照3.76%~10.66%。参试家系生长性状与木材品质性状的表型逆向差异性对优良家系的联合选择不利,但其是否受到遗传影响,还需进一步遗传分析。

图4 参试家系均值与对照对比性柱状图

表4 参试家系9 a林龄时与对照性状单样本T检验差值
2.2 遗传变异分析
在不同林龄时,树高、胸径、单株立木材积家系间差异均极显著,家系与区组的交互作用也均极显著(表5~表7)。1~9 a林龄时,树高的广义遗传力介于0.6386~0.7723间,狭义遗传力介于0.1992~0.3609间;造林当年(1 a)苗木根系恢复对家系间生长差异造成影响,遗传力估值较大,其它林龄时遗传力估值均较为稳定。4~9 a时,胸径的广义遗传力介于0.7226~0.8006间,狭义遗传力介于0.2991~0.3481间;单株立木材积的广义遗传力介于0.6627~0.7550间,狭义遗传力介于0.2445~0.2958间。9 a时红心材率在家系间差异不显著;木材基本密度在家系间差异极显著(表8)且广义遗传力为0.4751,狭义遗传力为0.3655。可见,测定群体的树高、胸径、单株立木材积、木材基本密度均受到中度偏下的遗传控制,而红心材率不受遗传影响,可对群体的生长性状和木材基本密度进行家系选择(后向选择)。不同林龄阶段,狭义遗传力与广义遗传力的比值树高介于0.3010~0.4673间,胸径介于0.4022~0.4348间,单株立木材积介于0.3645~0.3918间,而木材基本密度为0.7693。表明在受遗传控制的性状中,树高、胸径、单株立木材积受到明显的非加性遗传效应影响,前向选择速生优良单株作为无性繁殖材料可获得比家系选择更高的遗传增益;而木材基本密度主要受到加性遗传效应影响,在测定木材基本密度的样株中选择优良单株作为无性繁殖材料并不能获得比家系选择更高的遗传增益。

表5 树高方差分量及遗传力

表6 胸径方差分量及遗传力

表7 单株立木材积方差分量及遗传力

表8 红心率及木材基本密度方差分量及遗传力(9 a林龄)
2.3 遗传相关分析
参试家系树高、胸径、单株立木材积早晚表型相关系数和遗传相关系数见表9~表11。树高1~9 a间,胸径和单株立木材积4~9 a林龄间,均呈显著的表型正相关和遗传正相关,且两两林龄间遗传相关系数均大于表型相关系数。林龄相隔越近,遗传相关系数越大,9 a时,树高与5 a以后树高的遗传相关系数达到0.90以上,胸径与5 a以后胸径的遗传相关系数达到0.89以上,单株立木材积与5 a以后的单株立木材积遗传相关系数达到0.91以上。因此,杉木自由授粉子代测定林1~4 a时,需每年调查生长性状,5 a以后,可适当加大调查年度跨度。本试验中5 a时的生长数据能对9 a时的生长数据进行较高的遗传解释,调查年度跨度可达4 a。同时也说明5 a是杉木自由授粉子代测定林开展遗传选择的底限林龄。
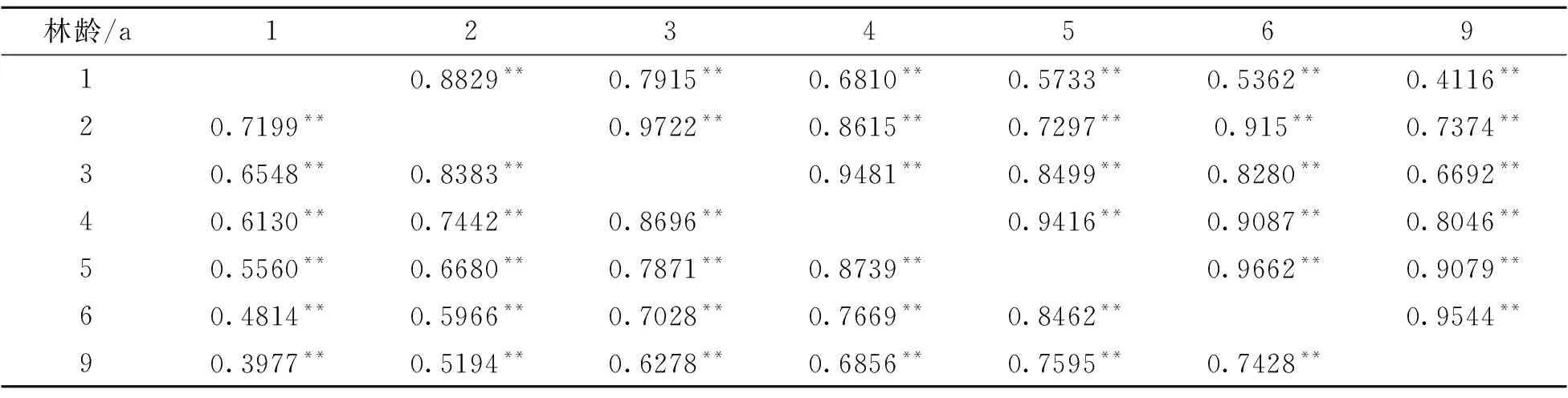
表9 树高早晚表型及遗传相关系数

表10 胸径早晚表型及遗传相关系数

表11 单株立木材积早晚表型及遗传相关系数
试验林9 a时参试家系性状间的表型相关系数和遗传相关系数见表12。树高、胸径、单株立木材积与红心材率呈弱度的表型正相关、强度的遗传正相关;而与木材基本密度呈弱度的表型负相关、中度的遗传负相关。红心材率与木材基本密度表型不相关,但呈强度的遗传负相关。
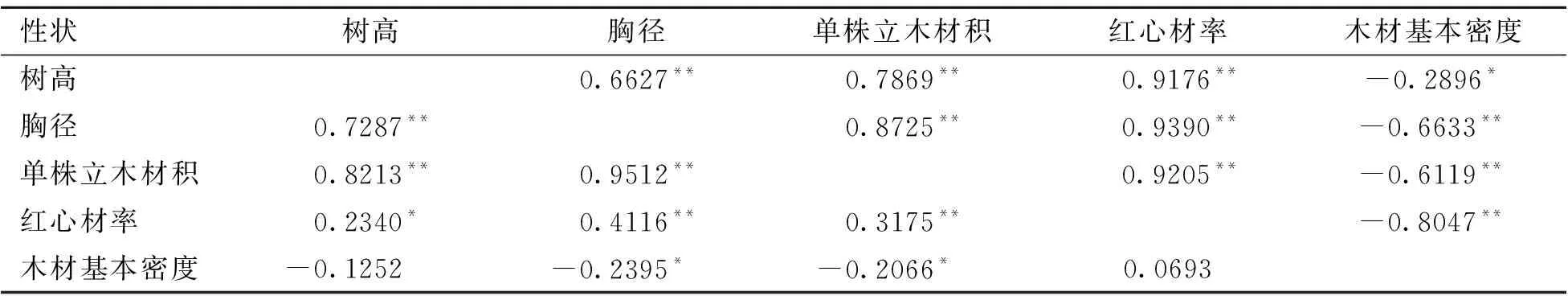
表12 9 a林龄时性状间表型及遗传相关系数
2.4 家系、单株和无性繁殖原株选择
在遗传变异分析及遗传相关性分析基础上,根据9 a时单株立木材积育种值和木材基本密度育种值排名,从参试的30个家系中选择优良家系作为杉木第3.5代种子园建园候选材料、家系林分用种及第3代种子园去劣疏伐依据,从测定木材基本密度的单株中选择优良单株充实杉木第4代种质资源。根据单株立木材积基因型值排名,从试验林中选择无性系繁殖原株,充分利用遗传因素对生长性状的加性和非加性遗传效应,丰富无性繁殖材料。
以单株立木材积和木材基本密度均具有正向遗传增益为标准,筛选出优良家系5个(表13),树高、胸径、单株立木材积、木材基本密度均值分别为10.62 m、14.0 cm、0.09365 m3、0.3292 g·cm-3,遗传增益分别为0.87%、1.38%、3.38%、1.24%。以单株立木材积遗传增益不低于30%,木材基本密度具有正向遗传增益为标准筛选出优良单株12株(表14),树高、胸径、单株立木材积、木材基本密度均值分别为13.03 m、18.9 cm、0.19136 m3、0.3391 g·cm-3,遗传增益分别为5.63%、14.00%、37.77%、2.31%。以单株立木材积遗传增益不低于100%为标准筛选出无性系繁殖原株11株(表15),树高、胸径、单株立木材积均值分别为13.70 m、19.3 cm、0.20843 m3,遗传增益分别为22.27%、34.57%、111.73%。
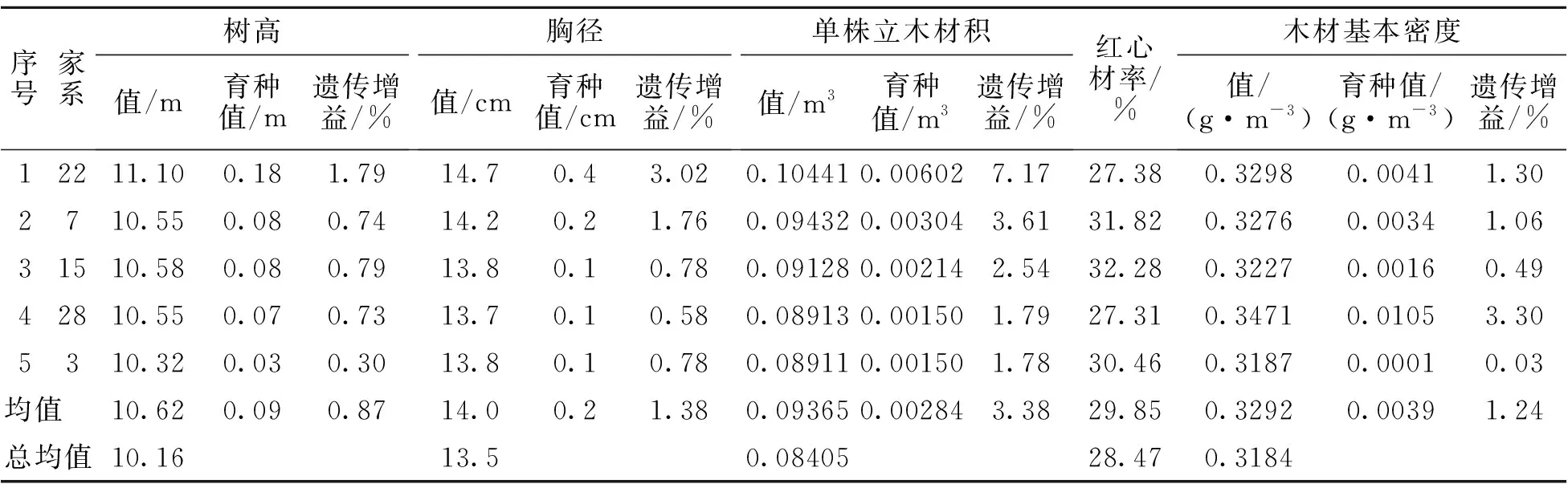
表13 优良家系(9 a)
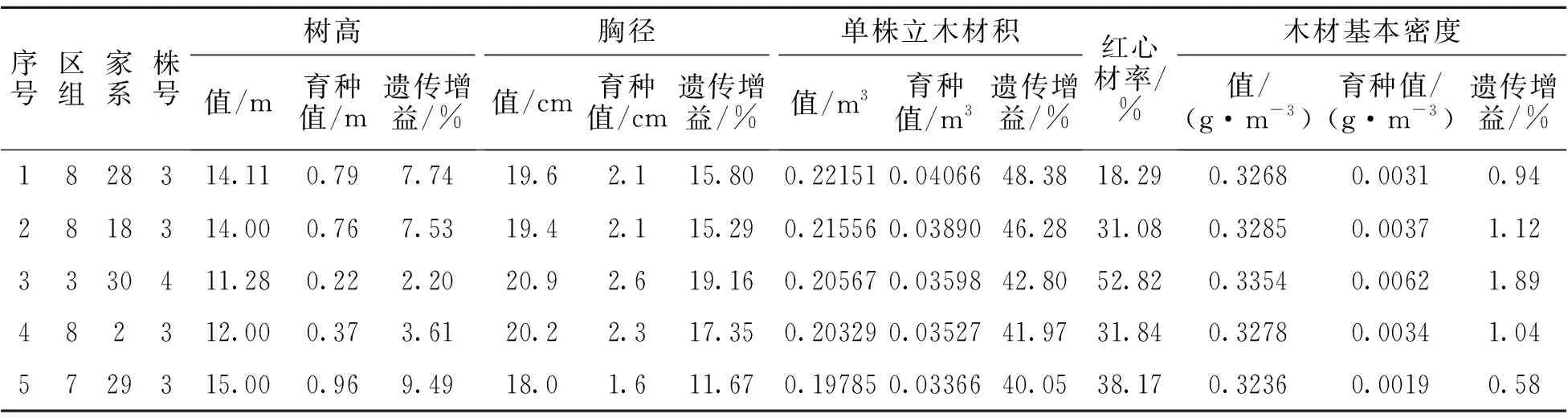
表14 优良单株(9 a)

表14(续)

表15 无性繁殖原株(9 a)
3 结论与讨论
杉木在我国森林乔木树种中的重要值排名第一,是我国最早开展遗传改良研究的树种之一[3],也是改良成效最优的树种之一[5,12]。长期的试验观测结果和实践经验证明,杉木第3代种子园的遗传增益可达76%[17]。在“生态建设、生态安全、生态文明”的林业基本战略要求下[18],以生态林业建设为中心,实现林业现代化是林业工作的主要内容[17]。因此,广泛开展杉木第3代种子园子代测定,优中选优,进一步巩固和发挥杉木第3代育种研究的成果和作用,依靠育种手段稳步提高杉木良种遗传增益,本着“林以种为本,种以质为先”的科学理念,推进杉木人工林高质量发展,充分发挥杉木在我国南方商品材林区的生态、经济和社会效益,是当前杉木人工经营面临的重要课题。福建省洋口国有林场2003年营建的杉木第3代种子园是福建省乃至全国第一个杉木第3代种子园,为福建省全面建设杉木第3代种子园提供了宝贵的建设经验和建园材料,也是福建省最早开展子代测定的杉木第3代种子园。其自由授粉子代测定林9 a时平均树高、胸径、单株立木材积分别为10.16 m、13.48 cm、0.08405 m3,年均生长量分别为1.13 m、1.5 cm、0.00934 m3,家系间表型变异系数分别为3.82%、6.03%、13.54%。
在不同林龄时,树高、胸径和单株立木材积家系间差异均极显著,1~9 a时,树高的广义遗传力介于0.6386~0.7723间,狭义遗传力介于0.1992~0.3609间,4~9 a时,胸径的广义遗传力介于0.7226~0.8006间,狭义遗传力介于0.2991~0.3481间,单株立木材积的广义遗传力介于0.6627~0.7550间,狭义遗传力介于0.2445~0.2958间。9 a时红心材率在家系间差异不显著,不受遗传控制;木材基本密度在家系间差异极显著且广义遗传力为0.4751,狭义遗传力为0.3655。树高、胸径、单株立木材积受到明显的非加性遗传效应影响,木材基本密度主要受到加性遗传效应影响。
9 a时的树高、胸径、单株立木材积与5 a以后的树高、胸径、单株立木材积遗传相关系数分别达到0.90、0.89、0.91以上,据此推断5 a是杉木自由授粉子代测定林遗传选择的底限林龄。树高、胸径、单株立木材积与红心材率呈弱度的表型正相关、强度的遗传正相关;而与木材基本密度呈弱度的表型负相关、中度的遗传负相关。红心材率与木材基本密度表型不相关,但呈强度的遗传负相关。因此,以9 a时单株立木材积、木材基本密度均具有正向遗传增益为标准,筛选出5个优良家系作为杉木第3.5代种子园建园候选材料、家系林分用种及第3代种子园去劣疏伐依据,树高、胸径、单株立木材积、木材基本密度均值分别为10.62 m、14.0 cm、0.09365 m3、0.3292 g·cm-3,遗传增益分别为0.87%、1.38%、3.38%、1.24%。以单株立木材积遗传增益不低于30%,木材基本密度具有正向遗传增益为标准筛选出12株优良单株充实杉木第4代种质资源,树高、胸径、单株立木材积、木材基本密度均值分别为13.03 m、18.9 cm、0.19136 m3、0.3391 g·cm-3,遗传增益分别为5.63%、14.00%、37.77%和2.31%。以单株立木材积遗传增益不低于100%为标准筛选出11株无性系繁殖原株,树高、胸径、单株立木材积均值分别为13.70 m、19.3 cm、0.20843 m3,遗传增益分别为22.27%、34.57%、111.73%。
杉木树高、胸径和单株立木材积等生长性状间一般具有较高的遗传增益,而生长性状与木材基本密度的遗传相关性较为复杂,受到参试群体、测定林龄、立地条件、气候等多因素的影响,有的具有负向遗传相关性,有的没有相关性;即生长性状与木材基本密度的遗传相关性不如生长性状间的遗传相关性稳定,这对优良家系和单株的选择策略具有重要影响。本研究中,生长性状与木材基本密度就具有明显的负向遗传相关性,因此需兼顾生长性状和木材基本密度均具有正向遗传增益进行家系和优良单株(育种材料)的选择,这样选出来的家系是遗传上综合表现最优(即育种值均为正直)的家系和单株,而并不是单一性状最优家系和单株。
