冬小麦对过量钼胁迫的代谢响应与耐钼机制
2021-09-27李启彪李路胡承孝谭启玲孙学成
李启彪,李路,2,胡承孝,谭启玲,孙学成
1.华中农业大学资源与环境学院/微量元素研究中心,武汉 430070;2.西藏自治区农牧科学院农业质量标准与检测研究所,拉萨 850032
钼是对人类、动物和植物正常生长发育至关重要的一种营养元素,其参与合成的钼辅因子在生物代谢活动中起着不可替代的作用,但生物对钼的需求量很低,且从缺乏到适宜的浓度范围非常小[1]。岩石圈中钼的平均含量为2.3 mg/kg,但钼矿的开采活动产生的尾矿与废渣会增加钼在环境中的浓度,并在矿区附近造成土壤、地下水和农田污染[2-5]。现普遍认为当土壤中钼含量高于10 mg/kg即会造成污染,但在我国的葫芦岛市、赤峰市、闽东地区,美国的加利福尼亚州以及加拿大哥伦比亚省等地区的钼矿区已发生的钼污染都远超该限值,如我国葫芦岛矿区周边果园土壤的平均钼含量高达720.61 mg/kg[6-10]。
大量研究表明,过量的钼会使植物受到氧化胁迫进而影响生长和产量[11-12],会扰乱类囊体排列并抑制光合作用[13],还会引起其他矿质营养元素如Mn、Mg的缺乏等症状[11]。同时许多植物对高浓度的钼有着较强的耐受能力,即使在受到钼污染的环境下也没有表现出毒性性状。据报道,黑麦草、玉米即使在含有1 000 mg/kg钼的土壤中也能正常生长[14],白菜幼苗暴露在10 mmol/L的钼酸钠下时叶绿素含量也没有明显变化[15],而小麦在土壤钼含量高达1 000 mg/kg下长势与产量也不受明显影响,说明小麦也拥有较强的耐钼能力[16-17]。
小麦作为世界产量最高、分布最广的粮食作物,部分小麦产区已有土壤钼污染的报道,如我国陕西省洛南县黄龙浦钼矿区的小麦农田土壤钼含量可达193 mg/kg[18],但目前过量钼胁迫对小麦生长的影响及其机制尚未明确。本研究以小麦作为试验材料,研究过量钼在小麦中的亚细胞分布并结合LC-MS代谢组学分析钼胁迫下的差异代谢物,探明小麦对土壤钼污染的代谢响应和耐受机制,以期为植物耐受过量钼胁迫的机制提供理论支持,并为植物修复土壤钼污染提供技术参考。
1 材料与方法
1.1 试验材料
供试材料为华中农业大学微量元素研究中心自繁留种的冬小麦品种97014,在前期的土壤盆栽试验中发现小麦在钼含量0.15 mg/kg至1 000 mg/kg的高钼环境下也能保持产量与长势未显著下降[16],因此设置了正常钼水平(1 μmol/L)和过量钼水平(1 mmol/L)2个处理,以钼酸铵 (NH4)6Mo7O24·4H2O为肥源,作4次重复。
采用营养液培养小麦,其中大量元素采用的Hogland营养液含有4 mmol/L Ca (NO3)2·4H2O、6 mmol/L KNO3、1 mmol/L NH4H2PO4、2 mmol/L MgSO4·7H2O;微量元素采用的无钼Arnon营养液含有100 μmol/L EDTA-Fe、46.2 μmol/L H3BO3、9.1 μmol/L MnCl2·4H2O、0.8 μmol/L ZnSO4·7H2O和0.3 μmol/L CuSO4·5H2O,营养液均使用去离子水配制(电阻率>18.25 MΩ·cm,25 ℃)。
所用试剂均为分析纯级别,试验所有器皿均用2 mol/L HNO3浸泡48 h,再用去离子水冲洗干净。质谱级乙腈、色谱级甲醇均购于Merck (Dannstadt,Gemany),甲酸购于CNW(Duesseldorf,Germany),其余试剂均为分析纯,过程所用水均为去离子水。
1.2 培养方案
小麦种子用0.5%次氯酸钠溶液消毒30 min后用去离子水冲洗干净,播于湿润的医用纱布上25 ℃恒温催芽。待芽长约1 cm时,于2015年10月18日选择大小相对一致的小麦幼苗25株,移植于盛有10 L营养液的方盒并放置在人工气候室中,人工气候室条件为20 ℃/18 ℃(16 h/8 h),光照强度为100 μmol/(m2·s)。首先将小麦幼苗用1/2浓度营养液培养5 d,再用全量浓度营养液培养,每5 d更换1次营养液。正常钼水平(1 μmol/L)培养15 d后,设正常钼水平(1 μmol/L)和过量钼水平(1 mmol/L)2个处理,继续培养10 d后收获测定地上部、地下部及其亚细胞的钼含量,并对叶片进行代谢组学分析。
1.3 钼含量的测定方法
小麦亚细胞钼含量的测定在Liu等[19]的方法基础上进行修改,称取0.5 g鲜样于用液氮预冷的研钵中,用5 mL提取液将其研磨成匀浆,提取液使用1 mmol/L二硫赤藓糖醇、50 mmol/L Tris-HCl、5.0 mmol/L抗坏血酸、250 mmol/L蔗糖和10 g/L聚乙烯吡络烷酮(PVP),并调节pH值至7.5。使用240 μm的尼龙网过滤所得残渣为细胞壁部分,将上清液在4 448 r/min下离心20 min,所得残渣为地上部的叶绿体或是地下部的原生质体,将后续上清液在10 896 r/min下继续离心35 min,所得残渣为细胞膜以及细胞器组分,而最后的上清液为可溶性部分,该过程所有步骤均保持在4 ℃下进行。将各亚细胞组分与混酸(HNO3∶HClO4=4∶1,V/V)混合后在180~230 ℃下消化,用去离子水转移定容并过滤后使用石墨炉(日立,Z-2000)进行钼含量的测定。
1.4 代谢产物的测定方法
本试验代谢产物的测定采用LC-MS(Ultimate 3000LC,Thermo)平台,色谱柱为Hypergod C18 (100 mm×4.6 mm,3 μm)。色谱分离条件设置为:流速0.3 mL/min、柱温40 ℃;流动相组成为A:水+0.1%甲酸,B:乙腈+0.1%甲酸;流动相的梯度洗脱程序见表1。进样量为4 μL,自动进样器的温度为4 ℃。
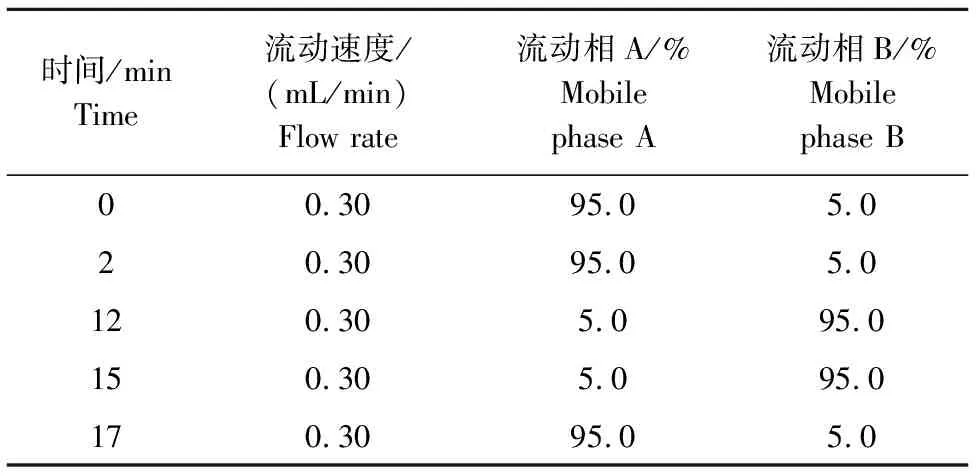
表1 流动相梯度 Table 1 The gradient of mobile phase
质谱参数:正模式,加热器温度300 ℃,辅助气流速15 mL/min,扫描气体流速1 mL/min,鞘气流速45 mL/min,喷雾电压3.0 kV,毛细管温度 350 ℃,透镜电压水平30%;负模式,加热器温度300 ℃,辅助气流速15 mL/min,扫描气体流速1 mL/min,鞘气(N2)流速45 mL/min,喷雾电压3.2 kV,毛细管温度350 ℃,透镜电压水平60%。
1.5 数据处理
使用Microsoft Excel 2010进行数据整理计算,Origin 2018绘图,SPSS 20.0进行方差分析,各处理平均值的多重比较釆用独立样本t检验(P<0.05)。
LC-MS数据使用SIEVE软件进行预处理后使用Microsoft Excel 2010 软件进行归一化处理和后期编辑,并将最终数据组织成二维数据矩阵,其中包含有变量(rt_mz)、相对分子质量(Comp MW)、观察量(样本)和峰强。此项目样本在正模式下共获得2 146个特征峰,在负模式下共得到1 394个特征峰,将编辑完成的数据矩阵导入SIMCA-P(13.0)软件进行后续分析,将得到的差异代谢物导入MetaboAnalyst 3.0以获得差异代谢物参与的代谢通路。
2 结果与分析
2.1 钼污染对冬小麦亚细胞中钼含量的影响
为了解正常与过量钼水平下冬小麦各个亚细胞的钼含量及分布,对根系及地上部样品进行了亚细胞分级,结果表明,正常钼水平(1 μmol/L)下小麦根系亚细胞钼含量表现为可溶性部分>细胞膜+细胞器>原生质体>细胞壁;过量钼(1 mmol/L)水平下则表现为可溶性部分>细胞壁>原生质体>细胞膜+细胞器。2个钼水平下钼在可溶性部分中的含量及分配比例均为最高(图1)。
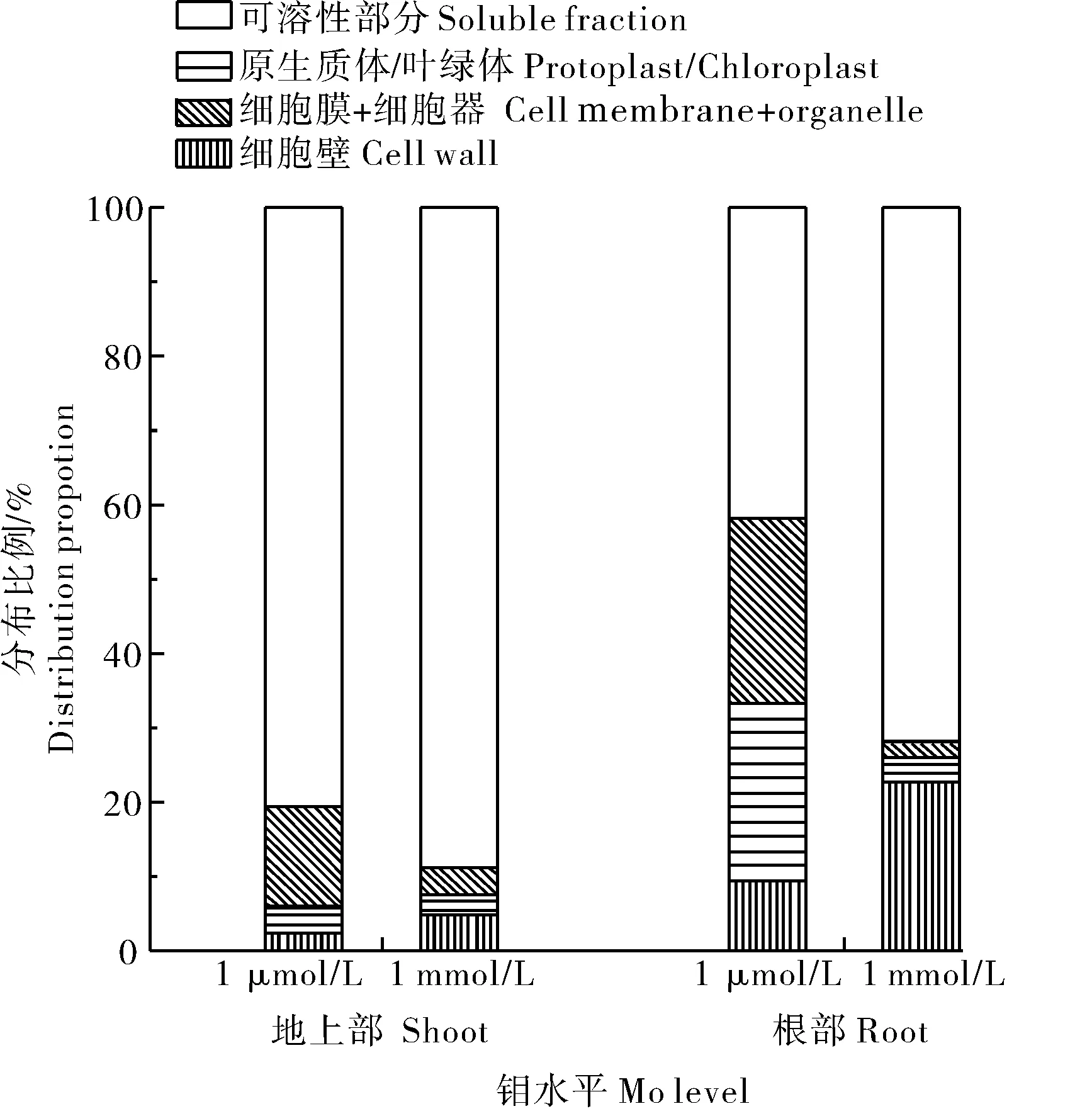
图1 冬小麦亚细胞中钼的分布Fig.1 Subcellular distribution of molybdenum in winter wheat
由表2可知,当培养介质中的钼含量从1 μmol/L上升到1 mmol/L时,根部可溶性部分、细胞壁、细胞膜+细胞器及原生质体中的钼含量分别上升了1 300、1 815、62、105倍,其中可溶性部分的钼含量最高,达533.0 mg/kg;从钼在各亚细胞组分所占比例来看,在钼过量条件下,钼在细胞壁和可溶性部分分配比例分别增加了13.3%和30.2%,总体占比可达22.8%和71.8%,可见过量钼水平下小麦根系中绝大部分的钼存储在液泡和细胞壁中,而液泡则是最主要的储存部位。
小麦地上部亚细胞钼含量在正常钼水平(1 μmol/L)下表现为:可溶性部分>细胞膜+细胞器>叶绿体>细胞壁;过量钼水平(1 mmol/L)下表现为:可溶性部分>细胞壁>细胞膜+细胞器>叶绿体;2个钼水平下钼在可溶性部分中的含量均为最高。当介质(营养液)中的钼含量从1 μmol/L上升到1 mmol/L时,可溶性部分、细胞壁、细胞膜+细胞器及叶绿体中的钼含量分别上升了19、35、5、12倍,也是可溶性部分中钼含量最高,达13.76 mg/kg;从钼在各亚细胞组分所占比例来看,在过量钼水平下,钼在可溶性部分的分配比例增加了8.0%,总体占比高达88.5%。由此可知,在2个钼水平下钼小麦地上部吸收的钼均主要存储在液泡中,其中过量钼水平下在液泡中的分配比例要更高。
2.2 过量钼对冬小麦叶片代谢物的影响
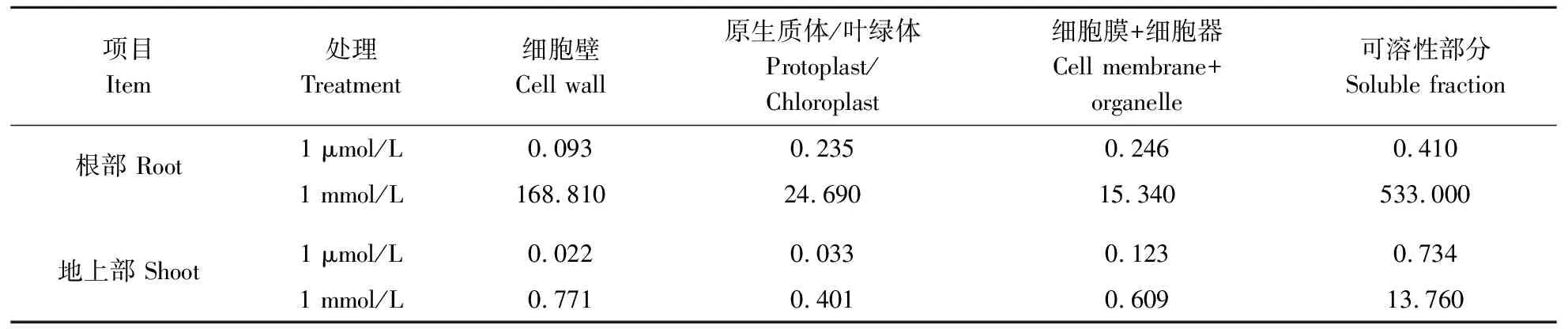
表2 冬小麦亚细胞中的钼含量 Table 2 Subcellular Mo content in winter wheet mg/kg
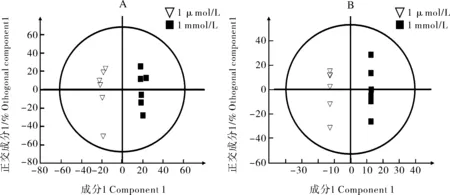
A:正模式;B:负模式。A:Positive mode; B:Negative mode.图2 正常钼和过量钼两组处理的OPLS-DA得分图Fig.2 Score plots of OPLS-DA of normal molybdenum and excess molybdenum treatment
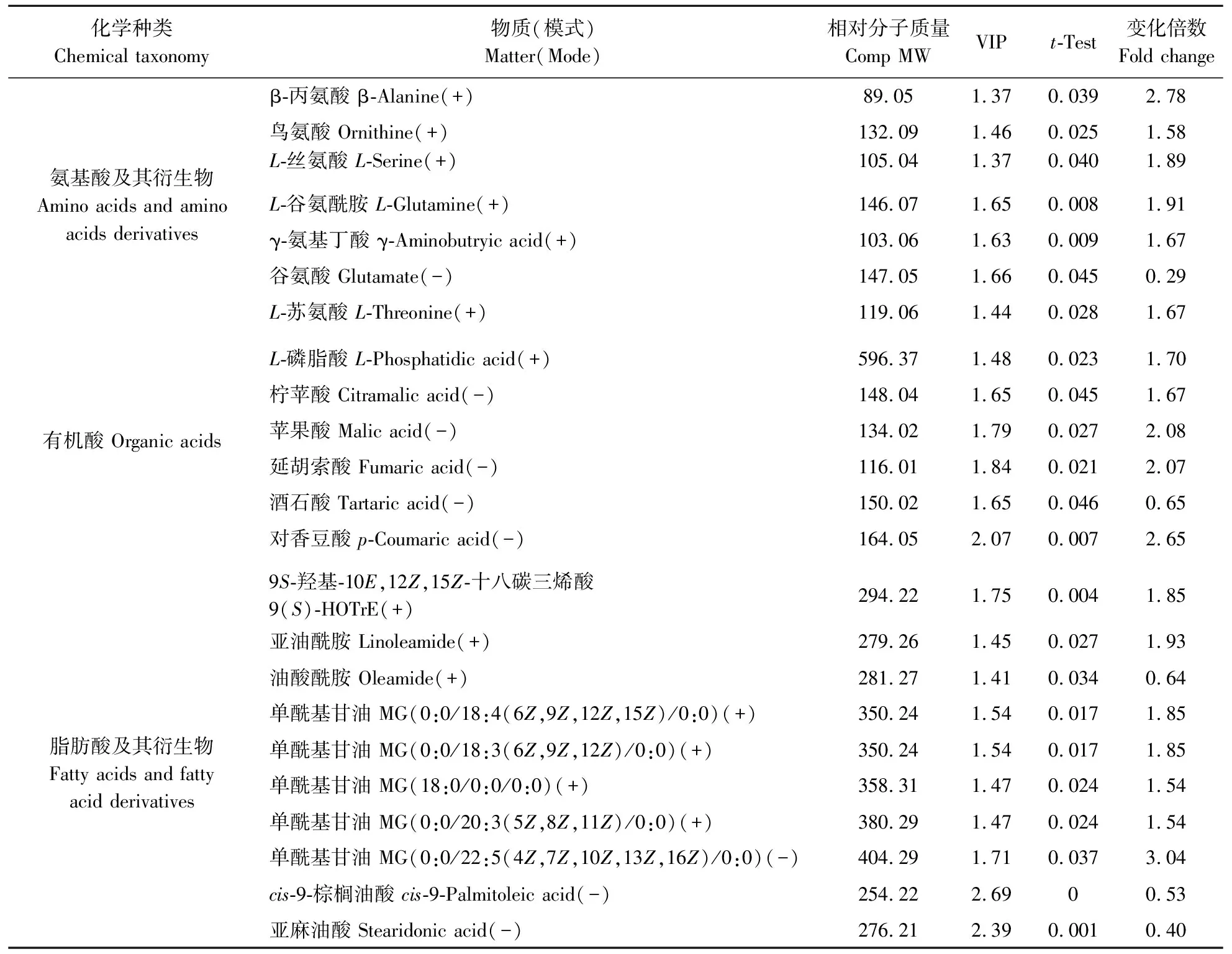
表3 过量钼下的冬小麦的差异代谢物 Table 3 Discriminatory metabolites of winter wheat under excess-Mo
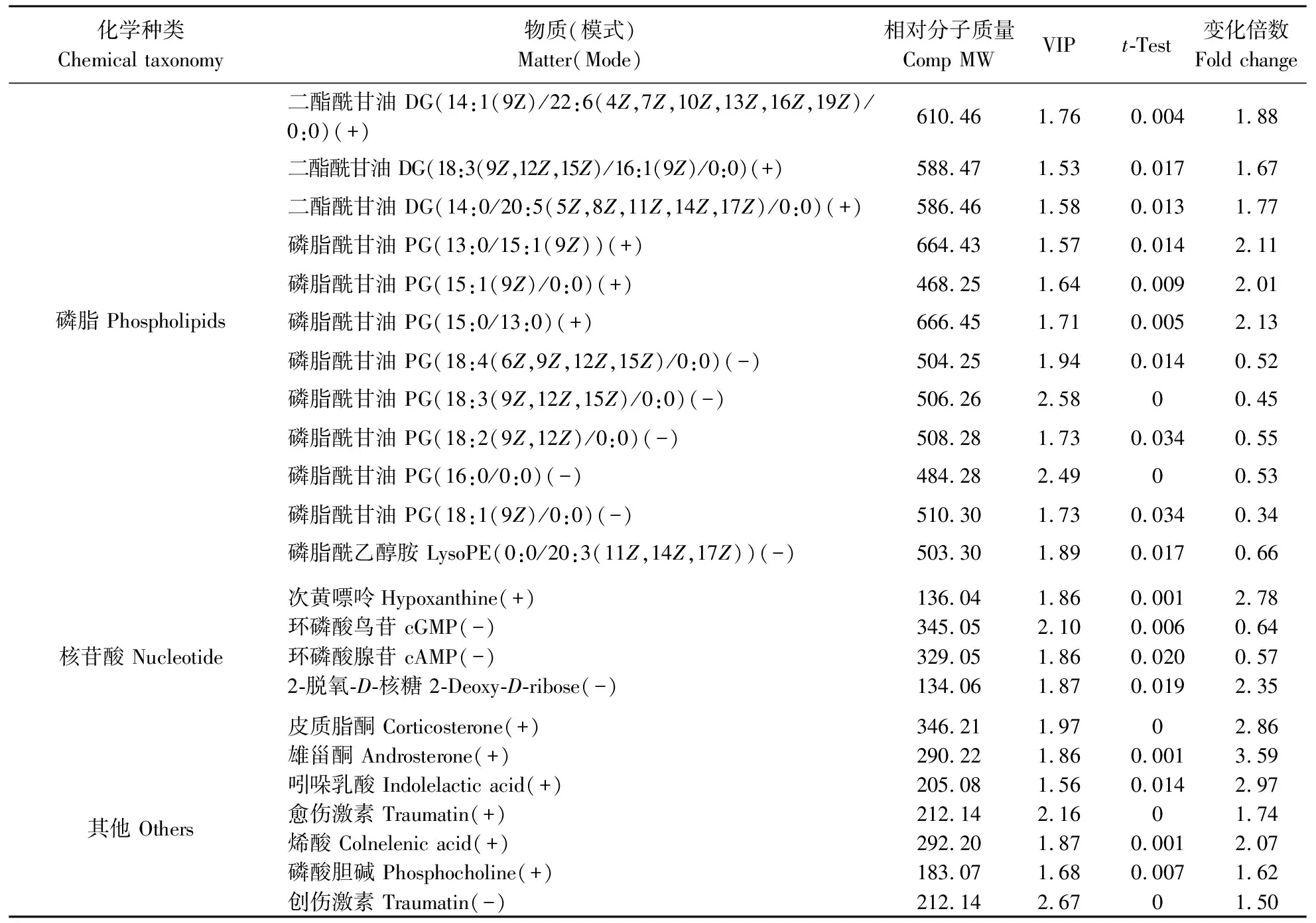
续表3 Continued Table 3
正常钼与过量钼小麦叶片共鉴定出48个差异代谢物,其中有34个代谢物上调,14个代谢物下调。代谢物中大部分的氨基酸、有机酸、脂肪酸及其衍生物均有显著上调,而磷脂和核苷酸类代谢物则是部分下调,在34种上调代谢物中包含6种氨基酸及其衍生物、5种有机酸。将得到的差异代谢物导入MetaboAnalyst 3.0软件后得到这些差异代谢物所处的代谢通路(图3),可发现过量钼促进了冬小麦叶片中氨基酸的合成与代谢过程,并促进了TCA循环中有机酸的合成过程。
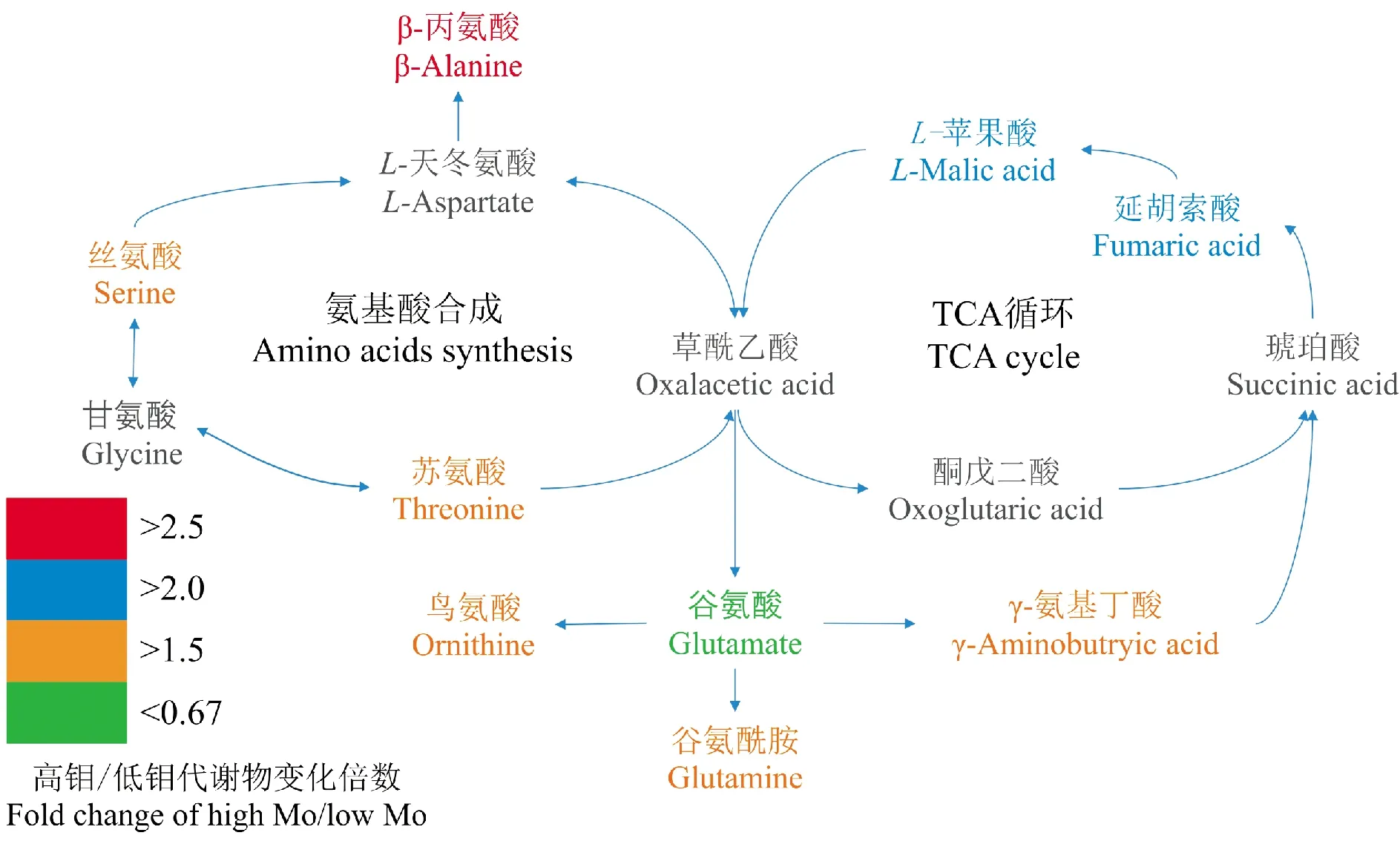
图3 过量钼胁迫下的冬小麦叶片的代谢通路分析Fig.3 Discriminatory metabolites of winter wheat under excess-Mo
3 讨 论
植物对钼的耐性较强,如小麦、玉米在钼含量1 000 mg/kg的土壤中生长,叶片中的钼含量分别可达4 000 mg/kg和400 mg/kg,而作物生长也未表现出显著抑制[15-17,20]。较早的研究发现,在钼污染环境中,植物根系钼含量通常远高于地上部,如在土壤钼含量1 000 mg/kg时,小麦根部钼含量是地上部的3倍,而在玉米、大豆、黑麦草等植物上也能观察到相同的趋势[14-16]。本试验基于前期土壤盆栽试验中发现小麦在钼添加量高达1 000 mg/kg的环境下也能维持正常产量的结果[16],设置了100 mg/L的过量钼水培试验以探究小麦的耐钼机制,结果表明在过量钼水平下,过量钼处理大幅增加了小麦根系亚细胞组分钼含量,细胞壁、可溶性部分、细胞器、原生质体/叶绿体的钼含量分别是地上部各亚细胞组分的219、38、25和62倍。前人在大豆、玉米、油菜等植物上的研究也表明,高钼介质中生长的作物根系钼含量分别能达到地上部钼含量的49、3和5倍[15,21-22]。这些结果说明小麦根部在过量钼环境下能够吸收并积累更多的钼,这是阻止过量钼向地上部迁移的第一道屏障。
将过量金属进行亚细胞区隔化以降低游离过量金属离子是植物重要的耐受机制之一,一些植物会将吸收的过量金属区隔在细胞壁中,如茶树能在过量铝环境下将69.8%的铝固定在细胞壁中,并将其与果胶和半纤维素结合[23];而液泡则是植物区隔金属离子的另一个重要部位,植物在吸收过量金属离子后能将其与氨基酸、有机酸和植物螯合素结合后转运储存在细胞的液泡中[24],如Vögeli-Lange等[25]报道了暴露在高镉环境下烟草细胞中的镉能与植物螯合素络合后转运至液泡中。本研究发现,正常钼水平下的钼在冬小麦根系及地上部的可溶性部分中分配比例分别为41.6%和80.5%,当钼水平从正常(1 μmol/L)上升到过量(1 mmol/L)时,钼在根系及地上部的可溶性部分分配比例分别上升了30.2和8.0个百分点,达到71.8%和88.5%,说明无论是在正常钼水平还是过量钼水平下,液泡都是冬小麦中钼的主要储存部位,且过量钼水平下钼在液泡中的储存比例更大。Xu等[26]在大豆上的研究结果也显示,当营养液质量浓度为1 mg/L时,钼在根系及地上部可溶性部分中的分配比例为26.4%~46.3%,当营养液钼质量浓度从1 mg/L上升到100 mg/L时,钼在大豆根系及叶片可溶性部分中储存比例达到66.3%~72.2%。Qin等[21]在油菜上的研究结果表明,当营养液中钼浓度从1 μmol/L上升到200 μmol/L时,钼在根系液泡中的分配比例从47.3%上升到59.5%,在叶片液泡中的分配比例从41.2%上升到62.8%。综上得出结论,小麦在面对过量钼胁迫时能将吸收的过量钼固定在根部以防止其向地上部运输,并把大部分的钼转运储存在根部的液泡中,相比于其他作物,小麦要更依赖于液泡对过量钼的区隔作用。
液泡是细胞中各种有机酸、氨基酸及各种金属离子的存储中心,氨基酸、有机酸、植物螯合素等有机物通常会在液泡中与游离金属离子发生螯合作用,这是植物耐受重金属的另一个重要机制[27]。代谢组分析结果表明,在钼胁迫下冬小麦中氨基酸的合成代谢和TCA循环过程被显著促进,参与该过程的苹果酸、延胡索酸含量分别上升了2.08倍和2.07倍,而许多有机酸如柠檬酸、苹果酸、延胡索酸、草酸等能够和Mo、Ni、Cd等金属离子螯合[28-31],另外本研究还发现过量钼处理使小麦的β-丙氨酸、L-丝氨酸、L-苏氨酸、L-谷氨酰胺的含量分别上升了2.78、1.89、1.67和1.91倍,已有大量研究表明植物体内的脯氨酸、谷氨酰胺等氨基酸能螯合Zn、Cd等金属离子[27,32-33];还有报道表明,钼与L-丝氨酸、L-丙氨酸、L-苏氨酸、L-天冬氨酸等氨基酸在体外反应体系下能形成配合物[34]。因此,我们推测小麦在过量钼环境下增加合成的延胡索酸、苹果酸、L-丝氨酸、L-丙氨酸和L-苏氨酸能在液泡中与钼酸根离子反应形成络合物,从而降低过量钼对小麦细胞的毒害。
在过量钼环境下,小麦亚细胞中的钼主要分布于细胞壁和可溶性部分,且根部的液泡在过量钼的固持过程中起主要作用,过量钼促进了小麦氨基酸合成代谢与TCA循环过程,该路径中多种氨基酸(β-丙氨酸、L-丝氨酸、L-苏氨酸、L-谷氨酰胺)和有机酸(苹果酸、延胡索酸)的含量都显著增加,推测这些氨基酸与有机酸能与钼进行络合并固定在液泡中,从而增强小麦对过量钼的耐受能力。
