灌浆期不同株龄老芒麦旗叶对光照强度的响应及光合基础环境指标差异
2021-09-25金鑫祁娟刘文辉吴召林杨航宿敬龙李明
金鑫,祁娟,刘文辉,吴召林,杨航,宿敬龙,李明
(1.甘肃农业大学草业学院,草业生态系统教育部重点实验室,甘肃省草业工程实验室,中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070;2.青海省畜牧兽医科学院,青海省青藏高原优良牧草种质资源利用重点实验室,青海 西宁 810016)
老芒麦(Elymussibiricus)为禾本科披碱草属植物,原产于我国高原地区,凭借其抗寒、抗冻和高产等特性,对高寒生境有极强的适应性,成为了青藏高原天然草地和高寒牧区人工草地的优势草种[1-2]。在青藏高原地区草地退化生态系统中,为解决草地生产力不足和草地生态恶化问题,采取大力发展人工草地的措施,其中老芒麦有着重要作用。但近年研究发现,高寒区人工披碱草草地在第4年呈现由生产稳定性急剧向生态稳定性转化,即明显退化趋势[3]。亦有研究发现老芒麦最佳利用年限为2~4年,此时产量也最大,随后出现下降趋势[4]。这些因素限制了多年生牧草在高寒天然草地改良与人工草地可持续发展中的应用与推广。
目前,关于密度[5]、施肥[6]、添加外源激素[7]等栽培措施对老芒麦生殖枝数、种子和草产量及营养品质的研究较多[8]。光合作用贯穿植物整个生命活动,也是老芒麦生产力的关键生理基础。老芒麦的光合性能直接或间接地影响着物质的合成、积累和分配。关于老芒麦光合生理的研究集中在光合特性[9]、光合日变化[10]、行距与光合的关系[11]及不同叶位叶片的光合特性[12]等。对白芷(Angelicadahurica)[13]、甘草(Glycyrrhizauralensis)[14]、紫花苜蓿(Medicagosativa)[15]等植物的研究表明,不同生长年限的植物叶片光合特性存在差异且不同年限对光强敏感性也不同。但在高寒区同一生境下不同生长年限的老芒麦光合特性的差异鲜见报道。鉴于此,本研究选取高寒区同一生境下3、4、5株龄的青牧1号老芒麦为研究对象,通过模拟有效光辐射,测定不同株龄老芒麦旗叶的光合参数差异,分析老芒麦株龄对叶片光响应能力的影响;同时通过测定不同株龄老芒麦株高和叶片形态参数及SPAD值,分析各株龄老芒麦叶片的基础光合环境差异,探讨高寒区不同株龄老芒麦对环境光合特性的响应,旨在从光合生理角度为延长高寒区人工建植老芒麦利用年限提供理论依据。
1 材料和方法
1.1 试验地概况
试验地位于青海省海北州国家草品种区域试验站(海晏),坐标为E 100°85′,N 36°45′,平均海拔3 154 m,属高原大陆性气候。寒冷期长,无绝对无霜期,年均温0.6℃;年降水量为369~403 mm,集中于7、8和9月。土壤类型为栗钙土,该试验区土壤理化性质[16]如表1所示:

表1 试验地土壤理化性状
1.2 试验材料与设计
于2019年选取种植年限分别为3 a、4 a和5 a的“青牧1号”老芒麦人工草地为研究对象,小区设置3次重复,每个小区面积为4 m×5 m,播种深度3 cm,行距30 cm、条播,播种量为45 kg/hm2,仅播种当年施基肥,其余生长年份均不施肥,自然降水无灌溉,每年定期人工清除杂草,并在种子完熟前收割种子。
各处理依次为:1)3 a,人工建植3 a的老芒麦草地,2017年建植;2)4 a,人工建植4 a的老芒麦草地,2016年建植;3)5 a,人工建植5 a的老芒麦草地,2015年建植。
由于不同株龄老芒麦样地内个体发育存在差异,且不同株龄老芒麦达到同一生育时期的时间也不一致。因此以老芒麦样地70%植株处在同一生育时期为标准,计有该样地生育时期。于7月23~8月20日分别在3、4、5株龄老芒麦样地进入灌浆期时,选择晴朗无风的早晨于9∶00-11∶00,利用LI-6400便携式光合仪测定不同生长年限老芒麦叶片的气体交换参数。每小区分别选择10株健康、光照均匀且长势一致的老芒麦旗叶进行测定,设置光子量照度为0,50,200,600,800,1 000,1 200,1 400,1 600,1 800,2 000 μmol/(m2·s)共11个内置光源,测旗叶的净光合速率Pn(net photosynthetic rate)、蒸腾速率Tr(transpiration rate)、气孔导度Gs(stomatal conductance)和胞间CO2浓度Ci(intercellular CO2concentration)等光合参数,并计算气孔限制值Ls(stomatal limit value),Ls=1-Ci/Ca(Ca为大气中CO2浓度),水分利用率WUE(water use efficiency),WUE=Pn/Tr。待光合参数测定完成后,继续测定该叶片的光合基础环境参数,包括株高、叶长、叶宽、叶面积及SPAD值。测定SPAD值时,利用手持式SPAD-502?Plus(原产自日本),每片叶片中部随机选取3点,每点重复3次,求平均值作为该叶片的SPAD值。
1.3 测定指标和方法
株高:用卷尺测定根茎基部至离生长点最近的展开叶顶端的绝对高度为株高。
叶片性状:用直尺测量旗叶的叶长、叶宽(最宽处),单叶面积=叶长×叶宽×叶面积系数(0.75)[17]。

P单表示单位叶面积光合速率,P测为仪器叶室宽度,W表示叶片宽度[18]。
1.4 数据处理
利用Microsoft Excel 2016和R整理数据并绘制图表;用SPSS 19.0对光合参数进行交互双因素方差分析和单因素ANOVA分析处理,LSD法进行多重比较(P<0.05)。以平均值±标准误表示。
2 结果与分析
2.1 不同株龄老芒麦叶片的光合基础环境指标差异
老芒麦株高随着生长年限延长显著降低(P<0.05),4 a和5 a老芒麦较3 a分别降低10.54%和24.50%;4 a叶长显著增大(P<0.05),3 a和5 a间无显著差异;4 a叶宽最大,达到0.87 cm,较3 a和5 a分别增加了19.18%和40.32%,且各株龄叶片间差异显著(P<0.05);4 a叶片的叶面积与3 a或5 a差异显著(P<0.05),3 a和5 a无显著差异;各株龄叶片间SPAD值无显著差异(P<0.05)(表2)。

表2 不同株龄老芒麦叶片的光合基础环境指标多重比较
2.2 老芒麦叶片光合参数随株龄和光照强度的变化响应
2.2.1 净光合速率(Pn)和蒸腾速率(Tr) 不同株龄老芒麦叶片Pn对模拟光强变化的响应过程可区分为4个不同阶段:1)在PAR<200 μmol/(m2·s)条件下,Pn随光子量照度升高而线性增高(图1-A);2)Pn随光子量照度升高而缓慢增高;3)在不同光子量照度下,均出现Pn下降;4)Pn随光子量照度进一步升高均再次增高。在0~200 μmol/(m2·s)的光子量照度下,不同株龄老芒麦叶片Pn均对光照响应敏感。在800~1 200 μmol/(m2·s)光子量照度下,5 a叶片Pn出现降低趋势,同样3、4 a叶片分别在1 200~1 400和1 000~1 400 μmol/(m2·s)光子量照度下存在一致现象,说明有光抑制现象,不同株龄叶片间差异表现为Pn初次降低时,高株龄的叶片较低龄光子量照度更小,且达到Pn最低点时,3 a和4 a叶片光子量照度均在1 400 μmol/(m2·s),5 a叶片在1 200 μmol/(m2·s)。总体上,0~200 μmol/(m2·s)的光子量照度,4 a老芒麦叶片的净光合速率较高,200~2 000 μmol/(m2·s)光子量照度,不同株龄老芒麦叶片净光合速率大小顺序为3 a>4 a>5 a。各株龄老芒麦叶片Tr随光照强度的变化趋势与Pn一致(图1-B),且在200~2 000 μmol/(m2·s)光子量照度,Tr值由高到低顺序也相同。
2.2.2 气孔导度(Gs)和胞间CO2浓度(Ci) 不同株龄老芒麦叶片的Gs对光照强度的响应差异明显,可区分为4个阶段(图1-C),变化趋势与Pn对光照强度的响应相一致:均有光强敏感期,缓慢增加期,光抑制期和再次增加期。总体上,0~200 μmol/(m2·s)的光子量照度,老芒麦株龄越高,其叶片的Gs对光照响应越敏感,200~2 000 μmol/(m2·s)光子量照度,不同株龄老芒麦叶片的Gs,大小顺序和Pn相一致。相同株龄老芒麦叶片随着光照强度的增加,总体呈现随Pn增高,Ci降低的趋势,但并非完全呈相反对应关系(图1-D)。首先光子量照度在0~200 μmol/(m2·s)时光合作用增强,CO2消耗增大,导致Ci急剧下降,其次,随光照强度增大,Pn缓慢增高,3 a和5 a叶片Ci呈现增高趋势,4 a叶片Ci存在波动,再者,随光强增大,均出现光抑制时,Pn降低,同时3 a和4a 叶片Ci缓慢降低,而5 a叶片Ci增高,最后,随光照强度增大而光合作用增强时,Ci再次降低。总体上,0~2 000 μmol/(m2·s)的光子量照度内,不同株龄老芒麦叶片的Ci,大小顺序为4 a>5 a>3 a。
2.2.3 气孔限制值(Ls)和水分利用率(WUE) 不同株龄老芒麦叶片的Ls对光照强度的响应各有差异,总体趋势与Ci相反(图1-E )。光子量照度<200 μmol/(m2·s)时,气孔导度增大,光合作用增强,Ci降低,对应Ls增大;当各株龄叶片出现光抑制时,Pn和Gs均降低,3 a和4 a叶片,Ci降低而Ls增高,5 aCi增高而Ls降低。在不同光照强度范围内,各株龄老芒麦的叶片水分利用率亦存在差异,在200~2 000 μmol/(m2·s)光子量照度内,4 a叶片WUE较高,在600~800 μmol/(m2·s)光子量照度内,5 a的叶片WUE较高,3 a叶片在800~2 000 μmol/(m2·s)光子量照度下WUE较高(图1-F)。

图1 不同株龄和光强梯度下的光合参数Fig.1 Effects of plant age and light intensity on photosynthetic characteristics
2.3 不同株龄老芒麦叶片光合因子间相关性分析
模拟光照强度下,3个不同株龄老芒麦的叶片各光合因子相关分析(图2)表明,净光合速率与气孔导度呈极显著正相关(P<0.01),相关系数3 a>4 a和5 a;蒸腾速率与胞间CO2相关性不同株龄间存在差异,3 a>4 a且极显著负相关,5 a不显著;气孔导度与水分利用率相关性不同株龄间存在差异,3 a>4 a且极显著正相关,5 a不显著;胞间CO2与净光合速率、蒸腾速率和气孔导度相关性存在差异:Ci与Pn,3个株龄均呈极显著负相关;Ci与Tr,3 a>4 a且极显著负相关,5 a不显著;Ci与Gs,3 a和4 a呈极显著负相关,5 a不显著。气孔限制值与净光合速率呈极显著正相关,相关系数3 a>5 a>4 a;气孔限制值与胞间CO2呈极显著负相关,相关系数3 a和5 a>4 a;水分利用率和气孔限制值呈显著负相关(P<0.05)。

图2 各株龄老芒麦叶片光合因子之间相关性分析Fig.2 Coefficient of correlation among Pn,Tr,Gs,Ci,Ls and WUE of the leaves in E.sibiricus with different plant age注:***相关显著性水平为0.001;**相关显著性水平为0.01;*相关显著性水平为0.05
2.4 株龄和光照对老芒麦叶片光合参数的影响
不同株龄老芒麦的叶片与光照强度共同作用对部分光合参数有显著影响(P<0.05),影响大小顺序为Pn>Gs(表3)。进一步分析表明,光照强度对光合参数均存有极显著影响(P<0.01),影响顺序为Gs>Pn>Tr>Ci;株龄对部分光合参数有极显著影响(P<0.01),主要为Ci。可见,光照强度主要影响气孔导度和净光合速率,株龄主要影响胞间CO2浓度,光照强度和株龄共同作用时,主要影响净光合速率和气孔导度。
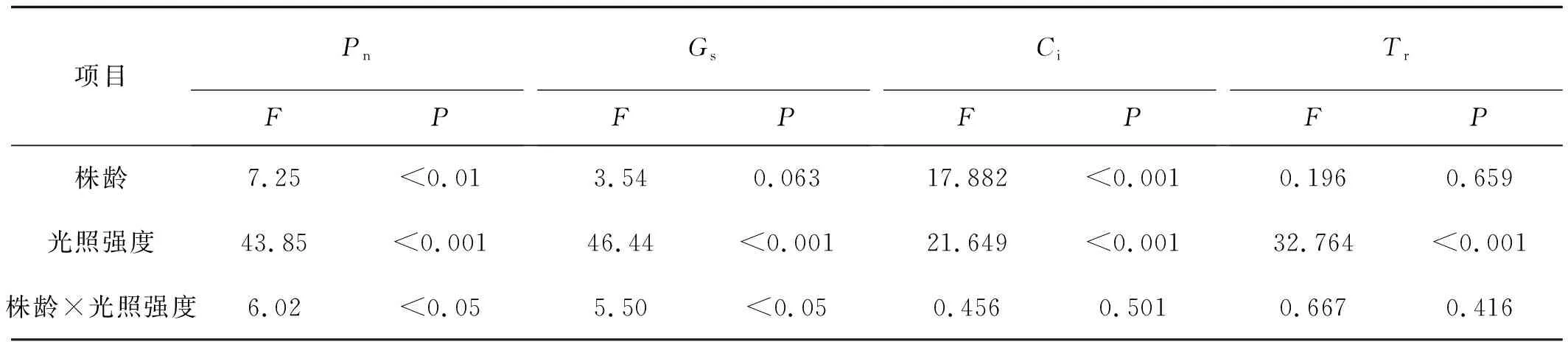
表3 不同株龄老芒麦叶片光合参数的重复试验双因素方差分析
2.5 光照强度对老芒麦叶片光合参数的影响
对0~2 000 μmol/(m2·s)光子量照度下不同株龄的叶片光合参数进行比较(表4),老芒麦叶片能够利用50 μmol/(m2·s)的光进行光合作用,光子量照度在1 000~1 400 μmol/(m2·s)光照下,老芒麦的Pn、Gs、Ci等光合参数均无显著(P<0.05)差异,Tr和Ls差异不显著。光照在1 600~1 800 μmol/(m2·s)时,Pn、Ci、Ls无显著差异,Gs和Tr差异不显著。光照强度在1 800~2 000 μmol/(m2·s)时,老芒麦叶片Ls无显著差异,Pn和Ci差异不显著,Gs和Tr差异显著(P<0.05)。光子量照强度在800 μmol/(m2·s)时,老芒麦叶片水分利用效率高于其他光子量照度。

表4 不同光子量照度下老芒麦叶片的光合参数
2.6 株龄对老芒麦叶片光合参数的影响
为了反映不同株龄老芒麦叶片在一昼夜的光合作用差异,采用模拟光子量照度在0~2 000 μmol/(m2·s)。株龄对老芒麦旗叶的多重比较表明(表5),3、4和5 a间相邻的叶片Pn和Tr差异不显著,3 a和4 a叶片Pn、Tr显著高于5 a叶片(P<0.05);4 a和5 a的叶片Ci差异不显著,但显著高于3 a叶片(P<0.05);各株龄的叶片间Gs和WUE均无显著差异(P<0.05)。

表5 不同株龄对老芒麦叶片的光合参数
3 讨论
3.1 各株龄老芒麦的叶片光合基础环境差异
叶片是植物进行光合作用的主要器官[19],叶片的空间分布、生理特征等对植物的生长发育和产量均有影响,尤其在植物生长的后期,旗叶对老芒麦种子产量和质量起到关键作用[20]。前人研究表明,日本落叶松针叶的非结构碳水化合物和叶绿素a/b随冠层的升高而增加,表明针叶对强光的利用效率随冠层的下降而减小[21]。自然光下,下位叶不仅受到冠层的遮蔽而且受到叶片老化的影响,使上位叶的光合贡献率显著高于下位叶[22]。本研究对象为高寒区同一生境下,同一品种的各株龄的老芒麦旗叶,既排除了外部光合环境的差异,亦排除了同一个体的叶位和叶龄间的差异。本研究发现株高大小为3 a>4 a>5 a,且各株龄株高间均差异显著(P<0.05),说明在同一生育期内虽同为旗叶,但随着老芒麦株龄的增加,旗叶获得的空间位置发生了变化。另有研究表明,在老芒麦灌浆期,叶长是影响的光合贡献率的关键因子[17],在大豆的开花期和结荚期,保持植株较大的叶面积,是延缓衰老,且提高后期干物质积累和高产的基础[23]。在老芒麦灌浆期,4 a老芒麦旗叶的叶长和叶面积均达到最大,且与3 a或5 a差异显著(P<0.05),3 a和5 a间无显著差异,说明4 a老芒麦旗叶有较大叶面积,有利于其进一步捕获光能和进行光合作用。灌浆期不同株龄老芒麦旗叶间SPAD值无显著差异,这和李映雪等[24]对小麦叶片和氮素营养的研究发现,开花后各施氮处理下,旗叶作为功能叶,衰老缓慢,其SPAD值高于其余位叶的结果具有相似性。
3.2 各株龄老芒麦的叶片光合特性随光强变化关系及各因子的相关性
光辐射作为重要的环境因子,通过光敏色素等途径调节植物的生长发育,是植物进行光合作用的能量来源[25-26],光合特征参数可反映植物光能利用效率。本研究发现,光子量照度为0~200 μmol/(m2·s)时,各株龄老芒麦叶片Pn和Tr均为线性增长,且呈极显著(P<0.01)正相关关系,这与祁娟等[27]对披碱草属植物进行模拟光合辐射的研究发现,老芒麦在光子量照度<200 μmol/(m2·s)时,Pn随光照呈线性增加,且Pn与Tr呈显著(P<0.05)正相关的结果相一致。本研究结果表明,200~2 000 μmol/(m2·s)光子量照度范围内,各株龄老芒麦叶片Pn,由高到低为3 a>4 a>5 a,说明随生长年限的延长,老芒麦旗叶的Pn呈现降低趋势,这与叶子飘等[28]研究设施栽培条件下三叶青时发现,三叶青一年生的最大净光合速率显著(P<0.05)高于二年生的结果相似。灌浆期内对于旗叶这一特定功能叶片,老芒麦株龄越小其Pn和Tr越高,有利于光合作用的进行,亦利于后期植物种子和产量的积累。
本研究中,各株龄叶片Pn与Gs或Ls呈极显著正相关关系,Pn与Ci均呈极显著负相关,说明老芒麦旗叶的光合作用与气孔导度、胞间CO2和气孔限制有紧密联系。已有研究发现,光合作用的降低与气孔导度、胞间CO2和气孔限制等密切相关[29-30]。当前普遍认为,植物的光合作用降低的主要因子分为气孔限制和非气孔限制[31],由于气孔导度为主要影响,导致胞间CO2浓度,不能满足光合需求,进一步造成光合作用降低称为气孔限制,非气孔限制指叶绿体与核酮糖-1,5-二磷酸羟化酶(Rubisco)等活性降低等引起光合能力下降[32]。本研究中,各株龄老芒麦均出现光抑制现象时,3 a的叶片在1 200 μmol/(m2·s)光子量照度,4 a的叶片在1 000 μmol/(m2·s),表现为Pn降低,Ci降低而Ls增高;5 a的叶片在800 μmol/(m2·s)光子量照度,表现为Pn降低,Ci增高而Ls降低,且各株龄间Pn与Ci均呈极显著负相关,Pn与Ls呈极显著正相关,表明灌浆期老芒麦旗叶出现光合抑制现象时,3 a和4 a气孔限制是主要因素,5 a主要因素是非气孔限制,可能是因为,在同一生境条件下,随生长年限的延长,老芒麦生长周期呈现缩短趋势,其旗叶出现早衰,即出现光合抑制时,5 a株龄叶片的主要因素为非气孔限制,这也部分印证了前人的研究成果[33-34]。
3.3 老芒麦的叶片对光照强度和株龄响应差异
高寒区同一生境下不同老芒麦的旗叶由于老芒麦的株高、旗叶、叶长、叶宽及叶面积等不同而导致其对光照强度和株龄响应能力亦有差异。本研究中,光照强度因素极显著影响老芒麦叶片Pn和Gs(P<0.01),株龄极显著影响叶片Ci(P<0.01),二因素共同作用时,显著影响老芒麦叶片Pn和Gs(P<0.05)。仅光照强度为因子时,各株龄老芒麦叶片均能利用50 μmol/(m2·s)光子量照度的光进行光合作用,且在1 000~1 800 μmol/(m2·s)光子量照度下,Pn均无显著差异,可能是因为研究对象均为老芒麦旗叶且处于相同生育期,即排除了在叶位和叶龄的差异,所以呈现无显著差异的结果,这也和张小晶等[35]研究虉草仅光强为因子时发现,不同位叶在800~1 700 μmol/(m2·s)光子量照度内,Pn无显著差异的结果类似。本研究中,仅株龄为因子时,各相邻株龄间老芒麦叶片的Pn和Tr差异不显著,且3 a和4 a的叶片Pn、Tr显著高于5 a,说明在高寒区老芒麦旗叶在灌浆期时,随株龄升高,其相应的净光合速率和蒸腾速率降低,即光合能力减弱,不利于老芒麦种子形成和产量的积累。
4 结论
灌浆期高寒区不同株龄老芒麦旗叶光合环境基础指标和光合参数均存在差异。随株龄增加其旗叶获光照的空间优势呈递减趋势;200~2000 μmol/(m2·s)光子量照度内,旗叶净光合速率和蒸腾速率呈极显著正相关关系(P<0.01),且均随株龄增加呈递减趋势;各株龄老芒麦旗叶均存在光抑制现象,3和4株龄以气孔限制因素为主,而5株龄主要为非气孔限制因素。
