小麦 TaP5CS基因抗旱相关分子标记的开发
2021-09-23常鹏举杨智全陈东升张晓科
于 铭,魏 凡,常鹏举,杨智全,武 军,陈东升,张晓科
(1.西北农林科技大学农学院,陕西杨凌 712100; 2.宁夏农林科学院农作物研究所,宁夏银川 750002)
小麦是我国最重要的粮食作物之一,其生产关系到国家粮食安全和社会稳定[1]。干旱是降低小麦品质与产量的重要因素,提升小麦的抗旱性是小麦育种迫切需要解决的问题,而抗旱相关基因分子标记的开发可以提高选育抗旱小麦品种的效率。
植物体内积累各种有机或无机物质,其中有机物质包括脯氨酸、甜菜碱等,能够提高细胞液浓度,降低渗透势,提高细胞吸水或保水能力,称为渗透调节作用[2]。脯氨酸的积累有助于增加渗透胁迫的耐受性[3],是高等植物应对干旱胁迫的适应性机制。研究发现,高等植物中脯氨酸合成的途径有两条,一条是从谷氨酸开始合成,另一条是从鸟氨酸或精氨酸开始合成[4];干旱等逆境胁迫下,谷氨酸途径是植物合成脯氨酸的主要途径[5],在吡咯啉-5-羧酸合成酶P5CS的作用下,谷氨酸转化为吡咯啉-5-羧酸,再经过吡咯啉-5-羧酸还原酶的反应最终生成L-脯氨酸。脯氨酸代谢过程中有很多酶的参与,其中 P5CS是脯氨酸合成的限速酶,对脯氨酸合成的速度起着决定性的作用。P5CS基因最早在豇豆中被克隆[3],干旱胁迫下转基因烟草中脯氨酸含量提高了数十倍,增强了烟草的抗旱性[5]。水稻OsP5CS1、OsP5CS2基因及菜豆PvP5CS1和PvP5CS2基因在各种非生物诱导后其表达量明显上调[6]。过表达P5CS基因使脯氨酸含量增加,提高了转基因马铃薯、水稻和小麦的胁迫耐受性[7]。小麦TaP5CS基因转入到拟南芥,促进了脯氨酸的合成积累,说明TaP5CS基因有增强抗旱性的积极作用[8]。在干旱等逆境胁迫条件下,抗旱性较强的品种,其TaP5CS基因的表达量和脯氨酸积累量高于弱抗旱性品种[9],推测两个小麦品种间TaP5CS基因的转录调控序列可能存在差异,可望开发TaP5CS基因抗旱相关的分子标记,为小麦分子标记辅助育种提供帮助。
本研究选用国内不同生态地区的小麦品种(系),在萌发期鉴定抗旱性,并分析干旱胁迫下叶片脯氨酸累积量与抗旱性关系,以及TaP5CS基因响应干旱的表达模式,通过两组极端抗旱品种间TaP5CS基因序列比对和自然群体验证,开发该基因抗旱相关分子标记,以期为抗旱小麦品种的选育与鉴定提供依据。
1 材料与方法
1.1 试验材料
试验材料为全国不同地区的119个小麦品种(系),其中,强抗旱性品种长6878、铜麦4号、沧麦6001、龙麦32、中梁24,中等抗旱性品种晋麦47,弱抗旱性品种西农2208、郑麦9023、云麦46,极弱抗旱性品种扬麦158,用于研究干旱胁迫下叶片脯氨酸积累量与其抗旱性关系;选取2个强抗旱性品种(泰山1号和碧蚂4号)与2个弱抗旱性品种(中麦9号和兰天23),用于TaP5CS基因组序列差异分析;选取一套中国春缺体-四体系材料(N1A-T1B、N1B-T1D、N1D-T1A),用于PCR扩增引物的染色体定位。
1.2 小麦品种(系)萌发期抗旱性的鉴定
按照GB/T 21127-2007“小麦抗旱性鉴定评价技术规范”[10],测定119个小麦品种(系)的相对发芽率,鉴定其萌发期抗旱性。抗旱性分为五级,即:极强(highly resistant,HR)、强(resistant,R)、中等(moderately resistant,MR)、弱(susceptible,S)、极弱(highly susceptible,HS)。
1.3 干旱胁迫下小麦幼苗叶片中脯氨酸含量的测定
将强抗旱性品种(长6878、铜麦4号、沧麦6001、龙麦32、中梁24)、中等抗旱性品种(晋麦47)、弱抗旱性品种(西农2208、郑麦9023、云麦46)和极弱抗旱性品种(扬麦158)每个品种分为两组,室温下用蒸馏水培养3~5 d,挑选生长一致的发芽种子,用1/2霍格兰氏营养液培养至两叶一心期[11]时,一组继续正常培养,作为对照;另一组用含-0.5 MPa PEG-6000的1/2霍格兰氏营养液进行培养,作为胁迫处理,每个处理3个重复,采用茚三酮比色法,测定胁迫120 h时小麦幼苗叶片中脯氨酸的含量[12]。
1.4 生物信息学分析
从Ensemble Plants(http://plants.ensembl.org/index.html)获取小麦基因组序列和基因组注释信息,绘制TaP5CS基因结构图。从Wheat Exp(https://wheat.pw.usda.gov/WheatExp/)获取干旱胁迫下小麦叶片中TaP5CS基因的表达量。
1.5 TaP5CS基因抗旱相关分子标记的开发
依据小麦TaP5CS基因组全长序列,设计嵌式引物,克隆3个同源基因组的全长序列。以萌发期强抗旱性小麦品种(泰山1号和碧蚂4号)与弱抗旱性小麦品种(中麦9号和兰天23)的基因组为DNA模板,分段扩增TaP5CS基因组全长。用DAMAN软件进行序列比对,针对两组极端抗旱性品种间的序列差异设计特异性引物(表1),扩增涵盖变异位点的序列,以此变异位点,分为TaP5CS-1A-a和TaP5CS-1A-b两种等位变异类型,TaP5CS-1A-b含有PflmⅠ限制性内切酶的作用位点,而TaP5CS-1A-a没有PflmⅠ的作用位点(图1)。
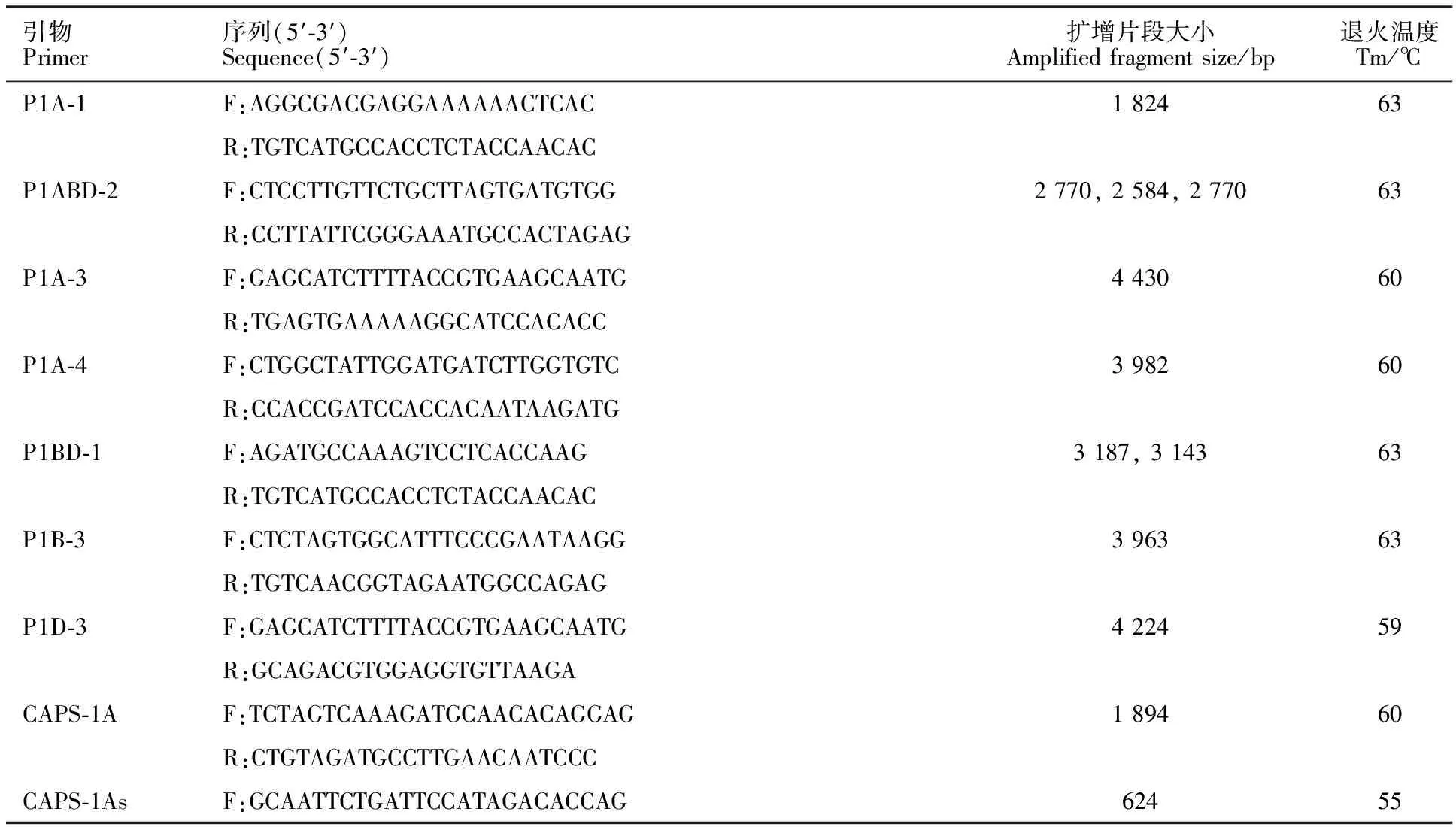
表1 本研究使用的引物信息Table 1 Primer information used in this study
以119个品种(系)的基因组DNA为模板,采用引物CAPS-1A-F和CAPS-1A-R进行扩增,扩增产物使用PflmⅠ进行酶切,酶切产物经 1.2%琼脂糖凝胶电泳,显影照相并统计基因组成;为了电泳条带更加清晰,采用半巢式PCR方法,以上述第一次PCR产物为模板,用引物CAPS-1As-F和CAPS-1A-R扩增涵盖变异位点的小片段序列,扩增产物进行PflmⅠ酶切,电泳检测其基因组成。用SPSS进行不同小麦品种(系)等位变异类型与萌发期相对发芽率的差异 分析。
2 结果与分析
2.1 小麦品种(系)萌发期抗旱性鉴定结果
按照“小麦抗旱性鉴定评价技术规范(GB/T21127-2007)”,鉴定119个小麦品种(系)萌发期的抗旱性。结果发现,供试小麦品种(系)的平均相对发芽率为52.9%,部分品种(系)间相对发芽率的差异达到极显著水平;极强、强、中等、弱和极弱抗旱等级的品种所占比例分别为 2.5%、14.3%、42.9%、33.6%和6.7%。所选材料抗旱性分布范围广泛,适合进行分子标记开发研究。

表2 10个不同抗旱等级的小麦品种模拟干旱胁迫120 h后叶片中脯氨酸含量、萌发期抗旱性等级和相对发芽率Table 2 Proline content,drought resistance level and relative germination rate after simulated drought stress for 120 h of 10 wheat varieties with different drought resistance
2.2 干旱胁迫下叶片中脯氨酸积累量与抗旱性的相关性
从表2可以看出,与对照处理相比,10个小麦品种经120 h的干旱胁迫后,叶片中脯氨酸含量均极显著增加,说明干旱胁迫会使叶片中脯氨酸含量明显积累。干旱胁迫120 h后,不同抗旱性品种叶片中脯氨酸的积累量存在明显差异,其中5个强抗旱性品种叶片中脯氨酸含量达到700 μg·g-1以上,中等抗旱性品种晋麦47脯氨酸含量为623.67 μg·g-1,弱抗旱性品种和极弱抗旱性品种脯氨酸含量低于400 μg·g-1;同时,10个品种干旱胁迫120 h后叶片中脯氨酸积累量与相对发芽率呈极显著正相关(R2= 0.98)(P< 0.01),表明小麦叶片响应干旱的脯氨酸积累量与抗旱性密切相关。
2.3 干旱胁迫对 TaP5CS基因表达模式的影响
从表3可以看出,随干旱胁迫时间的延长,TaP5CS3个同源基因的表达量均呈上升趋势,干旱胁迫1 h时,TaP5CS-1B的表达量迅速增加,与干旱胁迫0 h相比,增加189.88%;干旱胁迫6 h时,TaP5CS-1A的表达量剧增,与干旱胁迫1 h相比,增加1 254.70%,而TaP5CS-1B和TaP5CS-1D的表达量分别增加了605.65%和 1 026.80%。TaP5CS-1A表达量的剧增,说明TaP5CS-1A对小麦长时间干旱胁迫产生了强烈反应。
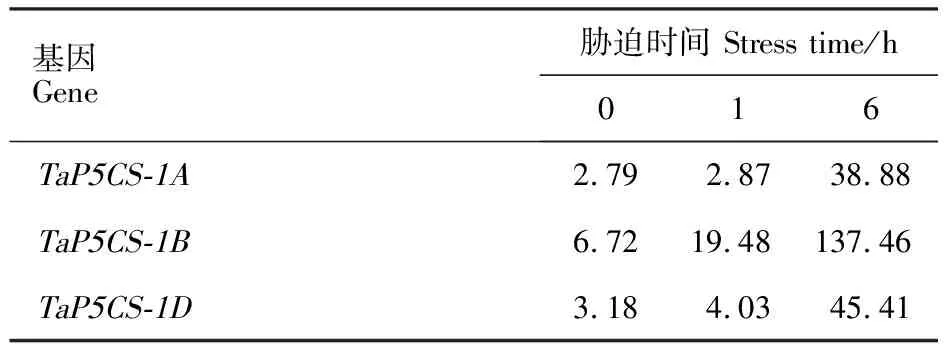
表3 不同干旱时间下 TaP5CS 基因的表达量(FPKM)Table 3 Expression level of TaP5CS genes under different drought time
2.4 TaP5CS基因抗旱相关分子标记的开发与验证
对TaP5CS基因进行分段克隆,分别获得预期大小的目的片段;将4个小麦品种泰山1号、碧蚂4号、中麦9号和兰天23的PCR扩增产物分别测序并拼接后获得各自的基因组全长序列,TaP5CS-1A大小分别为12 996、12 997、12 996和12 995 bp;TaP5CS-1B大小分别为9 559、 9 558、9 559和9 558 bp;TaP5CS-1D大小均为 9 821 bp。
对两组抗旱性极端小麦品种的TaP5CS基因组全长序列进行比对发现,TaP5CS-1B和TaP5CS-1D不存在相对应变异序列;TaP5CS-1A在两组品种间存在2个SNP位点,发生在基因第5、7内含子区域。由于TaP5CS-1A第5内含子565 bp处的碱基G突变成A时,可以被PflmⅠ酶切(图1),用引物CAPS-1A-F和CAPS-1A-R扩增涵盖此SNP位点序列,用中国春缺体-四体系引物进行染色体定位(图2),结果在N1A-T1B和水中未扩增出条带,说明该PCR引物定位在1A染色体上,并且条带大小与预期一致。PCR扩增的产物使用PflmⅠ进行酶切,酶切产物经琼脂糖凝胶电泳检测,发现在强抗旱性品种中酶切产物为单一条带,大小为1 894 bp(图3A),暂被命名为TaP5CS-1A-a等位变异类型;而在弱抗旱性品种中酶切产物为1 706 bp、188 bp两条带(1 894 bp的条带酶切不完全),被命名为TaP5CS-1A-b等位变异类型。
利用PCR扩增及酶切的方法,对119个小麦品种(系)进行TaP5CS-1A基因变异位点检测,分析不同等位变异类型与萌发期相对发芽率之间的关系。在119个小麦品种(系)中,晋麦 79等61个品种(系)在酶切产物中只存在1 894 bp的特异条带,为TaP5CS-1A-a等位变异;淮麦18等58个品种(系)在酶切产物中存在2个条带,分别为1 706 bp和188 bp,为TaP5CS-1A-b等位变异。为了便于观察电泳结果,又设计了一对引物CAPS-1As-F和CAPS-1A-R,采用半巢式PCR的方法,以上述第一次PCR产物为DNA模板,对第二次PCR扩增产物用PflmⅠ酶切,结果与第一次PCR产物酶切完全一致(图3B)。TaP5CS-1A-a和TaP5CS-1A-b等位变异类型品种(系)的平均相对发芽率分别为60.7%和 44.6%,前者的平均相对发芽率极显著高于后者(表4)。抗旱性极强的3个品种(系)的等位变异类型均为TaP5CS-1A-a,抗旱性极弱的8个品种(系)的等位变异类型均为TaP5CS-1A-b;在17个强抗旱性品种(系)中,15个是TaP5CS-1A-a等位变异类型,在40个弱抗旱性品种(系)中,有30个是TaP5CS-1A-b等位变异类型;在51个中等抗旱性品种中,TaP5CS-1A-a等位变异类型占 64.7%,TaP5CS-1A-b等位变异类型占35.3%。由此可见,TaP5CS-1A-CAPS分子标记与小麦萌发期抗旱性存在紧密的相关性,开发的分子标记可进行小麦种质资源抗旱性的鉴定筛选。

表4 TaP5CS-1A-a和 TaP5CS-1A-b等位变异类型的相对发芽率及抗旱性不同级别的品种数Table 4 Relative germination rate and number of wheat varieties with different drought resistance levels in TaP5CS-1A-a and TaP5CS-1A-ballelic variation groups
3 讨 论
脯氨酸是植物在干旱等逆境胁迫下渗透调节的重要物质之一,小麦体内脯氨酸大量积累可保护亚细胞结构和蛋白质的完整性,为小麦正常生长提供保障[3]。抗旱性不同的品种在干旱胁迫后,TaP5CS基因表达量存在极显著差异,且TaP5CS基因表达量增加,脯氨酸含量升高[9],说明小麦品种(系)抗旱性与TaP5CS基因的表达量呈正相关。在干旱胁迫6 h时,TaP5CS-1A基因的表达量与胁迫1 h相比增加了12.5倍,推测TaP5CS-1A基因在胁迫下发挥关键作用。TaP5CS-1A在基因结构上与TaP5CS-1B、TaP5CS-1D存在明显差异,TaP5CS-1A基因含有较多内含子区域,研究表明,内含子可介导基因的表达[13]。胡萝卜素生物合成途径中的关键酶八氢番茄红素合酶(phytoene synthase,psy)基因的内含子变异影响其表达,使小麦籽粒中的黄色素含量发生变化,由此开发了检测不同等位变异类型的分子标记[14]。由此,推测内含子调控TaP5CS-1A基因的表达,使脯氨酸含量升高来响应干旱胁迫。
小麦干旱胁迫后脯氨酸积累量与其相对发芽率呈正相关,高发芽率是小麦高产稳产的前提。本研究在两组抗旱性极端小麦品种间扩增出TaP5CS基因组全长序列,与数据库中提交的序列相似度较高。本研究将TaP5CS-1A基因在编码区6 742~6 983 bp的序列测出,为后续的研究提供基础。经比对2个强抗旱性和2个弱抗旱性品种间的TaP5CS-1A基因组全长序列,发现存在2个SNP位点,在第5内含子内的SNP位点两端设计扩增此SNP位点的特异性引物,由于引物3’端变异导致PCR扩增效率降低,甚至PCR过程启动失败,选择引物3’末端碱基与TaP5CS-1B基因存在差异,且引物3’末端碱基为AG,不容易出现二聚体和发夹结构[15],因此,上游引物和下游引物均仅存一个碱基变异就实现特异性扩增。但是引物CAPS-1A-F和CAPS-1A-R扩增的PCR产物进行酶切检测时,需要使用较大用量的限制性内切酶,酶切后的两条带1 894 bp和 1 706 bp分离需要较高浓度琼脂糖凝胶和较长电泳时间,酶切后188 bp的条带不利于观察,给检测带来诸多不便。因此,采用半巢式PCR方法,以第一次PCR产物为模板,用引物CAPS-1As-F和CAPS-1A-R扩增涵盖变异位点的小片段序列,此时PCR产物酶切条带均较小,使得未被酶切条带624 bp与酶切条带436 bp和188 bp容易分辨。
小麦TaP5CS-1A基因SNP位点存在2种等位变异TaP5CS-1A-a和TaP5CS-1A-b。经119个萌发期抗旱性不同的小麦品种(系)验证, 90.0%的萌发期抗旱性强及以上水平的品种为TaP5CS-1A-a等位变异类型,79.2%的抗旱性弱及以下水平的品种为TaP5CS-1A-b等位变异类型。又因为两种等位变异类型品种(系)的萌发期平均相对发芽率存在极显著差异(P<0.01),说明TaP5CS-1A基因与小麦抗旱性相关,同时也进一步证明了内含子序列的变异与萌发期抗旱表型相关。TaP5CS-1A-CAPS分子标记可为小麦品种抗旱性的检测和鉴定提供辅助方法,但内含子对TaP5CS-1A基因响应干旱的表达调控机理还需要进一步研究。由于脯氨酸代谢过程中有很多酶参与,加上响应干旱胁迫由多基因调控,所以在强抗旱性品种中存在2个TaP5CS-1A-b等位变异类型,在弱抗旱性品种中存在10个TaP5CS-1A-a等位变异类型。总之,小麦抗旱性机理十分复杂,单一基因功能标记不足以全面鉴定其抗旱性,开发多个抗旱相关基因的功能标记来综合全面提高鉴定小麦抗旱性的效率,可加速抗旱小麦品种的选育工作。
