气候因子与大气CO2浓度变化对德令哈祁连圆柏生长与水分利用效率的贡献
2021-09-22杨桃宋苗王晓锋
杨桃,宋苗,王晓锋
(1.中国科学院西北生态环境资源研究院,沙漠与沙漠化重点实验室,甘肃 兰州 730000;2.中国科学院大学,北京 100049)
自18世纪后期的工业革命开始,随着现代工业的发展和人类活动的逐步加剧,温室气体如二氧化碳、甲烷等在大气中的浓度持续增加,使得全球变暖现象越发明显[1-2].IPCC(intergovernmental panel on climate change)第五次报告指出,21世纪气候的变暖是毋庸置疑的,自工业革命以来,全球绝大部分地区都经历了地表增暖的过程.1951年至2012 年,全球地表平均气温升高速率为 0.12 ℃/10a[1],有研究表明,1750年以来,由于人类活动的影响,排放进大气中的温室气体浓度(CO2,CH4,N2O等)已经达到过去80万年来的最高水平,其中CO2浓度增加了41%[2].对于降水而言,全球降水资料的准确性相对温度的准确性降低,但依然指出,1901年来,北半球中纬度陆地区域平均降水量有所增加,但增加幅度存在明显区域差异[3].多年来,气候变化已对森林生态系统产生了一定的影响.有研究表明,由升温导致的干旱胁迫加剧是引起树木死亡的重要原因[4-5],此外,气候变化也可影响林线的变动[6-7]和植被物候的变化[8-9].
森林是陆地生态系统碳循环过程的重要组成部分,存储着整个陆地生态系统45%的碳[10].CO2作为绿色植物光合作用的底物,可以被植被吸收并最终将其转化为有机物,从而参与到生态系统的碳循环中.近些年,关于森林生态系统是否得益于急剧升高的二氧化碳浓度的问题已成为焦点,其中基于控制实验的研究可在叶片和冠层尺度上研究CO2肥化效应的机理[11-12],然而控制实验的样本数量一般有限,且监测时限较短,仅有几年或者十几年,无法在更长时间尺度上探讨植被生长对CO2的响应情况.因此,有必要寻求新的方法来解决上述问题.
树轮代用资料对于重建过去温度、降水、生物量变化等非常有效[13-15].近些年利用树轮宽度和稳定碳同位素年表揭示树木径向生长对CO2浓度升高和气候变化的响应研究可弥补控制实验时间序列长度上的不足.Van der Sleen等[16]利用热带森林的树轮稳定碳同位素年表发现该地区过去150 a间,由于大气二氧化碳浓度的上升,树木长期水分利用效率增长了30%~35%,但树木径向生长并未表现出加速生长的趋势.然而,有研究发现,在青藏高原东部的四川省境内,20世纪之后树木的加速生长及森林的扩张主要是在CO2浓度升高和气候变暖的协同作用下产生的[17].因此,为了更全面地理解树木生长与气候变化和大气CO2浓度升高之间的关系,在不同区域开展此类研究工作有重要意义.
我国西北干旱地区,由于地处内陆腹地,降水量偏少,导致植被多样性简单,生态系统较为脆弱,祁连圆柏作为该地区最重要的建群种,对防风固沙、涵养水源和维持森林生态系统的稳定至关重要,同时也对气候变化和人类活动的响应较为敏感[14,18].然而,前期研究很少考虑到树木径向生长对大气CO2浓度的响应.基于此,本研究以青海德令哈的祁连圆柏为研究对象,建立了树轮断面积指数序列(basal area index,BAI)和稳定碳同位素年表,计算了该地区过去百年的iWUE变化历史,定量分析了气候因子和大气CO2浓度对水分利用效率和树木径向生长的贡献,揭示了祁连圆柏生长的限制因子,在一定程度上可为该地区未来人工林的因地选育和森林经营管理提供科学依据.
1 材料与方法
1.1 研究区概况
本研究区位于柴达木盆地东北缘德令哈地区的宗务隆山(N 37°27′,E 97°47′)(图1),属于祁连山脉中段支脉,山地、高山和山间盆地相间分布;日均温低,日较差大;土壤贫瘠,腐殖质含量低;山地垂直带谱简单,森林呈狭窄的条带状分布.德令哈气象站的监测数据显示,1956~2014年间,该地区年平均气温为3.8 ℃,年平均降水量为170 mm,主要集中在5~9月;月平均最高温出现在7月,为16.75 ℃,月平均最低气温出现在1月,为-11.4 ℃.
1.2 树轮样本采集
本研究采样树种选择该地区的优势种祁连圆柏(JuniperusprzewalskiiKom),采样地点海拔3 750~4 000 m,坡度20°~50°.采样时选取生长健康、树龄较大的样树,使用口径为5.15 mm的生长锥沿与树高1.2 m左右平行和垂直山坡的两个方向分别钻取两根树芯,并进行封存、标记、编号.在宗务隆山玛尼图地区共计采集样树208棵,之后将样芯带回实验室,按照树轮样本处理的流程先进行晾干,随后将样芯固定在样品槽上,用砂纸进行打磨至年轮界限清晰可见.然后利用LINTAB 6树轮宽度测量仪对每个样芯进行宽度测量(精度0.01 mm),并利用TSAPwin软件进行交叉定年,最后用COFECHA程序对定年结果进行检查,确保定年结果准确无误.
1.3 断面积指数计算
当树轮宽度保持稳定或呈现变窄的趋势时,木材体积可能持续增加,说明树轮宽度可能并未完全消除由于树木老龄化导致的生长下降趋势[19].为了更好地消除这种下降趋势,同时保留气候因素对树木生长的影响信号,本研究将树轮宽度转化为断面积指数进行分析.为了尽量减少误差,我们选取了63棵带有髓心的树轮样本宽度计算BAI,先计算每个样芯的BAI值,随后平均为区域BAI序列(图2).基于本研究的目的和树轮稳定碳同位素序列的长度,选取的研究时段为1890~2010年.断面积指数计算公式[20]:
(1)
式中:Rn和Rn-1分别代表在第n年和n-1年到髓心的距离.
1.4 树轮稳定碳同位素年表建立
在所有样芯准确定年的基础上,选取6棵(表 1)轮宽清晰、无断裂、无污染的样本用于树轮稳定同位素测量,详细操作流程按照Wieloch等[21]文献中所述进行.首先对样品进行逐年剥离,利用特质单面刀片或手术刀剥离并切碎木材,剥离时需在LINTAB测试平台的物镜显微镜下进行;其次则为α-纤维素提取,该过程较为复杂,主要是利用化学试剂,如NaOH、NaClO2、盐酸等逐步对木材样本进行处理,从而得到α-纤维素;再次,用超声波仪对提取的α-纤维素进行进一步的均质化处理,然后在冷冻干燥机中冻干;最后进行同位素测定,测定时采用ONLINE分析测试系统,即高温转换元素分析仪和Finnigan MAT-253气体稳定同位素质谱仪测定13C/12C比率,分析误差小于0.15‰.德令哈树轮稳定碳同位素年表时段为1890~2010年,1951年之前为3 a分辨率,之后为年分辨率.
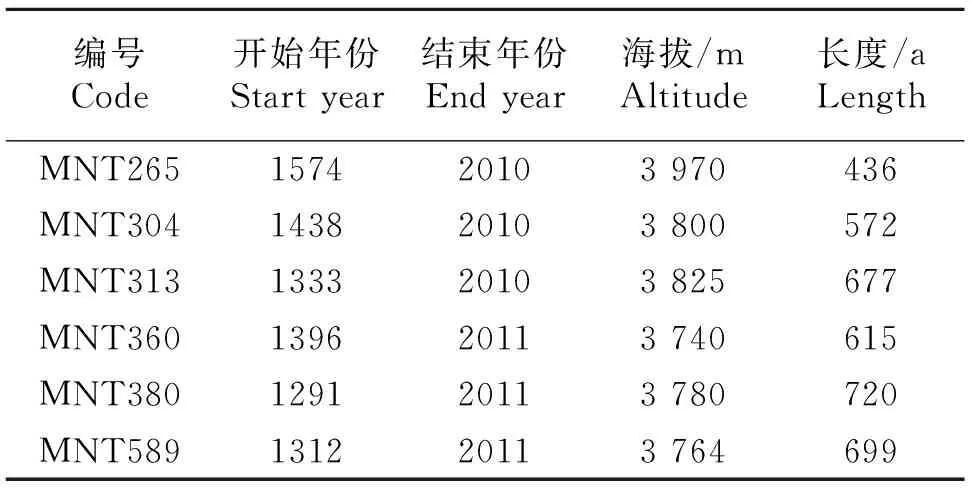
表1 德令哈地区树轮稳定碳同位素样本信息
1.5 气象资料收集
本研究所用的气象资料来自于国家气象信息中心(http://data.cma.cn/site/index.html),选取距离采样点最近的德令哈气象站1956~2010年的月值气候数据资料,包括平均温度、降水量、平均相对湿度,并利用得到的温度和相对湿度数据计算饱和水汽压差(VPD)[22].此外,选用标准化降水蒸散指数(SPEI,standardized precipitation-evapotranspiration index)格点数据(SPEIbase v.2.4 [Dataset],2016,http://hdl.handle.net/10261/128892)来代表本研究区干旱状况.
1.6 水分利用效率计算
植物在光合作用过程中,优先会摄入较轻的12C,较重的13C则会产生分馏,结果导致植物光合产物中的13C/12C比率低于大气中的比率.Farquhar等[23]研究发现C3植物叶片中稳定碳同位素分馏值(Δ)可通过下面的公式计算:
(2)
同时,C3植物体内碳同位素分馏还可以用下面的公式表示:
Δ13C=a+(b-a)Ci/Ca
(3)
式中:δ13Ca和δ13Cp分别代表大气中13CO2和树轮δ13C(图3).a表示扩散分馏系数(4.4‰),b表示羧化分馏系数(27‰),Ci为叶片内部的CO2浓度,Ca为叶片外部CO2浓度,即大气CO2浓度(图3).结合公式(2)和(3),可计算出叶片内外CO2比值(Ci/Ca),最后祁连圆柏长期水分利用效率可通过以下公式得到:
(4)
1.7 消除气候因素对水分利用效率的影响
为了去除气候信号对水分利用效率的影响,首先要将气候信号从叶片内CO2浓度中分离[24],具体做法为先将Ci与气候因子做逐月相关分析,找到相关性最显著的月份之后,对气候因子(温度、降水、VPD)做主成分分析,得到的第一主成分即为气候因子目标(PCT,optimum climatic target).然后利用经验公式Ci+τ×(Ca-280),通过τ取值的改变(τ∈[-1,2],步长0.004),得到若干条模拟的叶片内二氧化碳浓度序列(Ci-climate)[24].当模拟的Ci-climate序列对PCT的方差解释量最高同时二者的残差序列斜率最小时,此时的模拟序列被认为是最优的,即代表由气候因子引起的叶片内二氧化碳浓度变化,二者之差则为由CO2浓度导致的Ci变化,最后通过公式(4)可得到由CO2浓度升高导致的水分利用效率变化部分(cciWUE).
1.8 共性分析
共性分析(commonality analysis)是基于解释变量和响应变量之间的多元线性关系,计算解释变量对响应变量解释方差的模型[25],可由R语言“yhat”包中的commonalityCoefficients函数实现.本研究中计算了温度、降水、SPEI和CO2对在1956~2010年间水分利用效率和BAI的贡献,该贡献既包括各解释变量对响应变量的单独解释量,又包含解释变量对因变量产生的交互效应,即共同解释量.
2 结果与分析
2.1 树木生长与树轮稳定碳同位素年表的变化
从BAI的长期变化来看,德令哈地区的树木径向生长在近100 a有明显上升趋势,尤其是1950年之后上升趋势显著(图2).1890~2010年间,BAI平均值为2.95 cm2/a,变化范围为0.76~6.06 cm2/a.从图3可以看出,树轮δ13C年表在1950年之前相对平稳,之后呈现显著的下降趋势,德令哈祁连圆柏树轮δ13C在1890~2010年平均值为-19.5‰,变化范围为-20.97%~-17.99‰(图3-A).与碳同位素年表相反,大气二氧化碳浓度变化则是1950年之后表现出显著的上升趋势(图3-B).
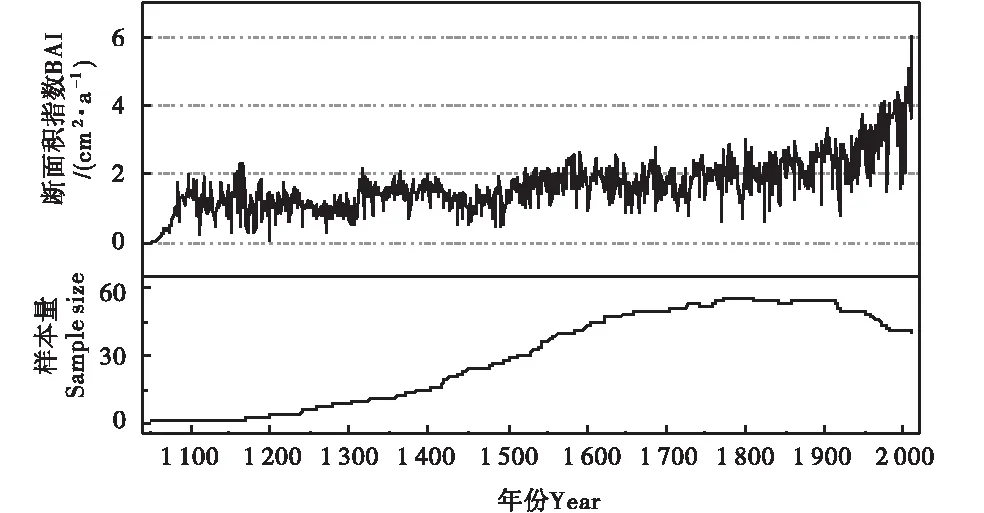
图2 德令哈地区BAI序列及样本量分布Figure 2 The BAI series and sample size of Delingha region

图3 树轮稳定碳同位素年表及大气13CO2(A)和二氧化碳浓度(B)在1890~2010年间的变化Figure 3 Tree-ring δ13C chronology,atmospheric 13CO2 (A) and CO2 (B) during the period 1890~2010
2.2 Ci、iWUE和BAI 对气候因子的响应
气候监测资料表明,1957~2010年间,德令哈地区年平均温度显著上升,上升速率为0.46 ℃/10年,不仅如此,年平均降水量也表现出显著的增加趋势,增长速率为2.24 mm/a,但相对湿度却呈显著下降趋势,下降速率为0.067%/a(图4).
相关分析显示,叶片内CO2浓度(Ci)与温度和饱和水汽压差(VPD)在绝大多数月份上都表现出了正相关关系(图5-A),相关性显著的月份集中在上年10月至当年4月及当年8~10月.相关性最高的月份为上年12月,Ci与温度和VPD的相关系数分别为0.63和0.59和(P<0.05).月份组合之后的相关性发现上年8月至当年5月气候因子与Ci之间的响应是最佳的.Ci与降水在绝大多数月份也表现为正相关关系,且显著性月份主要发生在当年1~3月和生长季,相关性最强的月份为当年7月(r=0.52,P<0.05)(图5-A).iWUE与温度、VPD、降水和SPEI在所有月份都呈正相关关系,与相对湿度负相关,生长季前期的温度和饱和水汽压差对iWUE的响应比较敏感,相关系数均超过0.6(图5-B),降水与iWUE的相关性较弱,当年7月、9月降水与iWUE相关性分别为0.41和0.39,SPEI与iWUE最高的月份发生在当年8月,相关系数为0.49(P<0.05).月份组合后发现上年10月至当年9月气候因子与水分利用效率的相关性是最强的(图5-B).树木径向生长(BAI)与上年10月至当年3月的温度正相关,在生长季期间,BAI对当年5、6月降水(r=0.47,P<0.05)和相对湿度(r=0.4,P<0.05)的响应均呈显著正相关,而与6月的温度显著负相关,6~8月的平均SPEI与树木径向生长的相关性最高,达到0.64(P<0.05,图5-C).
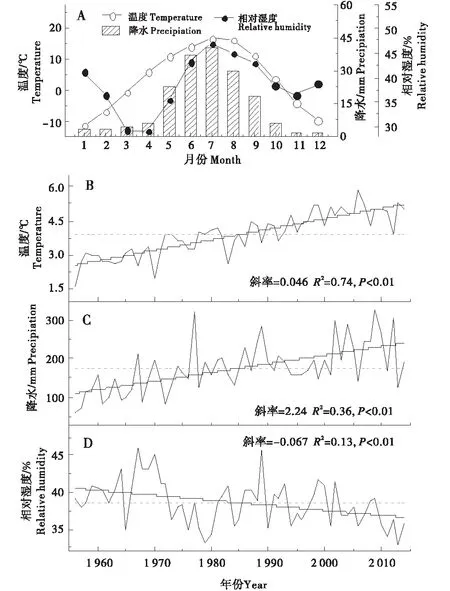
图4 德令哈气象站温度、降水及相对湿度的年内(A)和年际变化(B~D)Figure 4 The intra-annual (A) and annual variations (B-D) of temperature,precipitation and relative humidity of Delingha meteorological station

“*”表示相关性达到95%显著性水平.The coefficients significant at the 95% confidence level are displayed with “*”.图5 1957~2010叶片内CO2浓度(Ci)、水分利用效率(iWUE)和断面积指数(BAI)与各月份及其组合之间的相关分析Figure 5 Correlation analysis between tree-ring estimated Ci ,iWUE,BAI and climate variables and monthly combination from 1957 to 2010
2.3 Ci与iWUE的长期变化特征
1890~2010年间,大气CO2浓度升高了约70 ppmv(图3-B),由树轮稳定碳同位素计算的叶片内CO2浓度在相同时段内也呈现逐渐升高的趋势,最大值超过140 ppmv(图6-A),提高了30 ppmv,增长率为29%(图7-A).由气候因子影响的叶片内二氧化碳浓度变化序列(Ci-climate,τ=0.7时)对Ci的解释方差最高,达到72%(图6-B).德令哈地区iWUE也有明显升高趋势,尤其是1950年之后,在1890~1915年间,iWUE平均值为133.70 μmol/mol,相对最初时段(1890~1915)增加了30 μmol/mol,增长率为30%(图7-B~C).由图7可以看出,CO2浓度升高导致的水分利用效率变化部分也表现出逐步上升趋势,cciWUE的增长率为43%.
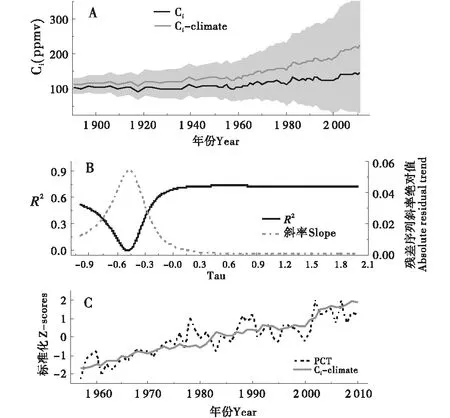
A:树轮δ13C计算的Ci与经验公式计算的叶片内二氧化碳浓度序列(Ci-climate)变化,阴影为τ由[-1,2]为变化区间计算的Ci-climate;B:Ci-climate对气候因子目标(PCT)的方差解释量及其残差序列斜率变化;C:Ci-climate与PCT对比.A:Tree-ring estimated Ci and the intercellular CO2 concentration series (Ci-climate),which was calculated by empirical equation; B:The grey area indicates all calculated Ci-climate time series when changing τ from -1 to 2;C:Ci-climate expalins the variance in instrumental target (PCT) and changes of the absolute residual trend;Comparision between Ci-climate and PCT.图6 由经验公式Ci+τ×(Ca-280)计算的气候因子引起的叶片内二氧化碳浓度变化Figure 6 Changes in intercellular CO2 concentration caused by climate factor calculated by the empirical formula Ci+τ×(Ca-280)

灰色阴影代表iWUE标准差.Grey shading area indicates iWUE standard deviation.图7 1890~2010年水分利用效率变化(A,B)及其与叶片内二氧化碳浓度(C)变化率Figure 7 The variations of iWUE and percent change of iWUE (A,B) and Ci (C) from 1890~2010
2.4 气候因子和CO2对iWUE和BAI的贡献
共性分析模型计算了1956~2010年温度、降水、SPEI和CO2对iWUE的贡献.结果显示,所选因子对 iWUE的总方差解释量达到88.6%,CO2对iWUE的方差解释量占据绝对优势,其单独贡献量为16.0%,气候因子对iWUE的单独贡献量相对较少(图8),表明德令哈地区祁连圆柏水分利用效率在1950年之后的加速上升趋势与大气CO2浓度的急剧升高有密切关系.共性分析结果还发现温度与CO2之间的交互效应对iWUE的共同解释量达到49.7%(图8),说明二者的协同作用对于iWUE的变化也产生了较强的影响.虽然温度的升高在一定程度上与大气CO2浓度的上升有关,但并非是严格相关的关系,这也是共性分析结果发现温度对iWUE的单独解释量不明显的原因.同样地,通过共性分析计算气候因子和CO2对树木径向生长的贡献之后,发现所有影响因子对BAI的总体方差解释量达到51.1%,其中降水量和SPEI对BAI的单独方差贡献量占14.7%,而大气CO2的单独贡献量非常小(图8),另外降水和SPEI之间的交互效应对BAI的共同解释量达到17.1%,说明在近50年,树木生长的年际波动与降水和SPEI的变化关系紧密.

图8 温度、降水、SPEI和CO2分别对iWUE和BAI在1956~2010年间的贡献率Figure 8 The contribution of temperature,precipitation,SPEI and CO2 in explaining the variance of iWUE and BAI for 1956~2010 by using commonality analysis
从图9可以看出,干旱指数与树木径向生长之间的正向关系非常显著,1951~2010年间二者的相关系数为0.61(P<0.05).基于此结果,本研究计算了每年高于和低于平均生长量40%的树木所占比例,发现树木径向生长在长期趋势上与干旱胁迫的协同变化也比较一致,尤其是在极端湿润或者干旱的年份,超过50%的树木则会表现出明显的高生长量或者低生长量(图9-B).
3 讨论
3.1 气候因子对树木生长过程的影响
树木生长的同时可记录影响其生长的环境要素信息[26].由相关分析结果可知,生长季前期的温度与祁连圆柏径向生长显著正相关,说明这个时期温度的升高有利于树木的生长.德令哈地区冬季的平均温度在-10 ℃左右,祁连圆柏处于生长休眠期,来年温度逐渐升高时,开始逐渐恢复其生理活动,此时较高的温度将有利于祁连圆柏生理活动的顺利进行,同时有利于存储在树木体内物质与能量的利用.Yang等[27]在青藏高原东北部利用树轮宽度和树轮生理生态模型的研究发现,温度的升高可导致树木生长季开始时间显著提前.进入生长季之后,我们发现祁连圆柏径向生长与6月份温度显著负相关,而与5、6月份的降水显著正相关.对于祁连圆柏而言,早材形成时期温度的升高导致潜在蒸散发增强,不利于树木生长,而此时较多的降水有利于缓解树木对水分的需求.有研究发现,在我国西北干旱半干旱区,祁连圆柏平均细胞生长速率快速增加时段主要发生在5、6月份,且其生长量主要由平均细胞生长速率控制,在祁连圆柏形成层活动期间,降水起着至关重要的作用[28].另外,我们的研究还发现,生长季期间祁连圆柏径向生长与干旱指数始终保持显著正相关,说明随着温度的逐渐升高,当降水量不足时,干旱状况成为限制祁连圆柏径向生长的主要气候因子,这与前期研究结果是一致的[14,29].

图9 德令哈地区祁连圆柏BAI序列和SPEI对比及树木生长量变化(A)及0值以上柱状图代表高生长量树木比例,以下代表低生长量(B)Figure 9 Comparison between BAI and SPEI for Qilian juniper in Delingha and the changes in tree growth(A),Bars above zero indicate an increase tree growth,while below zero indicate decrease in tree growth(B)
3.2 祁连圆柏水分利用效率变化原因
iWUE是指单位水量通过植物叶片蒸腾耗散时所能同化的光合产物[30].前期研究发现大气CO2浓度和气候因子是影响iWUE变化的两种重要因素[24,31].在外界环境条件适宜的情况下,植被叶片的气孔充分张开,随着大气中CO2浓度的增加,进入叶片内部的CO2浓度也会随之增加,从而提高植被光合速率,进而使得其水分利用效率升高[32].在本研究中,我们发现祁连圆柏叶片内CO2浓度明显升高,且iWUE也增长了约30%,cciWUE的增长率为43%(图7),说明大气二氧化碳浓度的升高对于iWUE的升高有一定促进作用.CO2浓度的升高使得植被iWUE增加的结论已被大多数研究证实[16,33-34].然而,在干旱半干旱区,气孔因素也是限制iWUE变化的另一重要因子,当植被处于轻度或者中度水分胁迫时,为了减少水分丧失,气孔开度减小,使iWUE升高[35].由相关分析结果来看,祁连圆柏iWUE与干旱指数在绝大多数月份上都呈显著相关关系,说明在德令哈地区,干旱胁迫对iWUE具有重要影响.王文志[31]发现祁连圆柏在干旱时期的iWUE高于湿润时期,主要原因是由于干旱时期气孔导度降低,植物蒸腾作用减弱,植物体内参与有机物合成的水分增加,水分利用效率增加.
3.3 祁连圆柏生长与水分利用效率的关系
共性分析结果表明,iWUE主要受到二氧化碳和温度的影响,而BAI则受到降水和干旱状况的控制,二者本身记录的环境影响因子就有差异,因此单纯利用简单的线性相关关系来分析iWUE对树木径向生长的影响是不准确的,这很可能是导致CO2肥化效应研究结果产生区域差异的一个原因.另一方面,光合作用作为树木生长过程中最重要的生理活动之一,大气二氧化碳浓度的上升会使得植被光合速率提高,这一点在许多研究中均有发现[12,36].然而,这部分由于光合速率提高而增加的碳水化合物可能并未被分配进树干中.祁连圆柏作为我国西北干旱半干旱地区特有的耐寒、耐旱的乔木树种,由于该区域降水较少,树木为了能够汲取更多的水分以满足自身生长的需要,则会形成较为发达的根部系统,以便吸收更深层的地下水[37].因此,祁连圆柏光合速率提高产生的碳水化合物可能会更多的用于根部的生长.相反的,如果这种干旱状况得以有效缓解,那么更多的碳水化合物将会被分配到植被的茎干和枝叶中.
4 结论
德令哈地区的祁连圆柏水分利用效率在20世纪有明显增加的趋势,与1890~1915年相比,水分利用效率增加了30 μmol/mol,增长率为30%.通过定量计算发现大气CO2浓度对水分利用效率的单独贡献量最大,为16%,气候因子对其的贡献量均较小.表明大气CO2浓度的加速上升促进了祁连圆柏iWUE的升高.另外,研究还发现,祁连圆柏径向生长也呈现显著升高的趋势,共性分析结果显示BAI主要受到降水和SPEI的影响.且圆柏径向生长在不同时段都与干旱指数呈现正相关关系,尤其是近几十年,随着德令哈地区的逐渐变湿,祁连圆柏径向生长趋势也显著增加,说明干旱状况的缓解有利于促进树木生长.
