大气CO2浓度升高和氮肥互作对玉米花后功能叶碳氮同化物的影响*
2021-09-17李迎春牛晓光何雨桐郭李萍
李 明,李迎春,韩 雪,牛晓光,马 芬,魏 娜,何雨桐,2,郭李萍**
(1.中国农业科学院农业环境与可持续发展研究所/农业农村部农业环境重点实验室,北京 100081;2.沈阳农业大学农学院,沈阳 110161)
全球气候变化已被科学观测所证实,美国国家海洋和大气管理局(NOAA)及地球系统研究实验室(ESRL)的NOAA-ESRL 协同空气采样网络观测结果显示,2019年大气中CO2浓度已达411.49μmol·mol−1。根据IPCC 2001年和2014年报告,不同温室气体排放情景下,到本世纪中期预计大气CO2浓度将达到450~550μmol·mol−1,到本世纪末 CO2浓度将增加到约900μmol·mol-1。CO2作为光合作用的原料之一,其浓度在大气中升高可以通过影响作物的生理代谢活动而影响根、茎、叶等器官的生长发育,并影响作物的产量和品质[1]。碳是干物质的主要成分,氮素是作物第一大营养元素,碳氮代谢对作物产量及籽粒品质的影响最为关键,因此,明确未来大气CO2浓度升高对作物花后碳氮代谢关键组分的影响具有重要意义。
目前国内外关于大气CO2浓度升高(elevated CO2,简称eCO2)对作物生长影响的研究主要借助环境控制试验(Controlled environment,CE)、开顶式气室(Open−top chambers,OTC)和自由大气CO2浓度富集试验(Free−air CO2enrichment,FACE)平台[2]等进行。由于FACE 系统建造及运行成本都较高,多数研究主要采用前两种方式进行,而其反映田间真实大气状况的代表性不足。eCO2对作物生长的影响研究方面,国内外主要对C3作物开展的研究较多,对C4作物的研究相对较少。碳代谢指碳水化合物代谢,包括光合作用各过程及光合碳产物的运输、贮存及分解等过程。碳水化合物是作物光合作用的主要产物,按其存在形式可分为结构性碳水化合物(SC)和非结构性碳水化合物(NSC)两大类,其中NSC 中的可溶性糖是光合碳同化物在作物源库间转运的主要形式,对植株生理代谢过程和产量形成起直接作用[3]。氮素是各种重要酶和功能蛋白的组成成分,在作物的生长代谢过程中都起到直接作用,根据氮素在作物体内的代谢形式可将其分为可溶性含氮化合物(包括硝态氮、氨基酸、可溶性蛋白等)和非溶性氮素化合物(包括类囊体氮、细胞膜氮、细胞壁氮等)两大类[4]。氮代谢指作物体内各种含氮化合物的转化,如由硝态氮还原为铵态氮,以及由铵态氮合成为氨基酸,而后合成蛋白质,以及各种细胞器物质和细胞壁等物质的同化等过程。碳氮代谢是作物生长最基本的生理过程,其在生育期间的动态变化与光合作用各过程及光合产物的形成、转化以及矿质营养的吸收和运输、以及蛋白质的合成等密切相关,并受到自身遗传特性和CO2浓度、光照、水分等环境因素的影响[5]。碳、氮代谢在很大程度上决定农作物的生长,其协调程度不仅影响作物生长发育进程,还是源−库关系协调的基础,最终关系到作物产量和品质[6−8]。eCO2对作物生长的影响还与养分供应及水分状况等环境因素密切相关[9−10]。玉米开花吐丝后开始灌浆,光合产物及储存在营养器官中的养分开始转移到籽粒中并决定穗粒数和粒重[4,11]。养分是影响作物生长的主要因素之一,生产中气候变化是与多种环境因子共同交互存在。前人研究结果表明,在eCO2下,一些作物碳同化能力增强、生物量积累加快,对氮素供给提出了更高的要求[12]。关于eCO2下碳氮代谢及其与氮互作对作物产量的影响报道还鲜少,目前有几种不同的研究结果。OTC 盆栽实验的研究表明[13],氮素胁迫在常规大气CO2(ambient CO2,简称aCO2)和eCO2下均显著降低了谷子叶片比叶质量和叶片氮;eCO2下谷子叶片光合性能较aCO2对氮素胁迫的响应更加敏感,但构成籽粒产量的参数并未显著下降。在小麦上的研究也发现,eCO2可提高冬小麦产量,并与氮肥有明显的正向互作关系,高氮肥处理可降低 eCO2对生育期的加快作用,提高光合能力,促进CO2肥效的发挥[14]。但是,也有研究显示,水稻产量在不同CO2浓度水平下无显著差异,但随施氮水平的提高而增加[15]。还有OTC 实验表明,eCO2显著促进水稻中等氮水平下叶片中非结构性碳水化合物的积累,但抽穗期水稻叶片氮含量却显著降低[16],这也说明eCO2下,由于碳的积累、碳氮代谢不协调,氮的代谢可能受阻。另外也有报道显示,在相同施氮水平下,eCO2对于C4作物玉米产量则不产生显著影响[17]。
当前关于eCO2单独作用及其和氮肥交互作用对作物影响的研究主要集中在C3作物上,并且大多数利用OTC 进行单一因素影响研究,对C4作物的研究较少,而且一些研究的结果也不尽相同。玉米作为重要的C4作物,在全球种植面积最广。气候变化与环境因子共同对作物生长及产量产生影响,在当前全球变化背景下借助理想的FACE 平台研究eCO2与其它因子如氮肥施用的交互作用对玉米生长的实际影响非常必要,无论从影响机理、产量变化及评估模型参数校准等各方面都迫切需要直接的田间试验数据。
因此,本研究利用与真实大气状况最为接近的FACE 平台,研究eCO2和氮肥两因素交互作用对C4作物玉米生殖生长期功能叶不同碳氮代谢指标的影响及动态变化,并同步监测对玉米生物量和产量的影响,以明确在未来以大气CO2浓度升高为代表的气候变化背景下,氮素施用及碳氮协调管理对玉米关键碳氮代谢物质量分数的影响及产量变化情况,以期为全球气候变化下玉米的稳产增产、养分管理及玉米作物评估模型调参提供数据支撑。
1 材料与方法
1.1 试验FACE 平台概况
FACE平台于2007年由中国农业科学院农业环境与可持续发展研究所建立,位于北京昌平(40.13°N,116.14°E)。FACE 试验系统主要包括CO2传感器、CO2气体供应装置和控制系统。FACE 圈由8 根CO2气体释放管组成八边形,圈直径为4m,芬兰产Vaisala CO2传感器放置于圈中心冠层上方,用以检测圈内CO2浓度。CO2浓度通过计算机程序控制,并根据具体风向和风速控制释放管电磁阀的开合度和方向,以实现预定浓度(550μmol·mol-1)供应,该FACE 平台已稳定运行十余年。
1.2 试验点气候及土壤状况
试验点所在地区属暖温带半湿润大陆性季风气候区,土壤类型为潮褐土,2019年夏玉米播前0−20cm 不同小区土壤基础理化性状为有机碳11.2~12.4g·kg−1,碱解氮92.7~129.1mg·kg−1,全氮0.94~1.45g·kg−1,速效钾129.7~156.2mg·kg−1,速效磷31.5~45.0mg·kg−1,pH 8.2。
1.3 试验设计
田间试验设4 个处理、3 次重复,即在aCO2(试验期间为400μmol·mol-1左右)和eCO2(550μmol·mol-1)下分别设置无氮和施氮(分别简称ZN:zero nitrogen 和CN:conventional nitrogen)2 个氮水平,对应的氮肥用量分别为0kg N·hm−2和180kg N·hm−2。基肥和追肥的氮肥用量分别占40%和60%,基肥在播前撒施后旋耕翻入土中,追肥在玉米喇叭口期降雨前撒施。各处理的磷钾肥用量相同,分别为 150kg P2O5·hm−2和 90kg K2O·hm−2;磷肥和钾肥全部作为基肥一次性施入。
FACE 系统共12 个试验小区,aCO2和eCO2各6个小区,小区内FACE 圈直径为4m,各圈外沿相距至少23m。施氮和不施氮处理分别有3 个圈。2019年夏玉米生长季整个生育期FACE 圈内与对照CO2浓度见图1,其中生育期FACE 圈91%的时间内CO2浓度在550±40μmol·mol-1;常规大气CO2浓度监测情况为91%时间内CO2浓度在400±35μmol·mol-1。从夏玉米出苗开始释放CO2气体,一直到玉米成熟时停止供气。通气时间为每日6:00−18:00,夜间不释放CO2。玉米季有2 层CO2气体释放圈,下层圈最高位置在穗位叶处,上层圈高度保持在冠层上方15cm 处。

图1 FACE 装置田间图Fig.1 Free Air CO2 Enrichment(FACE)system in the experimental field
1.4 田间管理
试验地于2019年6月19日撒施基肥后旋耕,夏玉米品种为“郑单958”,6月20日播种,9月28日收获,种植密度66666 株·hm−2。玉米于8月15日抽雄吐丝,9月15日进入乳熟期。期间,6月21日喷灌约3cm 以保证出苗整齐;7月29日雨后土壤湿润,8月1日撒施氮肥追肥;灌浆后期(8月30日)由于土壤墒情差,给予一次渠灌,灌溉量约5cm。气象数据来自试验站自动气象站,夏玉米生长期间每日气温及降水情况见图2。

图2 夏玉米生长季冠层CO2浓度(a)及气温和降水(b)情况Fig.2 Mean daily CO2 concentration(a),daily air temperature and precipitation(b)during summer maize growing season above the plant canopy
1.5 各指标取样及测定方法
1.5.1 生物量
在玉米六叶期(V6)、大喇叭口期(V12)、抽雄吐丝期(VT)、籽粒建成期(R2)、乳熟期(R3)和成熟期(R6)取3 株地上部整株。分别记录叶片、秸秆和穗的鲜重,之后105℃下杀青30min后75℃烘干至恒重。
1.5.2 测产
在玉米籽粒成熟期实收测产,每小区选择1m2内3行12 株玉米,待自然晾干后脱粒称重计算产量。
1.5.3 产量要素考种
从每小区所收获的玉米植株中选择3 株代表本小区内平均长势的玉米穗进行考种,测定指标包括穗长、秃尖长、穗周长、列数、列粒数和每株穗粒数等产量要素。
1.5.4 主要碳氮代谢物取样及测定方法
在VT 期、R2 期、R3 期取功能叶测定主要碳氮代谢物质量分数。于每小区中各选3 株长势代表本小区玉米植株穗位叶,从叶鞘处剪下,其中一部分用锡纸包裹立即置于液氮罐内用于测定主要碳氮同化物质量分数,另一部分杀青后烘干用于干样碳氮质量分数测定。
主要碳氮代谢物中,可溶性糖和淀粉质量分数采用硫酸蒽酮比色法[18]测定;硝态氮质量分数采用水杨酸比色法[19]测定,游离氨基酸质量分数采用水合茚三酮比色法[20]测定,可溶性蛋白质量分数采用考马斯亮蓝G−250 染色法[18]测定,细胞壁氮和类囊体氮质量分数按照文献[21−23]方法测定。
干样粉碎过100 目筛后利用碳氮元素分析仪对玉米功能叶总碳和总氮质量分数进行测定。
1.6 数据处理和分析方法
数据处理和图表绘制采用Excel 2016 软件完成。方差分析和显著性检验采用SPSS 18.0 软件的ANOVA功能进行(Duncan 法);CO2浓度和氮肥双因素交互作用通过SPSS18.0 软件中的一般线性模型进行评估。
2 结果与分析
2.1 大气CO2浓度升高和氮肥施用对夏玉米生物量和产量的影响
由表1 可知,夏玉米各处理地上部生物量随玉米生育期进程(V6、V12、VT、R2、R3 及R6 期)而逐渐增加(图3),R2 期至R3 期增长速率最快。成熟期籽粒产量为8.57~9.15t·hm−2。
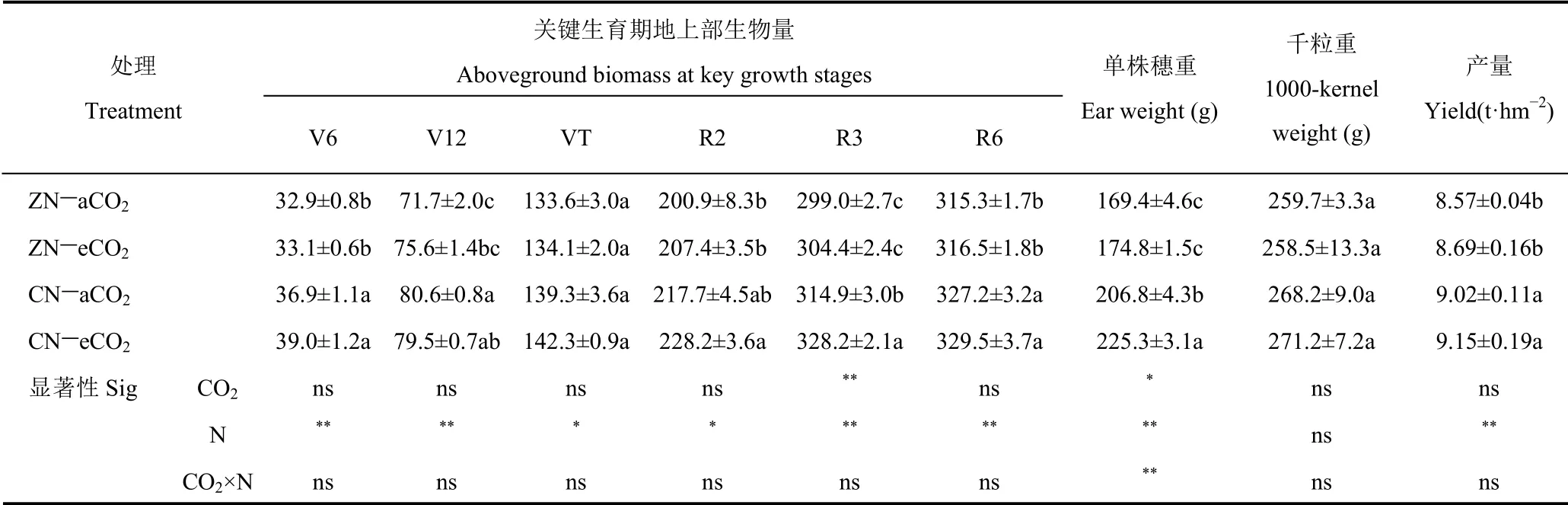
表1 不同CO2 水平下无氮和施氮处理夏玉米各生育期地上部生物量和产量及产量构成因素的比较Table 1 Comparison of aboveground biomass and yield and yield components of summer maize under ZN(zero nitrogen)and CN(conventional nitrogen)at different CO2 levels at key growth stages
不同时段各处理间单因素方差分析表明,单一eCO2因素对夏玉米生物量有一定促进趋势,但仅在R3 期CN 水平下较eCO2处理显著增加4.2%(P<0.05)。eCO2对产量构成因素中的单株穗重在CN 水平下影响显著(P<0.05),比aCO2处理显著增加8.9%(P<0.05)。单一aCO2因素对其余时期各处理生物量和产量其它因素无显著影响。单一氮肥因素下各个时期夏玉米地上部生物量显著增加(P<0.05),其中开花之前平均增加9.6%,花后平均增加6.6%。单一氮肥作用下夏玉米穗重和产量显著增加26.6%和5.3%(P<0.05)。说明两因素交互作用可以显著增加夏玉米单株穗重,而对夏玉米地上部生物量、千粒重和产量无显著影响。
因此,对于C4作物夏玉米而言,氮肥仍然是地上部生物量和产量增加的主要因素;eCO2总体上会促进生物量和产量及其构成因素的增加,但产量总体在统计水平上不显著。eCO2和氮肥的交互作用尽管会显著促进夏玉米单株穗重的增加,但对生物量、千粒重和产量影响不显著。
2.2 CO2浓度升高和氮肥互作对功能叶主要碳同化产物的影响
功能叶中可溶性糖质量分数的高低,可反映植株体内光合碳合成及同化物的源供应水平。光合作用合成的可溶性糖是碳同化物从源到库运输的主要形式,如玉米的碳同化物从源到库的运输形式主要是蔗糖;而合成的淀粉也可暂时在叶绿体内积累,成为叶中最丰富的复杂碳水化合物储存形式;夜间光合作用停止时,叶中储存的淀粉可以降解,为作物的各项必需生理活动提供能量。
由图3 可知,在夏玉米抽雄期(VT 期)、籽粒建成期(R2 期)和乳熟期(R3 期)各处理功能叶可溶性糖质量分数的测定结果显示,VT 期数值均较低,在2.1%~2.8%;之后两次测定到的叶片中可溶性糖略有增加,在3.2%~4.7%,到R3 期基本保持稳定。
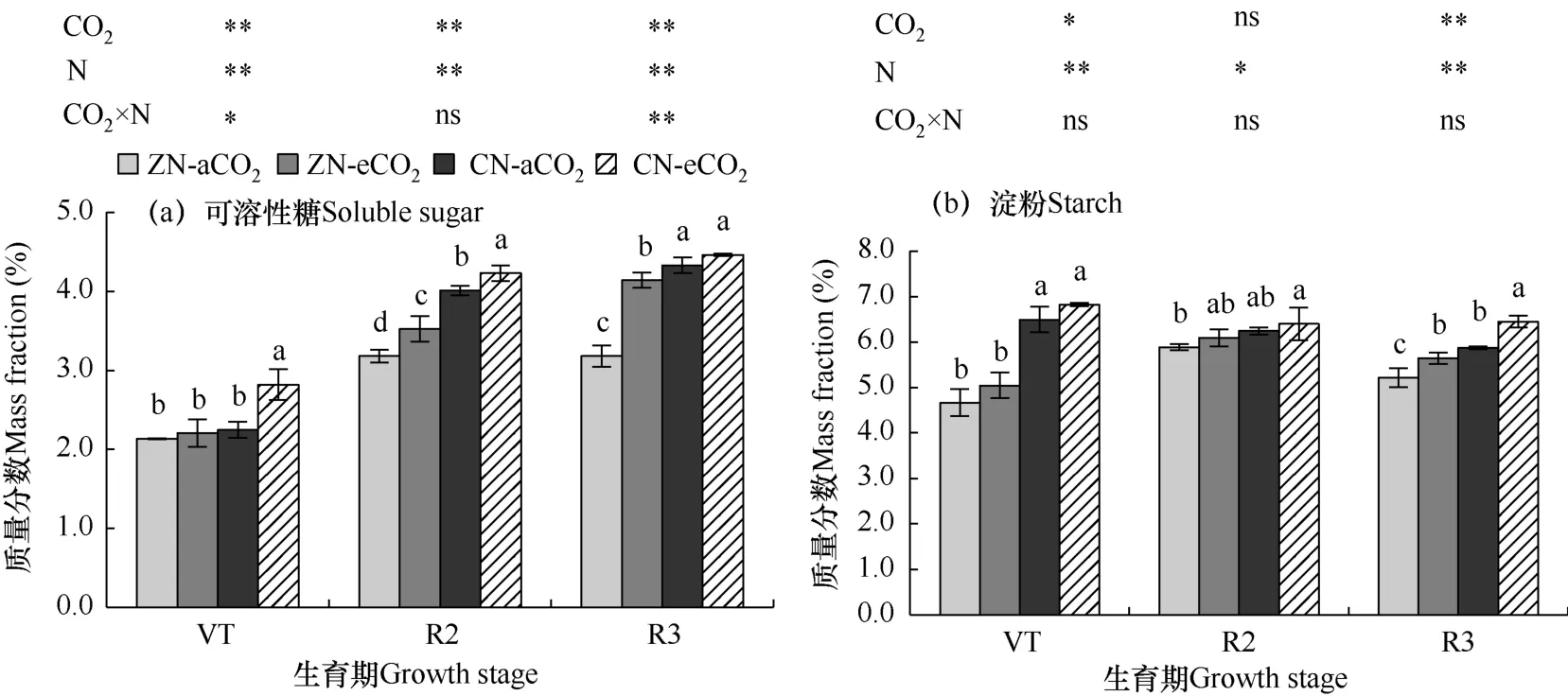
图3 不同处理夏玉米花后功能叶可溶性糖和淀粉质量分数Fig.3 Comparison of mass fractions of soluble sugar and starch in functional leaves of summer maize after flowering for different treatments
分别对各生育期不同处理进行单因素方差分析,结果表明,eCO2单一因素下功能叶可溶性糖质量分数相比aCO2处理显著增加(P<0.05)。其中VT 期eCO2处理在CN 水平下可溶性糖比aCO2处理显著提升25.6%;在R2 期,在ZN 水平下显著提升10.8%,CN水平下显著提升5.5%;在R3 期,ZN 水平下显著提升30.2%。单一eCO2下功能叶淀粉质量分数比aCO2处理增加。在R3 期两种施氮水平下分别显著增加8.2%和9.9%。在VT 和R2 期影响不显著。
氮素单一因素下,夏玉米功能叶可溶性糖质量分数相比无氮处理显著增加(P<0.05),其中VT 期显著增加16.8%;R2 期显著增加23.0%;R3 期显著增加20.1%。施氮同样也对功能叶淀粉质量分数增加有促进作用,在VT 期和R3 期增加显著(P<0.05),增幅分别为37.1%和13.4%。
对于双因素共同作用的处理(CN−eCO2),功能叶可溶性糖质量分数也较ZN−aCO2处理显著增加(P<0.05),是ZN−aCO2处理的1.35 倍。本试验条件下,eCO2和氮肥施用对玉米功能叶淀粉质量分数未显示显著的交互作用。
综上,eCO2和氮肥及其交互作用均会使夏玉米功能叶可溶性糖质量分数增加,两因素为相互促进作用,且eCO2的增加作用高于氮肥。eCO2和氮肥均会促进功能叶中淀粉质量分数的增加,但是交互作用不显著。
2.3 CO2浓度升高和氮肥互作对功能叶不同组分氮同化物的影响
2.3.1 可溶性含氮化合物
作物从土壤中吸收无机氮的形式为铵态氮和硝态氮,其中旱地农田土壤中速效氮的主要形式是硝态氮。作物吸收硝态氮后,在硝酸还原酶的作用下将硝态氮同化为铵态氮,随后进入氨基酸及蛋白质合成过程。游离氨基酸是植株氮素在合成结构性氮组分前的氮素过渡形态。可溶性蛋白是植株体内诸多生理活动所必需的酶的组成结构,对作物体内光合作用、物质代谢及物质转运等起着重要作用。
由图4 可见,各处理功能叶硝态氮质量分数从夏玉米抽雄期(VT 期)、籽粒建成期(R2 期)至乳熟期(R3 期)呈先升高后降低的趋势,VT 期硝态氮质量分数在532.4~730.91ug·g−1;R2 期较高,在824.7~915.3ug·g−1;R3 期硝态氮质量分数较低,在532.4~730.9ug·g−1。各处理功能叶游离氨基酸质量分数的测定结果表明,玉米从抽雄吐丝开始,功能叶游离氨基酸质量分数随生育进程的推进总体略呈降低趋势,范围在278.3~420.8ug·g−1。各处理功能叶单位叶面积可溶性蛋白质量分数的测定结果表明,在R3 期出现下降趋势,质量分数为1039.7~1358.5mg N·m−2。
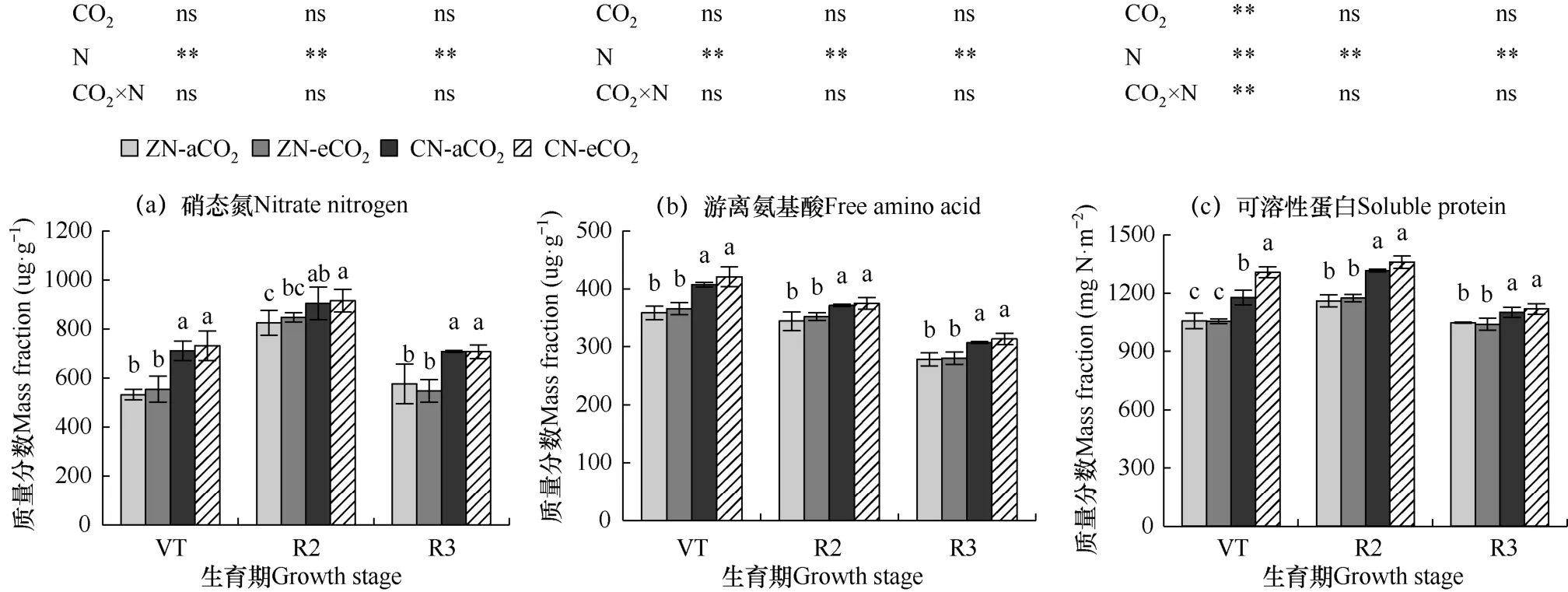
图4 不同处理夏玉米花后功能叶硝态氮、游离氨基酸和可溶性蛋白质量分数的比较Fig.4 Comparison of mass fractions of nitrate nitrogen,free amino acid and soluble protein in functional leaves of summer maize after flowering for different treatments
各生育期不同处理间单因素方差分析表明,eCO2单一处理下功能叶硝态氮和游离氨基酸质量分数比aCO2处理有增加趋势,但未达显著水平;单一eCO2处理下VT 期CN 水平下单位叶面积可溶性蛋白质量分数显著增加11.0%(P<0.05),其余处理差异不显著。氮素单一处理后夏玉米功能叶硝态氮、游离氨基酸质量分数和单位叶面积可溶性蛋白质量分数显著增加(P<0.05),CN 处理平均(CN−aCO2和CN−eCO2)分别比ZN(ZN−aCO2和ZN−eCO2)处理平均在VT 期、R2期和R3 期的功能叶硝态氮质量分数分别显著增加32.6%、8.8%和26.0%,游离氨基酸质量分数分别增加14.3%,7.2%和11.1%,单位面积可溶性蛋白质量分数分别增加17.7%、14.6%和6.3%。eCO2和氮肥施用对玉米功能叶硝态氮和游离氨基酸质量分数没有显示显著的交互作用,只在VT 期显示可溶性蛋白显著增加23.7%(P<0.05)。
因此,在本试验条件下,氮肥单独施用会显著增加夏玉米功能叶可溶性含氮化合物硝态氮、游离氨基酸和单位叶面积可溶性蛋白质量分数;eCO2单独作用对夏玉米功能叶硝态氮和游离氨基酸质量分数的增加有一定促进作用,但未达显著水平;eCO2也仅在CN 水平下显著增加抽雄期功能叶单位叶面积可溶性蛋白质量分数。交互作用显示,eCO2会削弱氮肥对功能叶硝态氮和游离氨基酸质量分数的增加作用;eCO2仅在VT期显示会显著促进氮肥增加功能叶单位面积可溶性蛋白质量分数的作用,后期作用不显著。
2.3.2 非溶性氮素化合物
与可溶性蛋白相比,类囊体氮和细胞壁氮属非溶性结构氮,在总氮供应不足的情况下,可溶性蛋白对作物各项生理活动的必要性比结构性氮(如细胞壁氮)更为迫切。细胞壁氮是作物细胞壁的主要构成组分,其在调节气孔导度、影响光合速率方面有一定作用。类囊体是叶绿体内光合作用的反应场所,类囊体氮素是光合器官氮素的组成部分。
根据图5,在夏玉米抽雄期(VT 期)、籽粒建成期(R2 期)和乳熟期(R3 期)对各处理功能叶细胞壁氮素和类囊体氮素单位叶面积质量分数测定结果表明,CN 水平下单位面积细胞壁氮质量分数在花后基本稳定在一定数值范围内,在330.8~393.5mg N·m−2。单位面积类囊体氮素质量分数花后呈现逐渐降低趋势,在342.2~139.7mg N·m−2范围内变化。各时段不同处理间单因素方差分析表明,eCO2单一作用会降低夏玉米单位功能叶面积内细胞壁氮素和类囊体氮素的质量分数,具体表现为eCO2水平下细胞壁氮素的降低在VT 和R2期都较为显著,在ZN 和CN 下分别比aCO2处理显著降低23.0%和11.0%(P<0.05);在R2 时期,ZN 和CN 水平下降低幅度分别为8.1%和15.5%;eCO2下类囊体氮素的降低在VT 时期和R3 时期CN 水平下较为显著,VT时期ZN和CN下分别显著降低21.0%和14.1%(P<0.05),R3 时期CN 水平下比aCO2处理显著(P<0.05)降低24.6%。氮肥单一作用会显著增加夏玉米功能叶单位面积非溶性氮素化合物质量分数(P<0.05),具体表现为在VT 期、R2 期和R3 期,CN 处理平均(CN−aCO2和CN−eCO2)分别比ZN(ZN−aCO2和ZN−eCO2)处理细胞壁氮素显著增加53.1%、24.4%和18.8%(P<0.05),类囊体氮素分别显著增加26.0%,27.3%和37.9%(P<0.05)。双因素方差分析结果显示,eCO2和氮肥施用对功能叶细胞壁氮仅在R2 时期有明显的交互作用,对类囊体氮素仅在R3 时期有正交互作用,其余时期交互作用不显著。
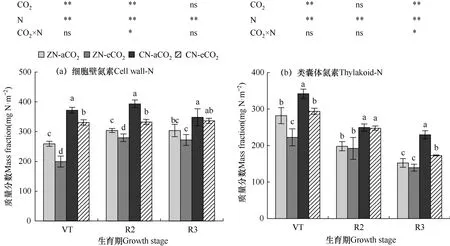
图5 不同处理夏玉花后单位面积功能叶细胞壁氮素(a)和类囊体氮素(b)质量分数的比较Fig.5 Comparison of the mass fraction of cell wall-N(a)of functional leaf and thylakoid-N(b)per unit area of summer maize after flowering for different treatments
可见,本试验条件下,eCO2处理玉米功能叶单位叶面积非溶性氮素化合物的质量分数减少,氮肥施用则无疑会促进非溶性氮化合物质量分数增加。两因素交互作用使R2 期细胞壁氮素和R3 期类囊体氮素质量分数增加。
2.4 CO2浓度升高和氮肥互作对叶片总碳、总氮和碳氮比的影响
叶片总碳包括叶片中所有形式的碳合成及代谢物,是结构性碳和非结构性碳的总量。叶片总氮也是指叶片内所有吸收及同化代谢物中的氮,包括非结构性氮(如硝态氮、氨基酸)和结构性氮(如可溶性蛋白、类囊体氮、细胞壁氮等),是叶片内氮素总体状况的反映。不同时期叶片碳氮比能反映植株体内碳氮养分总体合成或供应状况。
在夏玉米抽雄期(VT 期)、籽粒建成期(R2 期)和乳熟期(R3 期)各处理功能叶总碳质量分数的测定结果显示(图6),夏玉米花后功能叶总碳在41.4%~44.4%,从抽雄期开始随生育进程的发展,功能叶总碳总体呈现降低趋势。功能叶总氮质量分数从抽雄到完熟期也总体呈降低趋势,质量分数在3.2%~2.2%,功能叶内的养分逐渐向籽粒转移,叶片逐渐趋向衰老。碳氮比总体呈升高趋势,从抽雄期的13.8~16.7:1 增至乳熟期的16.1~19.3:1,表现为氮向籽粒转移。

图6 不同处理夏玉米花后功能叶总碳、总氮和碳氮比质量分数的比较Fig.6 Comparison of mass fractions of total carbon,total nitrogen and C/N ratio in functional leaves of summer maize after flowering for different treatments
各生育期不同处理间单因素方差分析结果表明,eCO2单一作用下功能叶总碳质量分数比aCO2处理增加。在VT 期,eCO2处理ZN 水平下总碳比aCO2处理显著升高1.7 个百分点(P<0.05);在R2 期CN 水平下,显著升高1.5 个百分点(P<0.05),其余时期eCO2对各处理的影响未达显著水平。eCO2单一作用下功能叶总氮质量分数总体略低于aCO2,各时期差异都未达显著水平。eCO2单一作用对功能叶碳氮比增加的影响主要在R2 时期ZN 水平下影响显著(P<0.05),显著增加7.2%。
氮肥单一作用对夏玉米功能叶总碳质量分数影响不显著,仅在R2 期比不施氮处理显著增加了1.1 个百分点(P<0.05)。单一氮素处理会显著增加各生育期功能叶总氮,各时期CN 处理比ZN 处理总氮质量分数高11.7%~17.0%(P<0.05)。单一氮素处理下C/N 在灌浆后期显著降低,降低幅度为9.8%~13.4%,归因于施氮对叶片总氮的增加效果显著(P<0.05)。
可见,本试验条件下,单一eCO2因素会促进夏玉米功能叶总碳质量分数的增加,而总氮质量分数降低,碳氮比增加。单一氮肥因素对夏玉米功能叶各生育期总碳质量分数的影响不显著,会显著增加功能叶总氮,降低功能叶碳氮比。二者交互作用对夏玉米功能叶总碳、总氮和碳氮比均无显著影响。
3 讨论与结论
3.1 讨论
3.1.1 CO2对玉米碳氮代谢及产量的影响
本研究显示,单因素eCO2可以促进夏玉米花后功能叶碳代谢增加,使得可溶性糖、淀粉和总碳质量分数在大部分时期显著增加,这与茶叶幼苗在eCO2条件下叶片可溶性糖[24−25]、淀粉和总碳增加[26]的研究结果一致,主要由于CO2是光合作用的底物,eCO2使作物净光合速率提高进而促进了光合产物可溶性糖的积累。
单因素eCO2下夏玉米花后功能叶可溶性含氮化合物指标硝态氮、游离氨基酸及可溶性蛋白的质量分数略微增加但不显著,这些指标均是作物生理过程中的简单氮组分。而一些非溶性氮素化合物(如细胞壁氮和类囊体氮)的质量分数则在大部分时期呈显著降低,这两类氮均为结构氮组分,eCO2下一些组成复杂的结构氮组分的合成可能受限,也可能与本研究中的氮用量(180kg N hm−2)依然不足有关。这与报道中eCO2下棉花苗期[27]和四季豆四叶期叶片[28]游离氨基酸提高、药用植物生长后期叶片可溶性蛋白增加[29]的结果一致。而类囊体是光合作用在叶绿体内进行的主要场所,本研究中eCO2处理中类囊体氮的质量分数在玉米花后有所降低,也可能与长期eCO2下的光合适应现象有一定关系,即淀粉和糖的过量积累引起类囊体与基粒的破坏。在多种作物[30−31]上的研究结果表明,eCO2对NSC 有促进效果,而对类囊体氮在后期有抑制效果,这与本试验的结果是一致的。
单一eCO2因素下功能叶总氮质量分数总体略低于aCO2,出现这种现象的原因可能是由于作物体内碳水化合物积累的增加从而稀释了总氮的质量分数,总体会使碳氮比增加,这种现象仅在R2 时期ZN 水平下有所体现。这也与非结构性碳水化合物在eCO2下增加及部分含氮化合物和总氮质量分数下降有关。该结果与一些研究对水稻在eCO2下植株地上部碳养分累积量增加、植株C/N 增加的结果一致[32−33]。
本试验条件下,单一eCO2会促进夏玉米地上部生物量的增加,这与美国伊利诺伊FACE 的研究结果[34]相似。本研究对产量组成要素的分析表明,eCO2在CN水平下显著增加了夏玉米穗重,但对其余产量构成因素及产量影响不显著。造成eCO2对玉米增产不显著的原因一方面可能是由于本试验中eCO2处理受病虫害影响的缘故。考种过程中发现,ZN 水平下aCO2和eCO2处理500 粒籽粒中受虫害侵染的玉米粒分别为1.2 粒和7.6 粒,eCO2下的虫害籽粒显著增多。对其它作物的实验也有类似报道,美国伊利诺伊州的SoyFACE 田间试验表明,eCO2下针对玉米及大豆的植食性害虫数量增加[35]。针对小麦的OTC 试验表明,eCO2下小麦蚜虫的繁殖量及相对生长率都提高[36]。针对C4作物谷子的OTC 试验也表明,eCO2下谷子灌浆期和收获期玉米螟数量显著增加,谷子穗重降低[37]。
总体来讲,本试验条件下,eCO2显著促进功能叶中碳同化物可溶性糖及淀粉的质量分数;eCO2对不同组分氮同化物的影响,主要体现在大部分时期结构氮组分(细胞壁氮和类囊体氮)质量分数显著降低,但并未在产量水平上有显著体现。
3.1.2 氮肥及其与eCO2互作对玉米碳氮代谢及产量的影响
单一氮肥因素会显著增加夏玉米多数生育期地上部生物量、穗粒重和产量,使得花后功能叶简单碳组分可溶性糖、灌浆前期和后期淀粉质量分数显著增加,对总碳的增加作用不显著。这与适当增加施氮量使玉米功能叶可溶性糖和淀粉增加[38]的其它研究结果一致。本研究中,氮肥施用对功能叶可溶性糖和淀粉促进作用在后期出现下降的原因,可能是由于碳水化合物主要由营养器官向籽粒转移,而在功能叶中的累积有所降低的缘故[39]。
单一氮素处理后夏玉米功能叶可溶性含氮化合物、非溶性氮素化合物和总氮质量分数均显著增加。这与关于施用氮肥会显著增加叶片硝态氮[40]、游离氨基酸和可溶性蛋白质量分数[41],增加细胞壁氮素和类囊体氮素[4]的相关研究结果一致。本研究结果表明,不施氮处理功能叶各项氮代谢指标在反映氮素供应及代谢方面均较为敏感,其中非溶性氮素化合物(平均降低31.3%)对于不施氮的敏感程度高于可溶性含氮化合物(平均降低15.4%)。也进一步说明结构性氮组分(细胞壁氮及类囊体氮)质量分数增加的滞后性。同时,这与有研究关于常规施氮和不施氮处理下,类囊体氮素动态变化在花后出现快速下降的时间均明显早于可溶性蛋白出现快速下降时间[42]的结果一致。单一氮素处理后C/N 在花后显著降低,降低幅度为9.8%~13.4%,说明施氮对叶片总氮的增加效果高于其对总碳的增加效果。
eCO2和氮肥的交互作用下夏玉米功能叶简单组分碳同化物−可溶性糖质量分数增加,两因素为相互促进作用,且eCO2的增加作用高于氮肥,而对功能叶淀粉质量分数则没有显著影响。针对棉花的OTC 试验表明,eCO2下植株体内氮素转化的2 个关键酶即硝酸还原酶和谷氨酰胺合成酶的活性都受到抑制,使氮素吸收后的转化及同化受阻;不过在氮素供应水平提高后,酶的活力也有所提升[43]。这可能是由于C4作物对CO2的亲和力高,因此,结构碳组分淀粉对外界eCO2及氮肥施用的交互作用反应不敏感[44]。
eCO2和氮肥交互作用在改进作物氮供应、形态构成和光合反应方面有一定潜在促进作用,表现为某些时期(如抽雄期)关键组分质量分数的增加。而在产量方面,CO2与氮肥的交互作用虽然显著增加了夏玉米穗重,但对产量没有显著影响。有研究显示,C4作物产量在水分及氮素都供应充足条件下,eCO2对产量并没有显示明显的促进作用。据国外FACE 试验结果的报道,eCO2对C4作物产量的作用在作物受胁迫条件下(如干旱)才显示其对产量的显著补偿作用[45]。本试验条件下eCO2夏玉米功能叶碳代谢显著增加而氮代谢中结构氮组分的质量分数在后期则略有降低;而本试验中氮肥使功能叶碳代谢略微增加、氮代谢显著增加;eCO2和氮肥的交互作用对总碳、总氮和碳氮比的影响不显著。本探索试验由于试验地等方面的限制没有设置更多氮水平梯度,未能全面反映二者交互作用对生物量和产量的多种情况。本研究结果显示,在大气CO2浓度升高条件下适当施氮对碳氮代谢协调及玉米生物量和产量及部分产量构成因素有促进作用,达到资源协调利用和高产优质的农业生产目标。
总体来讲,本试验条件下,氮肥施用对功能叶中不同组分的碳氮同化物及产量都有促进作用。eCO2和氮肥的交互作用在提高碳同化物质量分数方面,和单一CO2作用的效果类似;交互作用对促进氮同化物质量分数的作用只在抽雄吐丝期的可溶性蛋白方面作用显著,而后期作用不显著,交互作用对非溶性氮同化物的作用为负向,即eCO2条件削弱了氮肥对氮同化物的促进作用,可能由于eCO2下复杂氮化合物合成有一定不足或其它多种原因(病虫害)有关,因此,尽管eCO2下穗重增加,但有效籽粒产量并未增加。另外,eCO2对氮代谢物的作用也可能与试验供试品种对氮肥的敏感程度有关,需设置更多的不同氮肥梯度及氮效率品种验证eCO2下碳氮代谢及其产量反应。
本研究是对eCO2与氮素互作对C4作物不同组分碳氮同化物质量分数及动态的一些初步探索,还需要在不同地力水平及多氮肥梯度及针对不同氮效率品种开展更多关于eCO2和氮肥施用对结构氮组分的合成及不同器官(功能叶、老叶、茎、籽粒)中各氮素组分的质量分数及转运等生理过程及可能影响产量的病虫害等影响进行多方面研究,以针对性地给出生产角度的代谢调控策略。
3.2 结论
大气CO2浓度升高使C4作物夏玉米功能叶可溶性糖、淀粉和总碳质量分数显著增加;对氮同化物中的可溶性含氮化合物硝态氮、游离氨基酸和可溶性蛋白影响不显著;但eCO2使夏玉米大多数时期单位面积非溶性氮素化合物细胞壁氮素和类囊体氮素显著减少、结构性氮化合物的合成受到一定影响,从而使总氮质量分数降低,碳氮比增加。但eCO2对夏玉米地上部生物量和产量均未显著影响。氮肥施用显著增加了玉米地上部生物量和产量,以及功能叶中碳同化物主要指标可溶性糖、淀粉和氮同化物指标,使碳氮比显著降低,产量提升。
eCO2和氮肥互作下,夏玉米功能叶碳同化物中可溶性糖质量分数增加,两因素为相互促进作用;而对功能叶淀粉质量分数则无显著影响。eCO2和氮肥互作也显著增加了夏玉米氮同化物中可溶性蛋白和一些时期结构性氮化合物如细胞壁氮和类囊体氮的质量分数和穗重,对协调碳氮代谢和产量提升提供了可能潜力,使后期碳氮代谢向协调方向发展,对生物量和产量的增加提供了可能潜力。
在未来大气CO2浓度升高条件下,需根据地力合理调控碳氮代谢生理合成过程,以协调碳氮代谢,保证玉米高产稳产。
