烤烟叶片光合速率日变化特征及其影响因素分析
2021-09-16余凌翔鲁韦坤张加云
余凌翔,鲁韦坤,张加云, 朱 勇,逄 涛,赵 泽
(1.云南省气候中心,昆明 650034; 2.云南省烟草农业科学研究院,云南 玉溪 653100)
引 言
烟叶既是烟草(Nicotianatabacum)的营养器官,又是其经济器官。烟株的生长发育和产量品质的形成,取决于烟草植株个体与群体的光合作用。因此,光合作用是烟草产量和品质形成的基础[1]。作物的光合作用对环境条件变化的敏感性,不仅受自身生理特性的影响,还受品种、气候、土壤等环境因子的影响[2-8]。目前,有关不同烤烟品种光合特性的研究已有不少报道。张振等[9]通过田间盆栽试验,研究复合盐对烤烟光合特性的影响,认为经过0.0683 mol·kg-1复合盐浓度处理可提高烟叶的光合作用性能,促进烤烟生长。曹阳等[10]认为,合理的种植密度有利于优化烟株形态,协调烟叶光合速率,提高烤烟成熟度。王子腾等[11]研究发现,高温强光胁迫下,成熟期烤烟的蒸腾速率、气孔导度和净光合速率均显著降低,胞间CO2浓度升高,光合能力下降。然而烟草净光合作用与生理生态因子关系的研究,多在相同生境或单一烟草品种中进行,不同生境下不同烟草品种光合特性的差异及其主要影响因子研究还未见报道。由于不同烟草品种在不同生态区的质量差异较大[12-13],同一品种在各地表现也不可能一致,因此只有将烤烟品种自身特性与当地生态环境条件结合起来,才能发挥优势品种的生产潜力[14]。本文通过分析8个品种烤烟在昭通市3个植烟区的叶片光合速率日变化特征及其影响因素的关系,为昭通市提高烟叶质量、打造区域特色烟叶品牌提供科技支撑。
1 材料与方法
1.1 试验区域概况
昭通市位于云南省东北部,是云南传统主产烟区之一,属亚热带、暖温带共存的高原季风立体气候。全年平均气温15.4 ℃,年日照时数1307.1 h,年降水量869.5 mm。其中烤烟大田生长期(5-9月)平均气温21.6 ℃,累计日照时数673.4 h,累计降水量622.2 mm。昭通市植烟区的海拔高差大,各地烟叶品质风格特点明显不同。在全市范围内选取昭阳区布嘎乡花鹿坪村、镇雄县坡头镇堰塘村和巧家县白鹤滩镇水井湾村3地为试验点,分别代表昭通市3种不同植烟区:昭鲁平坝烟区、东北部烟区和金沙江干热河谷烟区。3地的海拔、土壤、气温、降水量和日照时数均存在明显差异,3个试验点基本情况见表1。历年和2011年烟草大田生长期气象条件见表2。
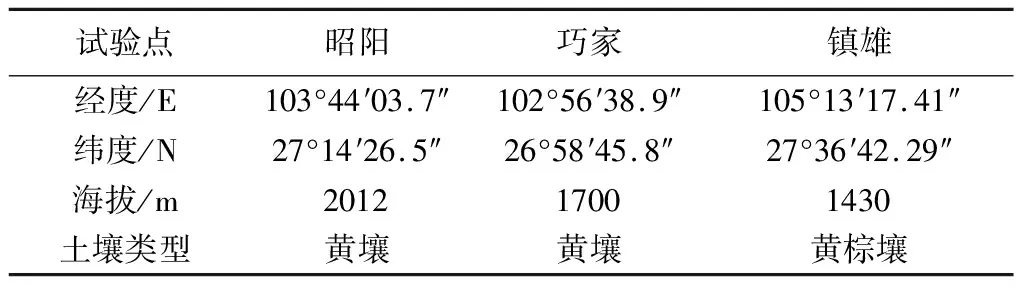
表1 试验点基本情况
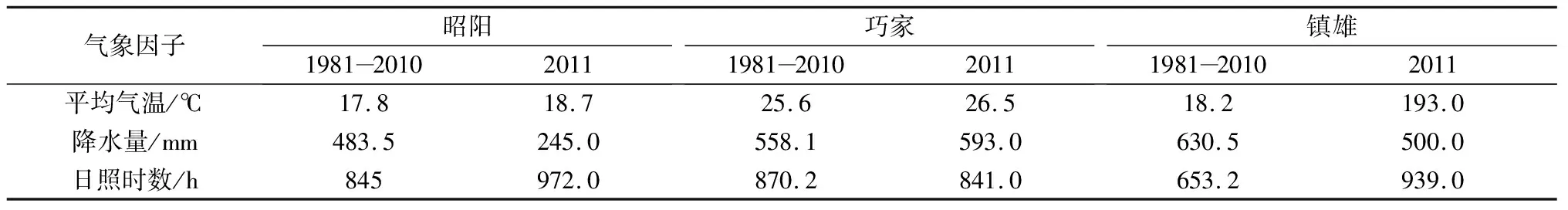
表2 试验点历年和2011年烟草大田生长期(5-9月)主要气象因子
1.2 试验材料与处理
根据昭通市烟草种植历史和品种更新计划,选择历史主栽品种、当前主栽品种和计划引进品种(K326、云烟87、云烟97、云烟99、云烟203、KRK26、红花大金元(红大)和PVH19)作为供试烟草品种。其中,红大是历史主栽品种,K326、云烟87是当前主栽品种,云烟97、云烟99、云烟203、KRK26、PVH19为计划引进品种。采用随机区组试验,共3个区组。昭阳小区面积36.3 m2,前茬作物为玉米,2011年5月10日移栽,株行距为0.55 m×1.1 m。巧家小区面积25.9 m2,前茬作物为小麦,2011年5月15日移栽,株行距为0.6 m×1.2 m。镇雄小区面积26.0 m2,前茬作物为小麦,2011年5月12日移栽,株行距为0.6 m×1.2 m。各试验点大田管理按优质烤烟管理工作方案执行,各项操作均保持一致。
1.3 光合作用测定
在烟草成熟初期(7月),在晴天选取同一品种长势一致、健康的烟草植株3株,以叶位接近、叶色一致、向阳的叶片作为测定对象。采用Li-6400便携式光合测定系统,测定3个试验点烟草在自然环境下中部叶片的光合日变化特征(08:00-19:00),包括净光合速率(Pn,μmol·m-2·s-1)、蒸腾速率(Tr,mmol·m-2·s-1)、气孔导度(Gs,mol·m-2·s-1)和胞间CO2浓度(Ci,μmol·mol-1)。记录太阳光合有效辐射(PAR,μmol·m-2·s-1)、空气CO2浓度(Ca,μmol·mol-1)、空气温度(Ta,℃)、空气相对湿度(RH,%)和叶面饱和水汽压亏缺(VPD,hPa)。每个品种测定重复3次,取平均值。
根据烟草生长进度,在同一生育期内,按照昭阳、巧家、镇雄的顺序,每个试验点在08:00-19:00内,逐小时对8个烤烟品种进行测量。
1.4 数据统计分析
数据经Microsoft Excel处理作图,方差分析、相关性分析、通径分析等在PASW 18.0和DPS数据处理系统中进行。
2 结果与分析
2.1 不同品种烟叶光合和环境因子日变化特征
在昭阳试验点,各品种净光合速率日变化过程为明显的“双峰”型(PVH19为“单峰”型)。第一峰值出现在09:00,之后净光合速率迅速下降,12:00-13:00“午休”过后,净光合速率开始上升,14:00或15:00出现第二个峰值,但此时峰值明显小于第一次峰值。PVH19在13:00净光合速率最大,之后急剧下降。
在巧家试验点,各品种烟叶的净光合速率日变化过程存在一些差异。PVH19的净光合速率在09:00达日最大值,10:00-11:00下降,净光合速率日变化波幅在10 μmol·m-2·s-1左右。云烟87、云烟97、云烟203和云烟99的净光合速率呈明显的“三峰”型变化,峰值分别出现在08:00、10:00和14:00(云烟203最后一个峰值出现在12:30)。云烟87、云烟97、KRK26和云烟99在11:00-12:30有明显“午休”现象,红花大金元“午休”现象出现在12:30,K326、云烟203“午休”现象出现在11:00(图1)。
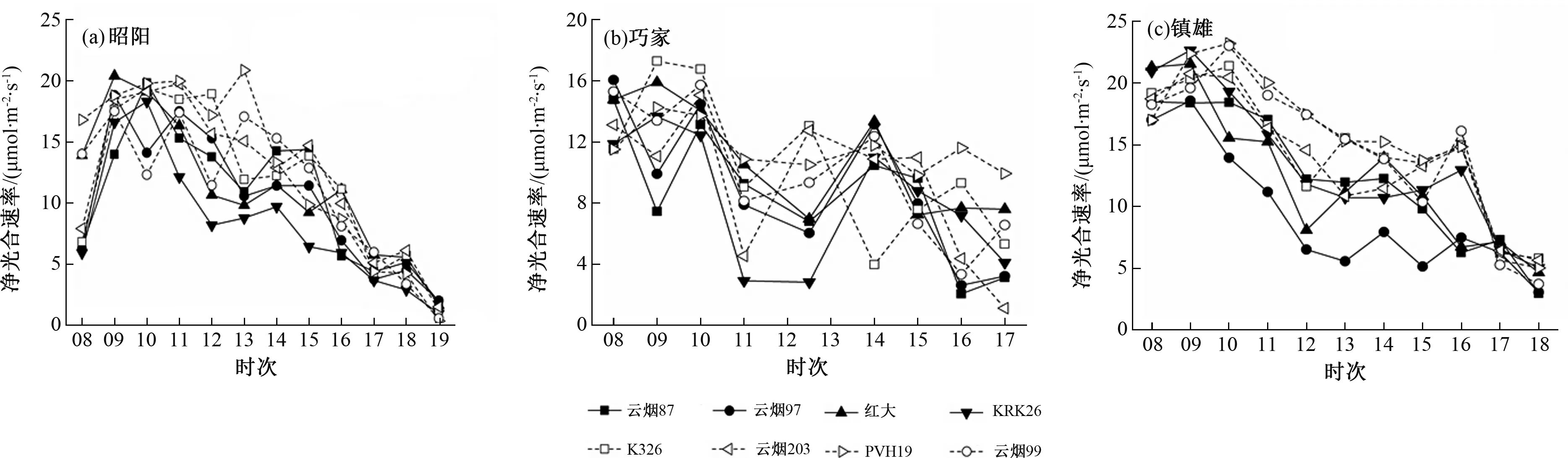
图1 3个试验点不同烟草品种净光合速率日变化比较
在镇雄试验点,各品种净光合速率日变化过程呈“双峰”型,但下降趋势明显。第一个峰值出现在08:00-10:00,之后则逐渐下降。KRK26、K326、云烟203、PVH19和云烟99第二个峰值出现在16:00,其他3个品种第二个峰值出现在14:00(图2)。
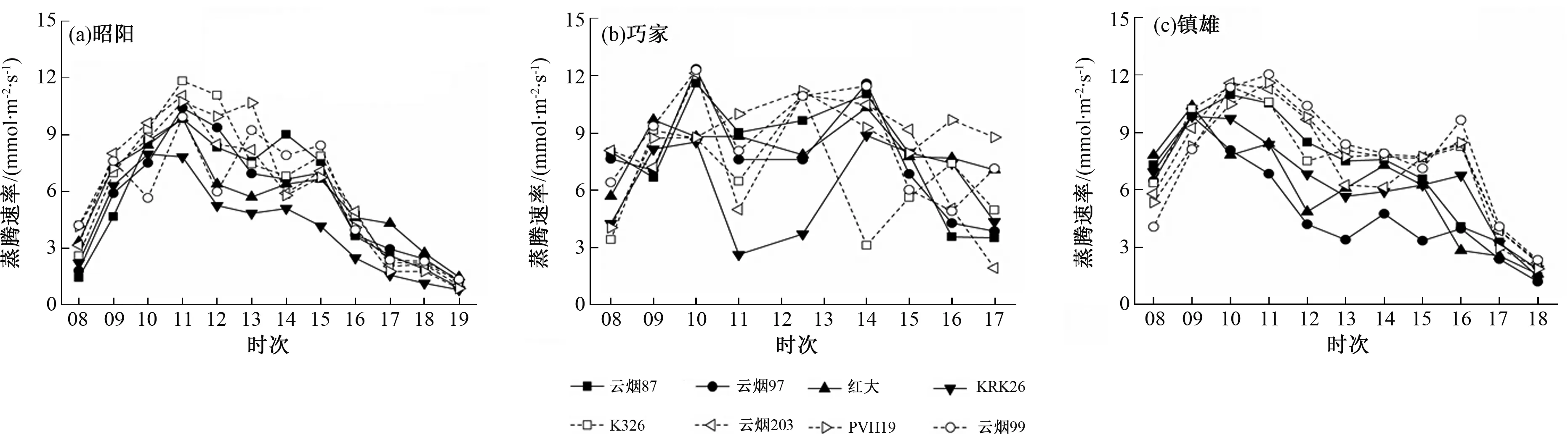
图2 3个试验点不同烟草品种蒸腾速率日变化比较
3个试验点各烟草品种的蒸腾速率日变化与净光合速率变化相似。测定期间,3个试验点环境因子有显著差异,镇雄的PAR较强(图3);巧家的VPD最高,其次是镇雄和昭阳的(图4);巧家和镇雄的Ta较高(图5)。
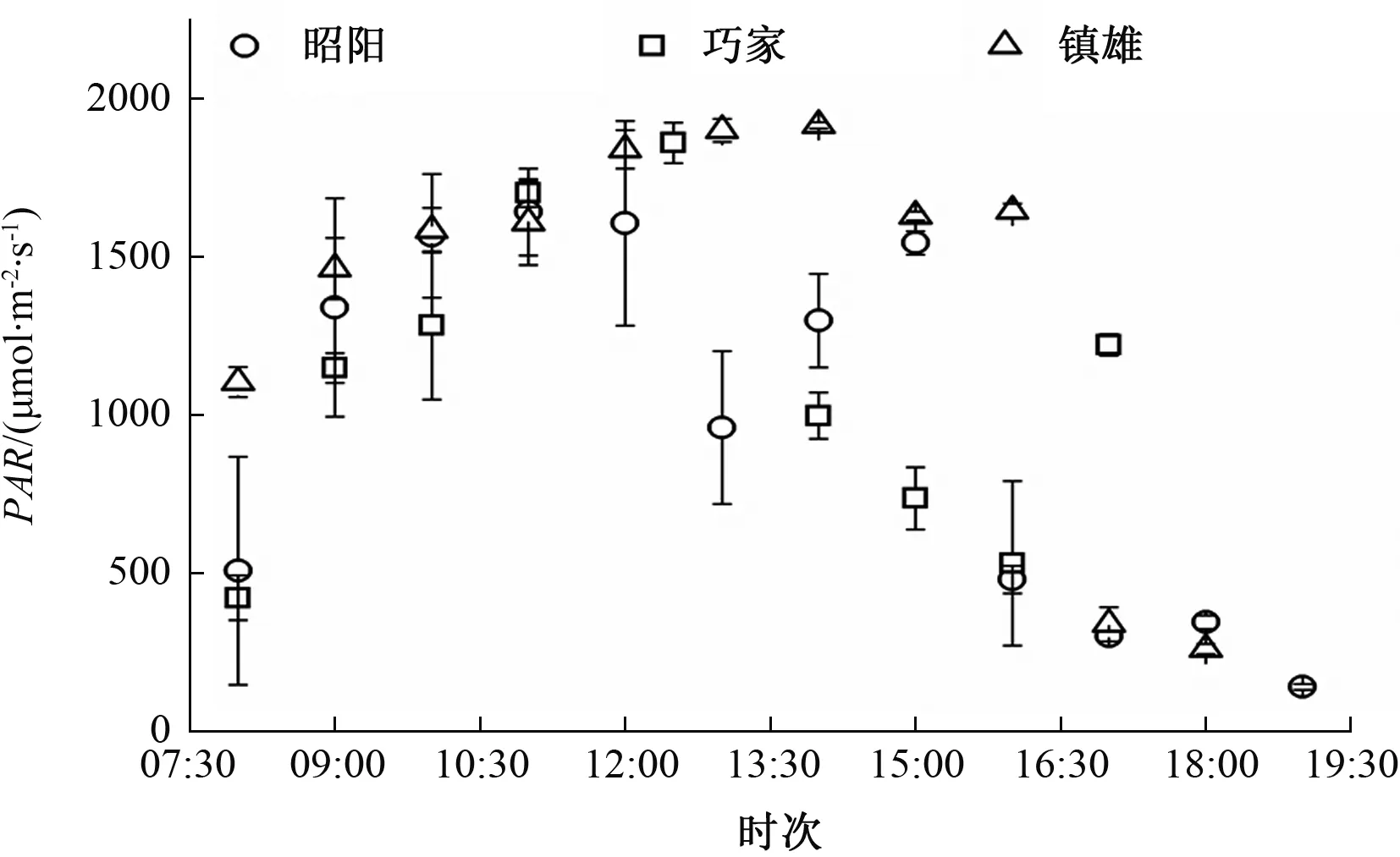
图3 3个试验点PAR日变化比较
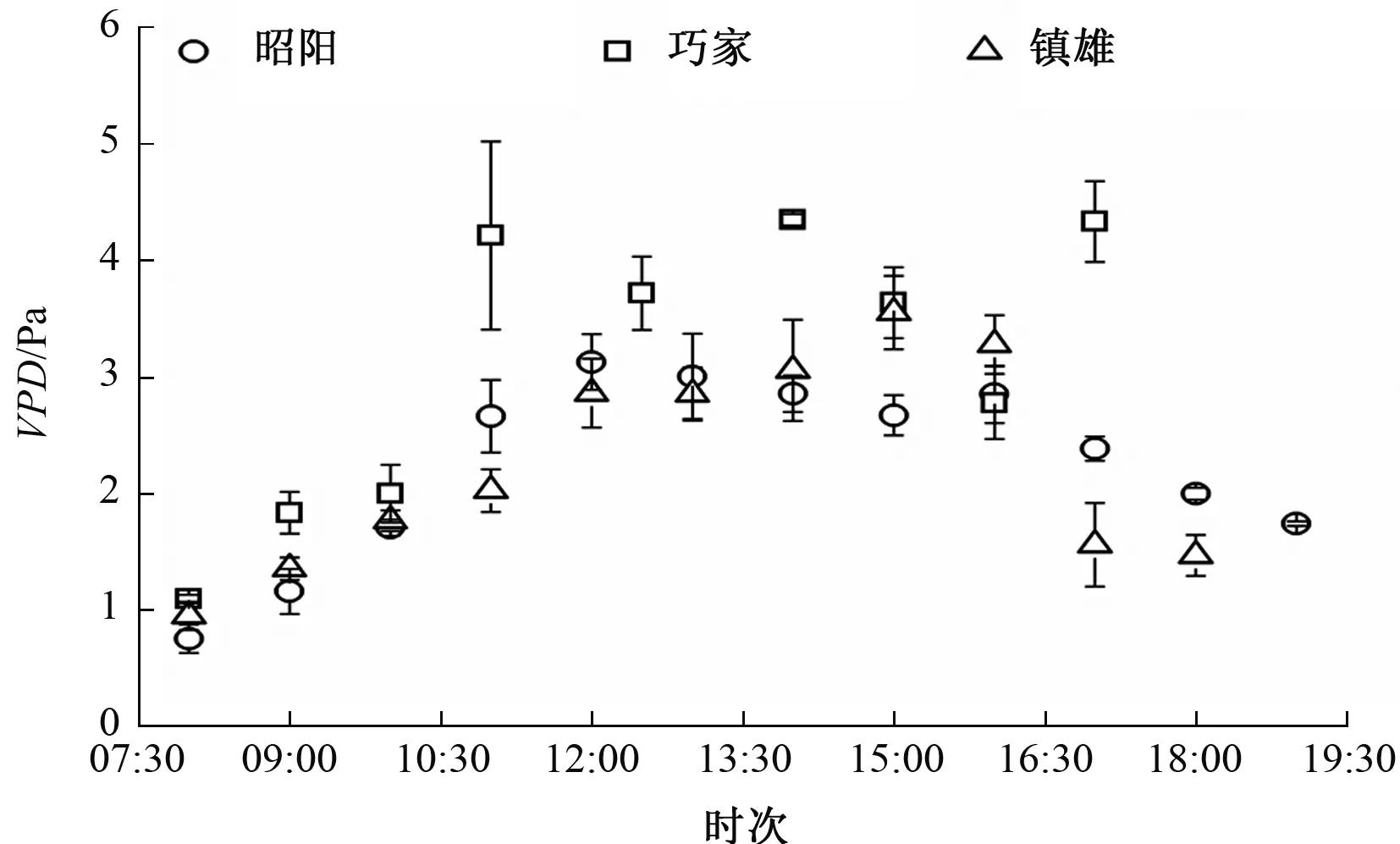
图4 3个试验点VPD变化比较
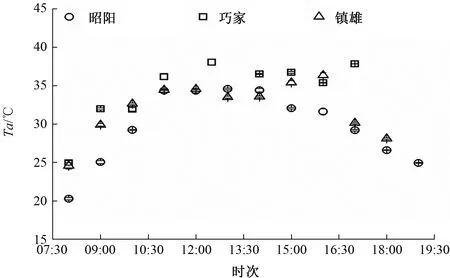
图5 3个试验点气温日变化比较
从各地烟叶净光合速率最大值和平均值(见表3)计算结果可见,昭阳PVH19净光合速率的日最大峰值最大,为20.88 μmol·m-2·s-1,云烟99为17.51 μmol·m-2·s-1,两者差异显著。巧家各品种净光合速率的日最大峰值范围为13.67~17.30 μmol·m-2·s-1,品种之间未达到显著水平。镇雄云烟87和云烟97净光合速率的日最大峰值分别为18.47 μmol·m-2·s-1和18.56 μmol·m-2·s-1;PVH19、云烟99和KRK26的净光合速率的日最大峰值为22.63~23.20 μmol·m-2·s-1,显著高于云烟87和云烟97的。各品种在不同生态环境下的表现也不同,昭阳云烟87和云烟97净光合速率最大峰值最高,其余各品种的则在镇雄最高。
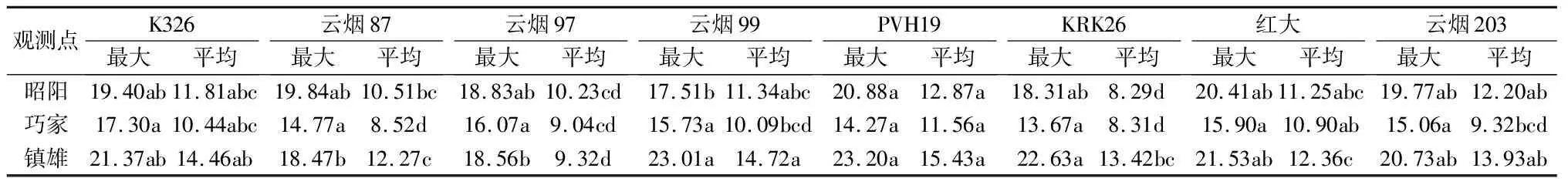
表3 昭阳、巧家和镇雄净光合速率的日最大值(Pm)和平均值(Pa) μmol·m-2·s-1
昭阳各品种净光合速率日平均值PVH19的>云烟203的>K326的>云烟99的>红花大金元的>云烟87的>云烟97的>KRK26的。昭阳PVH19净光合速率日平均值与云烟87、云烟97和KRK26的差异显著,云烟203的与云烟97和KRK26的差异显著,KRK26的仅与云烟97的没有显著差异。巧家各品种净光合速率日平均值PVH19的>红花大金元的>K326的>云烟99的>云烟203的>云烟97的>云烟87的>KRK26的。PVH19净光合速率日平均值与除红花大金元和K326以外的品种的差异显著,红花大金元的与云烟87、云烟97和KRK26的差异显著,K326的与云烟87和KRK26的差异显著,与其他品种的差异不显著。其他4个品种之间净光合速率日平均值差异不显著。镇雄各品种净光合速率日平均值PVH19的>云烟99的>K326的>云烟203的>KRK26的>红花大金元的>云烟87的>云烟97的。PVH19、云烟99、K326和云烟203这4个品种之间净光合速率日平均值没有显著差异,但PVH19和云烟99的与KRK26、红花大金元、云烟87和云烟97之间的差异显著。K326的和云烟203的与红花大金元、云烟87和云烟97之间的差异显著。红花大金元和云烟87之间净光合速率日平均值差异不显著,二者与KRK26的差异也不显著,但与云烟97的差异显著。云烟97净光合速率日平均值在昭阳最高,其余品种的在镇雄最高。
2.2 烟草净光合速率的生理因子影响分析
在昭阳、巧家和镇雄试验点,各烟草品种净光合速率与气孔导度Gs均呈极显著正相关关系(P<0.01),气孔导度Gs是影响烟草净光合速率的关键生理因子(见表4)。
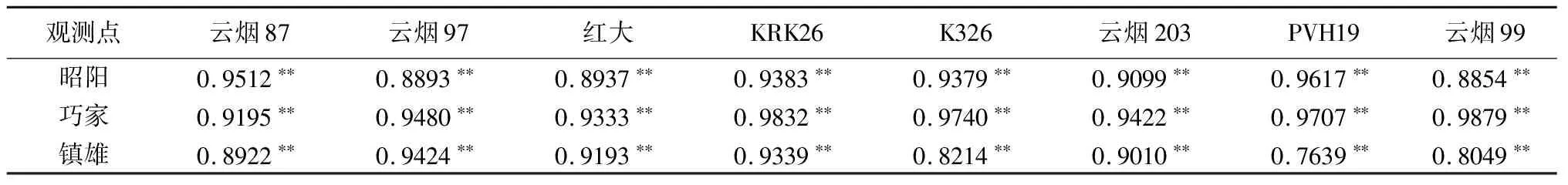
表4 昭阳、巧家和镇雄烟草净光合速率与生理因子的相关性分析
昭阳各品种叶片净光合速率和气孔导度的相关性PVH19的>云烟87的>KRK26的>K326的>云烟203的>红花大金元的>云烟97的>云烟99的,巧家各品种叶片净光合速率和气孔导度的相关性云烟99的>KRK26的>K326的>PVH19的>云烟97的>云烟203的>红花大金元的>云烟87的,镇雄各品种叶片净光合速率和气孔导度的相关性云烟97的>KRK26的>红花大金元的>云烟203的>云烟87的>K326的>云烟99的>PVH19的。红花大金元叶片净光合速率和气孔导度的相关性在3地相差不大;K326、云烟99的净光合速率与气孔导度的相关性在巧家的>昭阳的>镇雄的;KRK26、云烟203的净光合速率与气孔导度的相关性在昭阳和镇雄的基本一致,均小于巧家的;PVH19的净光合速率与气孔导度的相关性在巧家和昭阳的基本一致,均大于镇雄的;云烟87的净光合速率与气孔导度的相关性在巧家和镇雄的基本一致,均小于昭阳的;云烟97的净光合速率与气孔导度的相关性在巧家和镇雄的基本一致,均大于昭阳的。
2.3 烟草净光合速率与环境因子间的关系
2.3.1 相关分析
3个试验点影响烟草净光合速率的环境因子存在差异(见表5)。在昭阳,各烟草品种净光合速率与PAR之间有极显著正相关关系,K326、云烟203和云烟99净光合速率还与Ta呈显著或极显著正相关。在巧家,PVH19净光合速率与各环境因子之间相关均不显著,而其余各品种净光合速率与VPD呈显著或极显著负相关。云烟97、红花大金元和KRK26净光合速率还与Ta呈显著负相关,而红花大金元和KRK26净光合速率还与RH呈显著正相关。在镇雄,云烟203净光合速率与各环境因子相关不显著,云烟87、K326、PVH19和云烟99净光合速率与PAR正相关显著或极显著。
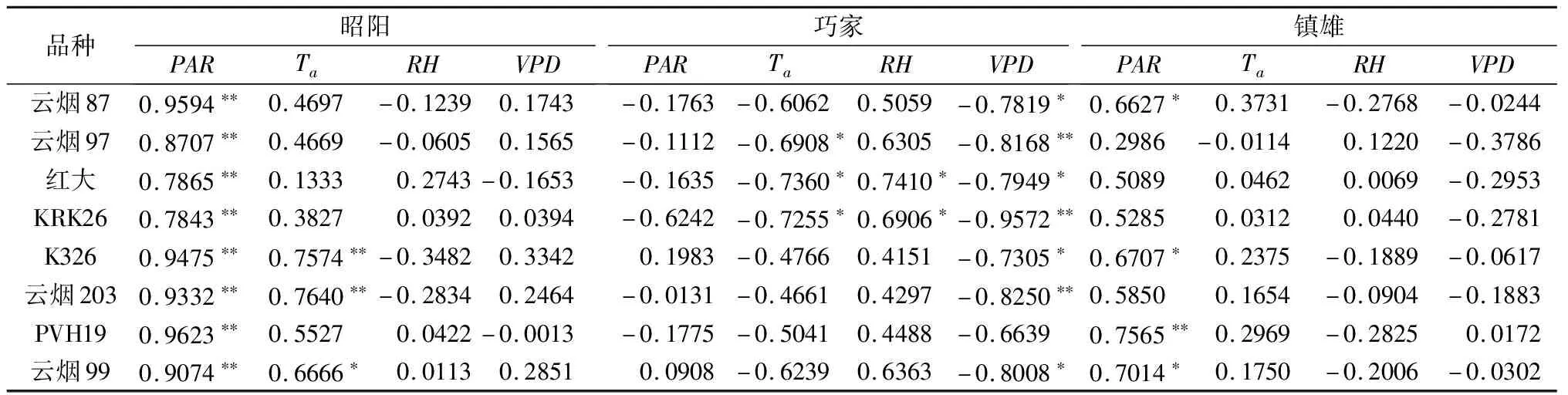
表5 昭阳、巧家和镇雄烟草净光合速率与环境因子的相关系数
2.3.2 决策系数
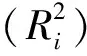
(1)
其中,Pi为自变量i对因变量的直接通径系数,riy为自变量i与因变量的Pearson相关系数。
把各自变量对因变量的综合作用进行排序,最终确定主要决策变量和限制变量[15-16]。决策系数最大的变量为主要决策变量,而决策系数最小且为负值的变量为主要限制变量。
各烟草品种叶片净光合速率在昭阳、巧家和镇雄三地的决策因子、限制因子是不同的(见表6)。在昭阳,各烟草品种净光合速率的主要决策因子均为PAR,限制因子均为VPD,这与该地烤烟大田生长期光照充足、降水量相对较少的气候特点一致。在巧家,各品种的表现差异较大,决策因子主要是RH,限制因子主要是Ta,这与该地烤烟大田生长期平均气温较高、空气相对干燥的气候特点一致。在镇雄,决策因子主要是PAR,而限制因子主要是VPD,这与当地烤烟大田生长期平均气温较低、降水相对较多的气候特点是一致的。
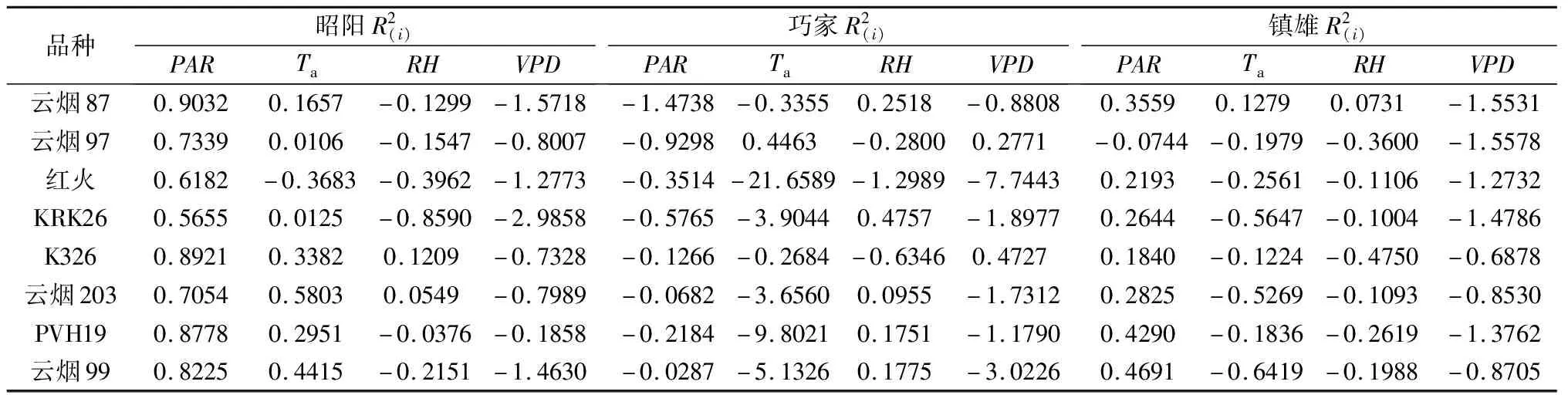
表6 昭阳、巧家和镇雄烟草净光合速率与环境因子的决策分析
3 结果分析
3.1 不同地点、不同烤烟品种的叶片光合比较
单位土地面积上一个生长季的总收获是光合速率、光合面积、光合功能期、呼吸消耗和经济系数等多种因素共同影响的结果。光合作用是植物生理的核心功能,可用来表征植物的健康状况与活力[17]。植物干物质90%以上来自光合作用同化的CO2,对于以叶片为主要收获对象的烟草来说,光合生产力与产量的关系更为密切。
叶片净光合速率的日变化,反映了一天中光合作用的持续能力。试验结果表明,各烟草品种无论净光合速率日最大峰值还是日平均值,镇雄的较高,其次为昭阳的,在巧家的较低。说明各烟草品种对镇雄生态环境的适应性比昭阳和巧家的强,对巧家生态环境的适应性最差。镇雄烤烟大田生长期平均气温19.3 ℃,累计降水量500 mm,累计日照时数939 h,相对较高的气温、较多的降水和光照有利于烟草生长。昭阳烤烟大田生长期平均气温18.7 ℃,累计降水量245 mm,累计日照时数972 h,各烟草品种净光合速率的主要决策因子均为PAR,限制因子均为VPD,烤烟大田生长期光照充足,但降水不足限制了烟草的气孔导度。巧家烤烟大田生长期平均气温26.5 ℃,累计降水量593 mm,累计日照时数842 h,各烟草品种净光合速率的决策因子主要是RH,限制因子主要是Ta,该地平均气温高,空气相对干燥、光照相对不足对烟草的光合速率产生了一定限制作用。
不同基因型烟草本身光合特性存在较大差异[14],但也受生态环境的影响。试验中,烟草净光合速率的品种与环境互作效应明显。PVH19、云烟99和K326在3个试验点的日平均净光合速率均较高,在3个试验点的适应性都较强;云烟87和云烟97在3个试验点的日平均净光合速率均较低,不太适合在3地栽培。红花大金元、云烟203和KRK26在不同生态环境下表现差异较大,其中KRK26在昭阳和巧家表现最差,在镇雄表现较好,表明相对较高气温、较多降水和光照有利于KRK26生长;云烟203生长在昭阳表现较好,但在巧家和镇雄表现较差,表明云烟203需要较强的光照,并且对干旱有一定耐受能力;红花大金元在巧家表现较好,但在昭阳和镇雄表现较差,说明高温环境有利于红花大金元生长。
3.2 生理因子对烟草净光合速率的影响
植物净光合速率与气孔导度、蒸腾速率、胞间CO2浓度等之间的关系因植物种类和研究条件(环境、气候等)而不同[18-20]。气孔导度对植物光合作用有重要的调节作用[21-22],当气孔行为对净光合速率起明显的主导控制作用时,Pn、Tr和Gs三者具有较好的协同响应[23]。试验结果表明,3个试验点各烟草品种Pn与Gs极显著正相关,说明气孔因子是影响各试验点烟草品种净光合速率的主要生理因素,3个试验点烟草各品种均是气孔决定型[24]。
3.3 环境因子对烟草净光合速率的影响
叶片净光合速率不仅与自身因素密切相关,还受光照强度、温度、CO2浓度、空气相对湿度、土壤水分等的共同影响,且各种环境因子对光合作用的影响因植物种类和生态环境而有所差别[25-28]。
试验中,昭阳各烟草品种的Pn与PAR有极显著正相关,Ta与Pn呈正相关,部分品种相关性达显著水平。决策分析结果表明,PAR是Pn的主要决策变量,VPD是Pn的主要限制变量,说明在昭阳较强的PAR对各烟草品种的Pn有促进作用,这与张广富等[5]的研究结果一致。较大的VPD可能增加了烟叶气孔和非气孔限制因素,从而对Pn的提高不利。在巧家,VPD与各烟草品种的Pn呈显著负相关,RH与Pn呈正相关,而PAR和Ta与Pn呈负相关。决策系数表明,RH对多数品种的Pn起决定作用,说明RH的增加促进了Pn的提高,与薛义霞等[29]对番茄的研究结果一致;高温和强光环境抑制了烟草的光合作用[30]。Ta是Pn的主要限制变量,可能该地较高的气温已对烟草净光合速率产生了抑制作用,导致其净光合速率日最大峰值和日平均值均较低。在镇雄,PAR与Pn呈显著正相关,PAR也是Pn的主要决定性变量,VPD和RH起限制作用,与昭阳的相似。但是,其较高的温度可能对光合酶活性也有一定的抑制作用,从而增加了非气孔限制因素。
虽然Pn的日变化仅能表征很短一段的环境因子对烤烟生长的适宜程度,但仍能为揭示不同生境下不同烟草品种的光合特性及其与环境因子的关系提供丰富的信息,这些信息可在烟草合理选址布局中发挥重要的指导作用。
4 结论与讨论
4.1 结 论
(1)昭阳烟区和镇雄烟区各烟草品种的Pn日变化均为“双峰”型;在巧家烟区,云烟87、云烟97、云烟203和云烟99的Pn日变化为“三峰”型,其他品种Pn日变化为“双峰”型。
(2)3个试验点PVH19、云烟99和K326的Pn日平均值均较高,云烟87和云烟97的均较低,说明PVH19、云烟99和K326在昭阳、巧家和镇雄3个烟区的适应性较强,云烟87和云烟97在这3个烟区的适宜性较差。KRK26、云烟203和红花大金元在不同生态环境下表现差异较大,KRK26在镇雄表现较好,云烟203在昭阳表现较好,红花大金元在巧家表现较好。
(3)3个试验点各品种烟草的Pn与Gs有显著正相关,各品种烟草的光合作用均是气孔决定型。
(4)在昭阳,PAR和Ta是烟草Pn的主要决策变量因子,VPD和RH是主要的限制变量因子;在巧家,RH对多数品种的Pn起决策作用,PAR、Ta和VPD是主要限制变量因子;在镇雄,PAR是Pn的主要决策变量因子,VPD、RH和Ta是主要限制变量因子。
4.2 讨 论
烤烟具有广泛的生态适应性,同时对生态条件的变化较为敏感。本试验为优质烤烟生产和发展提供了第一手参考资料,可用于验证烤烟气候区划结果的准确性,提高区划的精细化程度,实现动态区划。
光合作用是烟草产量形成的基础,不同气候区、不同烟草品种的光合作用日变化观测可为烤烟气象灾害监测和定量评价提供依据。若采用最新的田间光合作用监测技术系统,在不影响植物正常生长的条件下,对植物光合生理指标进行长期、多点、连续监测,并同步监测田间多种环境因子,可精确反映环境因子的变化对植物光合生理状态的动态影响。上述观测数据与气象综合数据库、精细化数值预报系统、烤烟气象灾害致灾指标相结合,可为烤烟栽培、灌溉决策、气象灾害防控提供精细化服务。
随着现代卷烟工业逐步走向精细化,为满足卷烟工业企业烤烟原料精准采购需求,需从卷烟工业企业的采购需求出发,加强生态因素对烤烟品质的影响研究,为不同烟区种植品种和品质筛选提供依据。
