毛乌素沙地不同地下水位埋深下土壤水补给特征及影响因素
2021-09-15裴艳武黄来明邵明安李荣磊张应龙
裴艳武,黄来明,邵明安,3,4,李荣磊,张应龙
(1. 中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室 北京 100101;2. 西北农林科技大学资源环境学院 杨凌 712100;3. 中国科学院大学资源与环境学院 北京 100049;4. 中国科学院水利部水土保持研究所黄土高原土壤侵蚀与旱地农业国家重点实验室 杨凌 712100;5. 陕西省神木县生态协会 神木 719399)
0 引 言
在干旱半干旱地区,土壤水是控制植被结构和生态系统演化的主要限制因子[1-2],而土壤水分供需平衡是维持土壤-植被系统健康发展的基础[3]。降水和地下水作为土壤水的潜在补给来源,其时空变化对土壤水分的供给和剖面分布有着重要的影响。因此,研究干旱半干旱地区土壤水的补给与转化特征对于优化区域水资源管理和促进生态系统可持续发展具有重要意义[4]。
土壤水中氢氧稳定同位素组成(δ2H和δ18O)受大气降水及水分在土壤中的蒸发、入渗等多种因素影响[5]。此外,一些存在浅层地下水的地区土壤水氢氧同位素组成及其剖面分布还受地下水位(Groundwater Level,GWL)季节波动的影响[6]。同位素技术作为“指纹”为辨识土壤水分的补给来源和运移规律提供了有效手段。与传统方法相比,氢氧稳定同位素作为示踪剂具有灵敏度高和准确性好等特点,对于水分补给来源具有良好的指示作用[7]。近年来,随着氢氧稳定同位素技术的发展与应用,众多学者对土壤水氢氧同位素分布特征进行了研究,为揭示区域土壤水的补给来源与转化规律提供了依据。田立德等[8]对青藏高原中部那曲地区不同深度土壤水稳定同位素变化规律进行了研究,结果表明表层土壤水δ18O受降水影响最为明显,而随着土壤深度的增加,土壤水δ18O受地下水的影响逐渐增强。张小娟等[9]通过对元阳梯田水源区降水及其4种典型植被类型(乔木林、灌木林、荒草地和荒地)下土壤水δ2H和δ18O进行分析,揭示了研究区土壤水的下渗机制。结果表明:乔木林和荒草地由于受到优先流的影响,深层(80~100 cm)土壤水δ18O要高于表层(< 40 cm)土壤,而灌木林与荒地受活塞流下渗的影响,深层(80~100 cm)土壤水δ18O低于表层(< 40 cm)土壤。Gazis等[10]分析了美国汉诺威地区附近6个不同地点降水和不同土地利用方式下土壤水中δ18O分布特征,结果表明,浅层土壤水(小于20 cm)由于受到蒸发作用相对富集重同位素(δ18O值较高),而富集轻同位素的较大降水事件(δ18O值较低)可通过活塞流的方式快速与浅层土壤水进行混合。相反,深层土壤水残留时间较长(大于 4.5个月),只有经过持续较长时间的雪融水和大量降水后才得到更新。Gazis等[10]进一步指出即使是在较小的空间尺度下,土壤水同位素组成也会因局部排水状况和土壤性质不同而发生变化。姬王佳等[11]研究了陕北黄土区4种土地利用方式(农地、草地、灌木地和乔木地)下土壤水的氢氧稳定同位素分布特征,结果表明,4个样地土壤水氢氧稳定同位素值在浅层(0~3 m)和深层(>12 m)土壤均无明显差异,而在中层(3~12 m)差异较大。不同样地土壤水可能接受不同强度降水的补给,农地和草地也可被强度小的降水事件补给,而沙柳地和杨树地可能主要接受夏秋季暴雨补给。上述研究表明不同地区土壤水分补给机制及同位素分布特征因降水量、土壤类型、植被种类、地形等多因素差异而不同。因此,在干旱半干旱地区研究典型环境下土壤水分的补给特征对于因地制宜提高水资源利用效率和植被可持续建设具有重要意义。
1 材料和方法
1.1 研究区概况
研究区位于毛乌素沙地东南缘圪丑沟流域(38°48′~38°54′N,109°11′~109°29′E),该流域位于陕西省神木市锦界镇西南约23 km处(图1),海拔为1 250~1 280 m。年均气温约为9.2 ℃,大于10 ℃的积温约为3 150 ℃,年均降水量为420 mm,暴雨相对集中,集中分布在7—9月,占全年降水量的60%~75%。流域内地形起伏较小,土壤类型主要为干旱砂质新成土[12]。流域内GWL较浅,不同林地GWL呈现明显的季节波动(图2)。
关于毛乌素沙区地下水来源目前尚无定论,主要存在两种观点,一种观点认为鄂尔多斯盆地丰富的地下水是由当地降水入渗补给,另一种观点认为地下水的主要补给源是通过断裂带输入的外源水[13],因此关于该流域地下水的来源问题暂不做讨论。近年来,由于大规模植树造林活动的开展,使得该区域植被盖度显著增加,然而,大规模的植树造林破坏了局部原有的生态水文格局,使得土壤水分供需矛盾加剧,流域内不同GWL埋深下的固沙植物部分已出现退化现象。目前,流域内植被类型主要以人工林、草为主,落叶灌木包括沙柳(Salixpsammophila)、紫穗槐(Amorpha fruticoseLinn.)、长柄扁桃(Amygdalus pedunculataPall.),常绿乔木樟子松(Pinus sylvestrisL. var.)和多年生草本沙蒿(Artemisia desterorumSpreng.)等。本研究采用稳定同位素技术,选取圪丑沟小流域的典型治沙植物(沙柳、樟子松和长柄扁桃)为研究对象,系统分析不同GWL埋深下3种林地土壤水同位素分布特征及影响因素。在试验初期对3种林地典型植物样地进行调查,各林地植物生长状况和耗水特征见表1。在试验开始前对各林地剖面土壤物理性质进行调查,调查结果详见表2。在各林地分别安装地下水位仪(HOBO probes,U20-001-04,Onset,Bourne,USA,±0.1 cm)用于自动监测GWL的季节变化特征,同时在流域内安装小型气象站用于监测和获取气象参数。
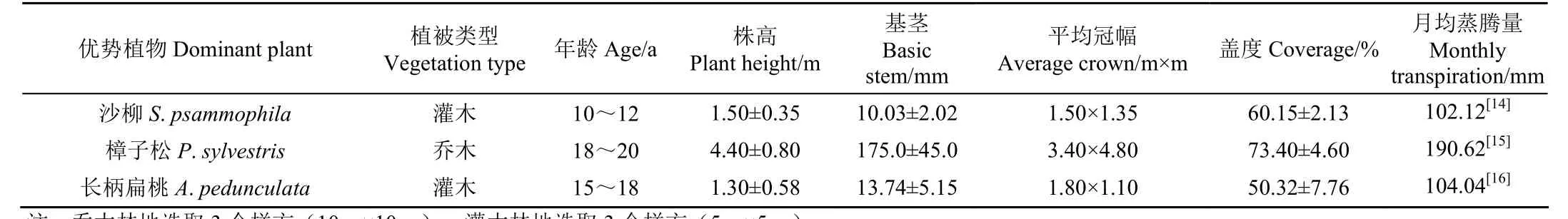
表1 林地植物生长状况Table 1 Vegetation growth status of the woodland
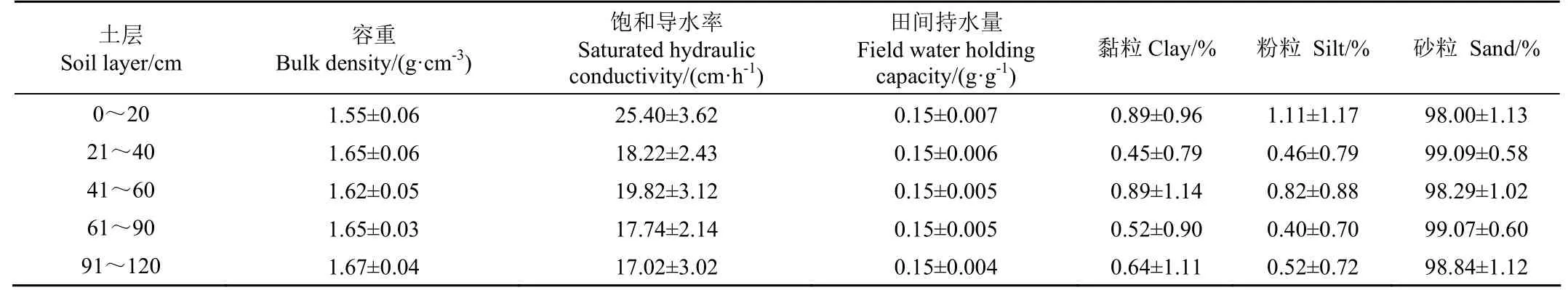
表2 林地土壤物理性质Table 2 Soil physical properties of the woodland
1.2 样品采集
试验期间(6—11月)于每月中旬定期采集沙柳、樟子松和长柄扁桃林地各土层土壤水和地下水样品。采用土钻法对土壤样品进行采集,0~60 cm每间隔20 cm采集一次,60 cm以下至地下水界面每间隔为30 cm采集一次,每个林地各采集3次重复。将采集后的土壤样品分为2份,一份置于棕色玻璃瓶(30 mL)中用于测定氢氧稳定同位素组成,另一份置于铝盒中带回实验室烘干后用于土壤质量含水率的测定。降水样品按降雨事件采集。地下水样品取自各林地水位观测井(井口直径D=10 cm),将采样瓶放入观测井中距水面约40 cm处采集地下水样品,采样前对采样瓶进行润洗,每次采集3次重复。将上述用于同位素测定土壤、降水以及地下水样品用封口膜密封后放于-20 ℃冰箱冷冻保存。
1.3 样品测定
利用低温冷冻抽提系统提取土壤水,采用液态水同位素分析仪(LGR 测量精度:δ18O 为± 0.3‰,δ2H为 ±1‰)对土壤水、地下水和降水氢氧同位素组成(δ2H和δ18O)进行测定。氢氧同位素测定结果根据国际标准物平均海洋水(Vienna Standard Mean Ocean Water,VSMOW)同位素浓度的千分差计算获得
式中R样为待测样品的氢或氧同位素组成,RVSMOW为国际标准物平均海洋水的氢或氧同位素组成。
采样期间降水δ2H和δ18O的加权平均值(δp,mean)计算如下:
式中δi(‰)和PPTi(mm)分别表示每次降水事件的同位素组成和降雨量。
采样期间土壤水δ2H和δ18O的加权平均值(δi,mean)计算如下:
式中δi表示土壤水δ2H 或δ18O(‰),SWCi代表各土层土壤储水量(mm)。
A:可以了解整个印刷行业的发展和变化情况。世界顶尖的印刷企业都会参加这种比赛,通过这个比赛可以了解到其他企业的优点,也可以了解到竞争对手在做什么、发展是怎样的。我们一般都会派技术人员去现场学习。
1.4 数据分析
采用SPSS 18.0 统计软件进行数据统计与分析,采用Origin 2021制图软件进行绘图。
2 结果与分析
2.1 监测期间不同林地降水、土壤水和地下水中δ2H和δ18O统计描述
表3 为沙柳、樟子松和长柄扁桃林地6—11月降水、土壤水和地下水中δ2H和δ18O值统计比较。表3显示各水源δ2H和δ18O加权平均值大小依次为降水>土壤水>地下水,其中降水δ2H和δ18O加权平均值显著高于地下水(P< 0.05),而土壤水δ2H和δ18O加权平均值与地下水相近,这表明土壤水受地下水的补给作用大于降水对土壤水的稀释和混合作用。与土壤水和地下水相比,降水相对富集重组分同位素,这是由于降水在到达地面之前受强烈的蒸发作用而产生同位素非平衡分馏现象。3种林地土壤水δ2H和δ18O值接近,且变异系数(< 25%)均低于降水(表 3)。这是由于降水进入土体后与土壤水中轻组分同位素发生了不同程度的混合,使得混合后的土壤水趋于富集轻组分同位素。地下水δ2H和δ18O变异系数显著低于降水和土壤水(表3),表明研究区地下水氢氧同位素组成相对稳定。在全球降水中由于氢氧同位素组成存在平行分馏效应,因此降水中氢氧同位素组成呈现一定的线性关系,Craig[17]将其定义为大气水线,大气水线的截距和斜率能够在一定程度上反映区域气候条件和降水情况。本研究结果表明研究区当地大气水线为δ2H=7.79δ18O +8.87(图3),其斜率和Y轴截距均略小于全球大气水线(δ2H=8δ18O +10)[17]。研究区氢氧同位素组成表现为重组分相对富集,表明该地区气候相对干燥,蒸发作用较为强烈高于全球平均水平[18]。3种林地(沙柳、樟子松和长柄扁桃)土壤水δ2H和δ18O值大部分分布在大气水线的右侧,其斜率介于5.69~7.13之间,截距介于-19.95~-5.11之间,且均小于当地大气降水线,表明降水在转化为土壤水之前经历了一定程度的蒸发富集现象。这与姬王佳等[11]在陕北黄土区对当地降水和土壤水氢氧同位素值分析的结果一致。邢丹等[19]同样研究表明同时期荒漠地区土壤水同位素较降水富集重组分。相对于樟子松和长柄扁桃林地,沙柳林地的土壤水线斜率更接近与当地降水线,且显著高于其它两个林地。这可能与不同植被冠层截留和局部微环境的差异有关[20]。

表3 研究区各林地监测期间不同水源δ2H和δ18OTable 3 δ2H 和δ18O values of different water sources in various woodland during the monitored periods in the study area ‰
2.2 不同林地土壤水δ18O剖面分布特征
检测期间沙柳、樟子松和长柄扁桃林地土壤水δ18O垂直分布特征如图4所示。由于沙柳、樟子松和长柄扁桃林地土壤水δ2H和δ18O存在较好的线性相关关系(图 3),因此本文以氧同位素为例阐述检测期间土壤水δ18O剖面分布动态变化特征。3种林地土壤水δ18O值剖面分布随着深度的加深表现出不同的季节变化特征(图 4)。研究区旱季(6月、10月和11月),不同林地土壤水δ18O值随着深度的加深均呈逐渐减小的趋势,表层土壤水重组分同位最为富集(图4)。在这一期间降水分布相对较少,土壤水受蒸发作用影响大于降水对土壤水的补给作用,重组分同位素相对富集。这与该时期土壤含水率的剖面变化特征(图5)相吻合。随着雨季(7—9月)降水量的增加,这一情况得到显著改善,3种林地(沙柳、樟子松和长柄扁桃)雨季(7—9 月)土壤剖面δ18O平均值分别低于旱季(6月,10月和11月)0.81‰~2.67‰、0.50‰~3.32‰和1.26‰~2.95‰(图5)。这一时期降水在转化为土壤水过程中对土壤中“老水”稀释混合作用频繁,使得土壤水表现为轻组分同位素相对富集而重组分同位素相对贫化。
为进一步量化降水和地下水对土壤水的补给深度将不同土层土壤水δ18O与降水或地下水δ18O(月加权平均值)进行了相关分析(图6)。从图6可以看出,沙柳林地0~20 cm土层土壤水δ18O与降水δ18O显著相关(P<0.05),而樟子松和长柄扁桃林地0~20 cm土层和20~40 cm土层土壤水δ18O与降水δ18O均显著相关(P< 0.05),而其它土层土壤水δ18O与降水δ18O未发现相关关系(P>0.05)。因此,可以认为降水对沙柳林地土壤水的平均补给深度主要集中在0~20 cm土层,对樟子松和长柄扁桃林地土壤水的平均补给深度主要集中在0~40 cm土层。相对于樟子松和长柄扁桃,沙柳林地降水补给土壤水的深度较浅,这可能是由于沙柳冠幅较大且密度较高,冠层截流较强导致雨水对土壤水的补给减少,此外,沙柳林地表层广泛分布苔藓层也可能是导致该林地降水补给较浅的一个重要影响因素。除降水对土壤水补给外,地下水也是该地区土壤水的另一重要补给来源。如图5所示沙柳林地>180 cm土壤水、樟子松林地>60 cm土壤水和长柄扁桃林地>120 cm土壤水δ18O平均值分别为-10.22‰,-8.53‰和-10.83‰,均与地下水δ18O平均值(-10.38‰,-8.69‰和-10.99‰)接近。相关性分析表明沙柳林地180~210 cm土层和210~240 cm土层土壤水δ18O均与地下水δ18O显著相关(P< 0.05),而樟子松林地60~90 cm土层、长柄扁桃林地120~150 cm土层土壤水δ18O与地下水δ18O具有显著的相关关系(图6,P< 0.05)。因此,可以认为沙柳林地180 cm以下土壤受到地下水补给影响显著,而樟子松林地60 cm以下土壤和长柄扁桃林地120 cm以下土壤受到地下水补给影响显著。此外,由于不同季节GWL处于波动状态,因此根据监测期间GWL动态变化特征,同时结合地下水的补给深度,可以计算出地下水向上补给土壤水的深度和范围。计算表明3种林地(沙柳、樟子松和长柄扁桃)GWL变化范围分别为253~260、87~93和172~176 cm,因此不同林地地下水向上补给土壤水的深度和范围分别约为73~80、27~33和52~56 cm。不同林地地下水补给土壤水的深度和范围不同,一方面可能与林地所处的沙丘部位和GWL不同有关,另一方面可能与不同植被根系生物量在不同土层中的含量(图7)及耗水量不同有关,未来仍需进一步研究。
3 讨 论
3.1 降水的入渗补给特征
在干旱半干旱地区降水是土壤水的重要补给来源。本研究结果显示3种林地(沙柳、樟子松和长柄扁桃)土壤水δ2H和δ18O值大部分位于当地大气水线的右侧,且不同林地土壤水线斜率(5.69~7.13)均小于当地大气水线斜率(7.79)(图3),表明降水在转化为土壤水的过程中存在一定程度的蒸发分馏现象。这与前人[11,21]在该地区研究的结论一致。雨季(7—9月),降水相对较为频繁,各林地浅层土壤水得到快速补充,平均土壤含水率(< 40 cm)显著高于旱季(6月,10月和11月)(图 4)。当降水事件发生后,各林地表层土壤含水率(<40 cm)迅速达到0.20 g/g,高于田间持水量(约为0.15 g/g),表明降水在向下入渗过程中主要以重力水的形式向下运动并与土壤中原有“老水”发生混合和稀释致使土壤水氢氧同位素组成表现为轻组分富集[22]。Hsieh等[23]认为降水补给开始于土壤表层,并逐渐向下补给,0~40 cm土壤水δ18O的变异性大于40 cm以下土壤。本文研究结果与Hsieh等[23]的研究结果一致。王锐等[24]研究发现黄土高原长武塬区0~10 cm土壤水直接受大气降水影响,雨水的稀释作用导致0~10 cm土壤水δ18O降低。本研究降水与各林地0~40 cm土壤水氧同位素相关性分析表明降水与各林地浅层土壤水氧同位素存在显著的相关关系。这表明浅层土壤水更易受到降水补给的影响。这与张军红等[25]对毛乌素沙地西缘流动沙丘浅层土壤水的动态变化研究结论一致。Dansgaard[26]在热带及中纬度地区的研究结果表明,受降水的补给浅层土壤水表现出重组分同位素贫化轻组分同位素富集的现象,降水量与土壤水同位素值呈负相关关系。随着土壤水的进一步入渗,土壤水由以重力水向下入渗的方式转变为以薄膜水扩散方式从高含水率土壤向低含水率土壤扩散运动,此时进一步与“老水”混合稀释,氢氧同位素组成亦表现为重组分相对富集。雨季土壤含水率剖面分布特征进一步印证了这一结论。此外,本研究根系采样结果显示(图7),3种植物(沙柳、樟子松和长柄扁桃)土壤表层(< 40 cm)根系生物量占总根系生物量的75.23~80.46%,这有利于形成降水入渗通道,且研究区土壤质地以砂土为主,因此,当降雨强度较大时该地区土壤水分运动可能主要以优先流[27]的形式向下入渗。Konz等[28]发现瑞士中部山地不同土地利用方式下10~35 cm土壤含水率对降水响应较深层土壤水更快,并强调表层植物根系活动和疏松质地是主要影响因素,Gazis 等[10]同样指出降水入渗土壤的方式受土壤质地、孔隙、植物类型以及降雨强度等多种影响呈现出不同的运动方式。
3.2 地下水位动态变化对土壤水的补给影响
在本研究区域,土壤水的补给除降水外,浅层地下水也是土壤水重要的补给来源。过去的研究表明荒漠地区浅层地下水是土壤水重要补给来源同时也是荒漠植物赖以生存的基础[29-30]。GWL的时空变化直接影响深层土壤水的补给与分布特征,从而影响植被的生长与演替[31]。地下水与其临近的土壤通常处于不断的水分交换与循环过程。刘君等[32]在呼和浩特西南部研究表明深层土壤水与浅层地下水的水分交换十分活跃,使得深层土壤水氢氧同位素组成与地下水接近。本研究结果表明3种林地地下水与相邻土层土壤水δ2H和δ18O显著相关(图6),且地下水δ2H和δ18O组成相对稳定。冯蕴等[33]在浑善达克沙地南缘研究结果同样表明与地下水相邻土层中土壤水氢氧同位素组成处于相对平稳状态,δ2H和δ18O同样具有弱的变异特征。这进一步表明浅层地下水对深层土壤水氢氧同位素分布具有显著影响。因此,浅层地下水作为重要水分来源对研究地区土壤水的补给具有导向作用。
浅层地下水对土壤水的补给主要通过毛管作用力来补给相邻土层[34-35]。然而本研究区土壤类型为砂土,毛管孔隙大,毛管重力水上升作用有限,对浅层土壤的补给可能主要以膜扩散的方式由高含水量向低含水率方向扩散运动。而氢氧同位素值在剖面上表现为逐步接近地下水氢氧同位素值[36]。本研究结果显示,在旱季(6月,10月和11月)随着土壤深度的增加,土壤水氢氧同位素重组分逐渐贫化。地下水在向上补给土壤水过程中,氢氧同位素发生分馏作用,其轻组分逐步贫化。这进一步印证了这一结论。依据不同土层土壤水氢氧同位素与地下水同位素组成结合土壤含水率的剖面分布特征可以得到土壤水的补给深度和范围。本研究结果显示3种林地(沙柳、樟子松和长柄扁桃)地下水向上补给土壤水的深度和范围分别为73~80、27~33和52~56 cm。不同林地地下水补给土壤水的深度和范围不同,一方面可能与林地所处的沙丘部位与GWL不同有关,另一方面可能与植物根系剖面分布及耗水差异有关。这需要进一步研究。
3.3 展 望
降水、土壤水和地下水是陆地生态系统水资源的主要存在形式。毛乌素沙地蒸发强烈,在降水季节分布不均的气候条件下,土壤水成为当地植物生存发展最为重要的限制因子。本文采用氢氧稳定同位素分析技术,通过定期测定降水、土壤水和地下水的氢氧同位素组成,分析了不同GWL埋深下土壤水δ2H和δ18O剖面分布动态变化特征及其影响因素,同时结合土壤含水率探讨了土壤水的补给来源和运移规律。本研究结果显示:研究区降水主要补给浅层土壤水(沙柳林地0~20 cm、樟子松和长柄扁桃林地0~40 cm),而不同埋深地下水主要补给深层土壤水(沙柳林地180 cm以下,樟子松林地60 cm以下,长柄扁桃林地120 cm以下)。由于毛乌素沙地气候干旱、降水季节分布不均,浅层地下水对深层土壤水的补给可为人工固沙植物生长提供重要的水分来源。然而,随着全球气候暖干化[37],区域植被生长面临的干旱胁迫将日益加剧[38-39]。特别是毛乌素沙地近年来大规模开展植树造林,可能进一步加剧植被耗水与土壤供水之间的矛盾,从而威胁区域生态安全和植被可持续建设。因此,未来需查明毛乌素沙地地下水资源储量以及来源,并明确不同固沙植物吸水来源与耗水量,从而做到“以水定林,量水而行”,促进毛乌素沙地人工生态系统健康与可持续发展。
4 结 论
本研究通过定期测定毛乌素沙地圪丑沟小流域3种林地(沙柳、樟子松和长柄扁桃)降水、土壤水和地下水氢氧同位素组成,研究了不同地下水位埋深下土壤水δ2H和δ18O剖面分布动态变化特征及其影响因素,并定量分析了降水和地下水对不同林地土壤水的补给深度和范围。监测期间(6—11月)沙柳林地0~20 cm土壤水、樟子松和长柄扁桃林地0~40 cm土壤水均表现出重组分同位素贫化轻组分同位素富集的现象,且与降水同位素呈显著相关关系(P < 0.05),表明不同林地浅层土壤水(< 40 cm)更易受到降水补给的影响。3种林地深层土壤水(沙柳林地180 cm以下,樟子松林地60 cm以下,长柄扁桃林地120 cm以下)与地下水δ2H和δ18O的均值接近,且土层深度接近地下水时土壤水δ2H和δ18O逐渐贫化,并趋于稳定。不同埋深地下水(沙柳林地地下水位范围253~260 cm、樟子松林地地下水位范围87~93 cm、长柄扁桃林地地下水位范围172~176 cm)向上直接补给土壤水的深度范围分别为73~80、27~33和52~56 cm。毛乌素沙地浅层地下水对深层土壤水的补给可在一定程度上缓解旱季土壤水分亏缺,为保障该区人工固沙植被生长提供潜在的水分来源。
