以桉树皮为原料的有机基质育苗效果
2021-09-14黎书会吴疆翀贺思腾孙恒郑益兴张燕平张体坤刘剑金童清唐红燕孙启涛
黎书会 吴疆翀 贺思腾 孙恒 郑益兴 张燕平 张体坤 刘剑金 童清 唐红燕 孙启涛

摘 要:為了以环保的方式提高桉树皮的利用价值,利用微生物腐熟发酵,使其成为理化性质与泥炭土相似的有机基质。通过培育经济植物辣木、青花、生菜、番茄、烟草和生石花,探讨桉树皮原料基质的育苗效果。研究不同腐熟程度桉树皮的主要养分含量,以及复配不同比例珍珠岩(10%、20%、30%)对6种供试植物种子发芽率,苗期株高、冠幅和地径的影响。结果表明:腐熟60 d的桉树皮内速效氮磷钾的含量都高于泥炭土,且6种供试植物的生长量都显著高于生料(未经腐熟)与腐熟40 d的桉树皮(P<0.05)。其中腐熟60 d的桉树皮作为基质培育青花、番茄与生菜的效果均优于对照泥炭土,并以复配30%珍珠岩的效果为佳。而生石花的发芽率,以及辣木和烟草的生长量显著低于泥炭土(P<0.05),推测其受桉树内源生长抑制物的影响。研究结果表明腐熟60 d的桉树皮可作为基质代替泥炭土适用于青花、生菜和番茄育苗。而桉树皮腐熟程度与抑制物含量的关系,以及在辣木、烟草与生石花等抑制物耐受性较低植物上的应用,还有待进一步研究。
关键词:桉树皮;经济植物;抑制物;育苗效果
Abstract: In order to improve the utilization value of Eucalyptus bark in an environmentally friendly way, microorganisms are used to decompose it and make it become an organic substrate with physical and chemical properties similar to peat moss. Through cultivating the economic plants Moringa oleifera, broccoli, lettuce, tomato, tobacco and lithops, the seedling effect of Eucalyptus bark as the raw material for substrate was discussed.. The main nutrient contents of Eucalyptus bark with different maturity and the effects of Eucalyptus bark substrate with different proportions of perlite (10%, 20% and 30%) on seed germination rate, seedling height, crown diameter and ground diameter of six tested plants were studied. The content of available N, P and K in Eucalyptus bark decomposed for 60 days was higher than that in peat moss. The growth of six tested plants at seedling stage in Eucalyptus bark decomposed for 60 days were significantly higher than that in raw and 40 days decomposed Eucalyptus bark (P<0.05). Moreover, the effect of Eucalyptus bark as the substrate decomposed for 60 days used to cultivate broccoli, tomato and lettuce was better than that of the control peat moss, and Eucalyptus bark decomposed for 60 days with 30% perlite was the best. However, the germination rate of lithops and the growth of M. oleifera and tobacco were significantly lower than those in peat moss (P<0.05), which was speculated to be influenced by the endogenous growth inhibitors of Eucalyptus. It is suggested that Eucalyptus bark which was decomposed for 60 days could be used as the substrate instead of peat moss, which was suitable for growing seedlings of broccoli, lettuce and tomato. Nevertheless, the relationship between the maturity of Eucalyptus bark and the inhibitor content, and the application of Eucalyptus bark to plants with low tolerance to inhibitors such as M. oleifera, tobacco and lithops requires further study.
Keywords: Eucalyptus bark; economic plants; inhibitors; nursery effect
近年来,轻型有机育苗基质促进了农林业育苗由自然种植方式向工业化生产方式转化,显著增进了植株的商品化。传统育苗基质泥炭土因其质轻并含有丰富的腐殖质,育苗效果佳,因而市场需求一度旺盛。但其天然不可再生,属于非环保型基质,大量开采泥炭土是对地球之肾——湿地环境的破坏[1-3]。而林产加工剩余物价格低廉、材料广泛、理化性质稳定,可作为规模化加工有机育苗基质的原料。桉树速生成材、轮伐期短,已成为我国南方地区重要的速生用材树种[4-6]。但随着大面积營造桉树人工林,桉树内源抑制物随树体枯落物释放至土壤,对林下许多植物的种子萌发和幼苗生长产生了抑制作用[7-11]。由于较难以环保方式加以利用,每年加工桉树木材产生的大量树皮木屑多被烧掉,造成大气污染;或自然堆弃分解形成仅剩无机质的灰分,失去利用价值,其生态问题日益得到重视[12]。而以桉树皮作为基质原料时,内源抑制物如果分解不完全会抑制种子萌发和幼苗生长[13-14]。据查,现国内外尚无完全攻克该技术要点的报道。发酵桉树皮仅用作少量添加的辅料或只适合某些菌菇生长[15-17]。本研究旨在通过实际育苗效果,检验桉树皮作为基质主料进行经济植物育苗的可行性,并为进一步研究如何生物降解桉树原料内源抑制物以及评估桉树皮基质应用前景提供参考依据。
1 材料与方法
1.1 材料
1.1.1 实验时间、地点 育苗实验与测定实验于2019年9—11月在中国林业科学研究院资源昆虫研究所内进行,地理位置为东经10276,北纬256,海拔1919.2 m。育苗大棚内日最高温41 ℃,最低温9 ℃,平均温25.8 ℃,平均相对湿度64%,平均日光照时长11.4 h。
1.1.2 实验材料 尾巨桉(Eucalyptus urophylla ×E. grandis)树皮粉碎后纤维长2~8 mm的生料,分别腐熟40 d和60 d;粒径为3~6 mm的珍珠岩;市售市场占有率高且育苗效果口碑好的泥炭土(丹麦品氏Pindstrup,购自昆明斗南花卉农资市场);供试植物6种,分别为蔬菜青花(碧玉西兰花)、生菜(奶油生菜)和番茄(粉冠2号),多用途经济树种辣木(中林3号辣木良种)、生石花(南非混种)以及烟草(品种K326)。
1.2 方法
1.2.1 实验设计 本实验以3种不同腐熟程度桉树皮复配珍珠岩(体积占比10%、20%与30%),共设10个处理。1、2、3号处理为分别加入10%、20%、30%珍珠岩的桉树皮生料;4、5、6号处理分别为加入同上配比珍珠岩的腐熟40 d桉树皮;7、8、9号处理分别为加入同上配比珍珠岩的腐熟60 d桉树皮;0号处理为对照(泥炭土)。育苗方式采用12孔漂浮育苗盒育苗。播种后发芽率超过50%时,每周对各处理植物幼苗(辣木随机选取30株,其他植物随机选取50株)的发芽数、株高、冠幅与地径进行调查测定,于第10~11周测量每个处理30株植株的鲜重及干重。
1.2.2 桉树皮的微生物腐熟发酵 桉树皮原料的腐熟发酵方法参见本研究组前期国家发明专利“以思茅松树皮为原料的有机育苗轻基质的制备方法”(ZL 201210498290.8)。
1.2.3 主要养分含量测定 参照国家农业行业标准NY和林业行业标准LY测定桉树皮生料、腐熟40 d桉树皮、腐熟60 d桉树皮的主要养分含量。pH测定参照土壤pH的测定(NY/T1377—2007)方法;有机质测定参照土壤有机质的测定(NY/T1121.6— 2006)方法;速效氮的测定参照森林土壤氮的测定(LY/T1228—2015)方法;速效磷的测定参照土壤有效磷的测定(NY/T1121.7—2014)方法;速效钾的测定参照土壤速效钾和缓效钾含量的测定(NY/T889—2004)方法;腐殖质的测定参照土壤腐殖质组成的测定(NY/T1867—2010)方法。
1.3 数据处理
数据统计采用Microsoft Excel 2010软件;方差分析和差异显著性检验(Duncan新复极差法)采用SPSS 17.0软件;作图采用Sigma Plot 12.5软件。
2 结果与分析
2.1 不同腐熟程度桉树皮内主要养分含量
3种不同腐熟程度桉树皮与泥炭土内主要养分含量见表1。从表1可见,随着腐熟程度的加深,各种养分都被逐步有效释放,值得注意的是,从生料到腐熟40 d,然后继续腐熟至60 d,树皮原料内部主要养分的释放经历了先慢后快的过程。对比腐熟60 d桉树皮与泥炭土的养分含量,可以发现,除了有机质外,其速效氮磷钾含量都较泥炭土高。对比pH与腐殖酸的含量,未经腐熟的生料pH呈弱酸性,加入尿素等腐熟材料后,pH升高。经过腐熟40 d后,其中有机质经过各种微生物的作用,形成腐殖质,因此腐殖酸含量升高,直至进一步的腐熟,腐殖酸被大量有效释放,pH降低,呈中性。泥炭土内腐殖酸含量高于桉树皮,pH低于桉树皮,呈酸性。
2.2 不同腐熟程度桉树皮作为基质对植物生长的影响
2.2.1 不同腐熟程度桉树皮作为基质对植物发芽率的影响 每种植物的发芽率对10个处理的响应如图1。截止播种4周后青花发芽率为50.00%~74.44%,生料与腐熟60 d桉树皮显著高于泥炭土(P<0.05);播种4周后生菜发芽率为78.33%~92.22%,8号处理(80%腐熟60 d+20%珍珠岩)发芽率与泥炭土(90.00%)无显著差异(P>0.05);播种5周后番茄发芽率为32.22%~ 85.00%,9号处理(70%腐熟60 d+30%珍珠岩)发芽率与泥炭土(90.00%)无显著差异(P>0.05);播种6周后辣木发芽率为51.67%~74.17%,2号处理(80%生料+20%珍珠岩)和9号处理都显著高于泥炭土(P<0.05);烟草播种6周后发芽率和生石花播种5周后发芽率在不同腐熟程度桉树皮中都显著低于泥炭土(P<0.05)。
2.2.2 不同腐熟程度桉树皮作为基质对植物生物量的影响 不同处理基质对每种植物整株鲜重的影响具有显著差异,经过LSD处理后检验可以看出(图2):3种蔬菜(青花、生菜、番茄)在腐熟60 d桉树皮中的鲜重显著高于生料和腐熟40 d桉树皮(P<0.05),而生菜和番茄在生料与腐熟40 d桉树皮中的鲜重无显著差异(P>0.05)。青花在6号处理(70%腐熟40 d+30%珍珠岩)中的鲜重显著高于腐熟40 d桉树皮复配10%与20%珍珠岩的4号和5号处理(P<0.05),且显著优于生料(P<0.05)。青花在9号处理(70%腐熟60 d+30%珍珠岩)中的鲜重和在泥炭土中的鲜重差异不显著(P>0.05)。生菜在8号处理(80%腐熟60 d+20%珍珠岩)中的鲜重、番茄在7号(90%腐熟60 d+10%珍珠岩)和9号处理(70%腐熟60 d+30%珍珠岩)中的鲜重都显著高于泥炭土(P<0.05)。此外,辣木的整株鲜重对不同处理的响应同上述3种蔬菜不同,主要表现在辣木在腐熟60 d桉树皮中的鲜重显著低于泥炭土(P<0.05),并且在生料中的鲜重显著优于腐熟40 d和60 d的桉树皮(P<0.05)。烟草在腐熟60 d桉树皮中鲜重与其在泥炭土中的鲜重均无显著差异(P>0.05),但显著优于生料(P<0.05)。生石花在腐熟60 d桉树皮中的鲜重和干重与其在泥炭土中的鲜重和干重均无显著差异(P>0.05),但显著高于腐熟40 d桉树皮和生料。
分析不同处理对以上各种植物整株干重的影响,发现与其对鲜重影响的不同在于9号处理(70%腐熟60 d+30%珍珠岩)的青花整株经过烘干后的干重与泥炭土差异显著(P<0.05),而7号(90%腐熟60 d+10%珍珠岩)与8号处理(80%腐熟60 d+20%珍珠岩)的生菜干重、7号(90%腐熟60 d+10%珍珠岩)与9号处理(70%腐熟60 d+30%珍珠岩)的番茄干重与泥炭土差异不显著(P>0.05)。2号处理(80%生料+20%珍珠岩)中的辣木干重与泥炭土差异不显著(P>0.05)。烟草在腐熟60 d桉树皮中干重显著低于其在泥炭土中的干重(P<0.05),但显著高于生料与腐熟40 d桉树皮(P<0.05)(图3)。
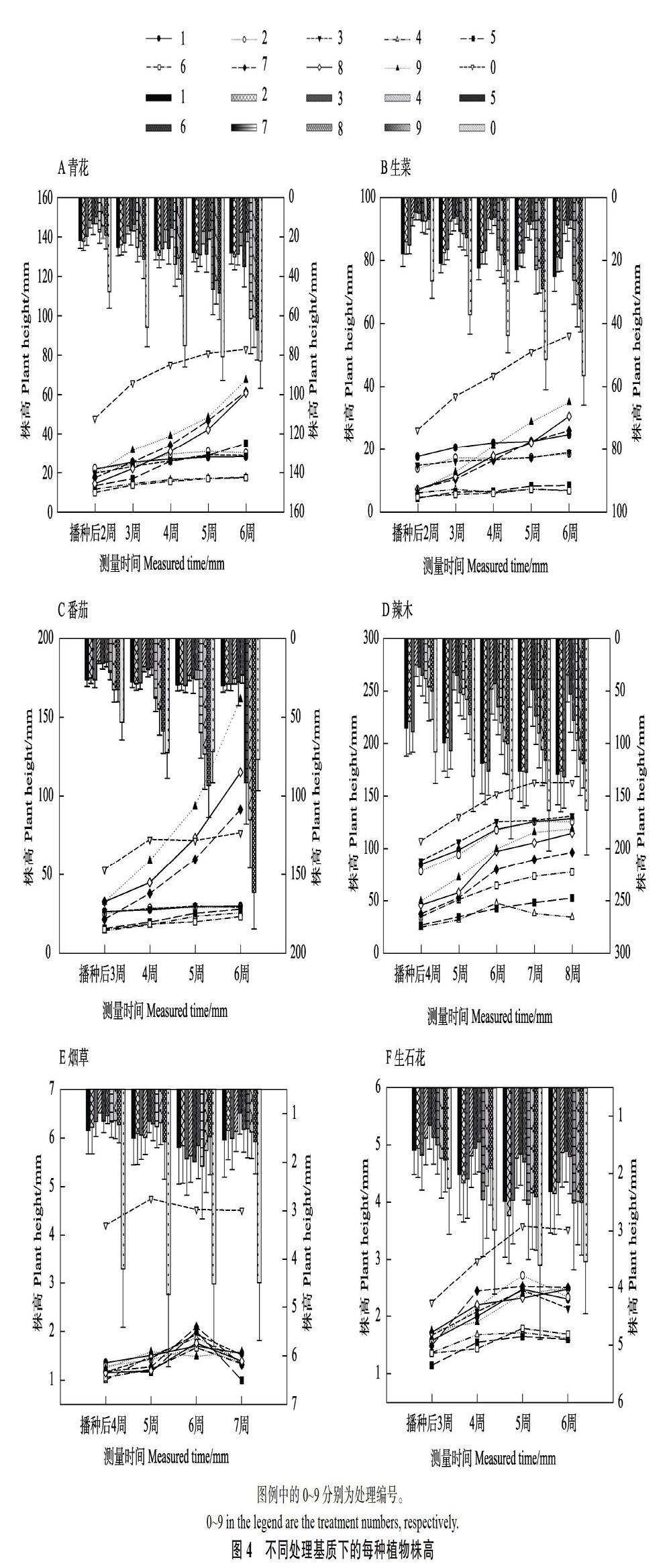
2.2.3 不同腐熟程度桉树皮作为基质对植物株高的影响 经对以上每种植物的观察和测量发现,各种植物在泥炭土中普遍优先发芽,与9号处理基质(70%腐熟60 d+30%珍珠岩)相比,蔬菜早1 d,辣木与烟草早3~4 d,生石花同天发芽。而青花与生菜于泥炭土中生长大约1周后出现徒长现象,番茄在泥炭土中生长5周后缺肥致叶片发黄。
经过一般线性模型的重复测量分析发现,每种植物每周的株高生长都有显著差异(P<0.05),且不同处理基质对每种植物的株高生长在不同测量时间上的影响差异显著(P<0.05)。青花在腐熟60 d桉树皮中的株高生长显著优于腐熟40 d与生料(P<0.05),且加入30%珍珠巖的9号处理显著优于加入10%和20%珍珠岩的7号与8号处理(P<0.05),4号(90%腐熟40 d+10%珍珠岩)与6号处理(70%腐熟40 d+30%珍珠岩)对青花株高的影响无显著差异(P>0.05)(图4A)。生菜在不同腐熟程度桉树皮中的株高生长差异显著(P<0.05),在腐熟60 d桉树皮中,生菜株高生长呈线性增高趋势,而在腐熟40 d与生料中的株高生长平缓,9号处理(70%腐熟60 d+30%珍珠岩)与1号处理(90%生料+10%珍珠岩)无显著差异(P>0.05),而除对照泥炭土外,9号与1号处理显著优于其他处理(P<0.05)(图4B)。10种处理对番茄株高的影响表明,泥炭土中因养分不足致番茄株高增长趋于停滞,而番茄在腐熟60 d桉树皮中的株高生长非常良好,其中又以9号处理表现最为显著(P<0.05)(图4C)。辣木在不同腐熟程度桉树皮中的株高生长显著不及泥炭土(P<0.05),而在生料中的株高生长显著优于腐熟40 d和60 d桉树皮(P<0.05)(图4D)。烟草在各处理中的株高生长显著低于泥炭土(P<0.05),且桉树皮各处理间无显著差异(P>0.05)(图4E)。生石花在桉树皮中的株高生长显著低于泥炭土(P<0.05),除腐熟40 d桉树皮外,其他处理间无显著差异(P>0.05)(图4F)。
2.2.4 不同腐熟程度桉树皮作为基质对植物冠幅的影响 不同处理对每种植物苗期冠幅的影响相比株高更为明显(图5)。青花与生菜在泥炭土中由于茎的徒长导致两种蔬菜在苗期冠幅生长不足,而3个腐熟60 d处理中两种蔬菜冠幅的生长显著更优(P<0.05)。在不同处理对青花冠幅的影响中(图5A),腐熟60 d桉树皮对青花冠幅的生长显著优于对照泥炭土和其他处理(P<0.05),其中复配30%珍珠岩的9号处理显著优于复配10%与20%珍珠岩的7号和8号处理(P<0.05),且5号处理(80%腐熟40 d+20%珍珠岩)也明显优于其他复配10%与30%珍珠岩的4号与6号腐熟40 d桉树皮和生料(P<0.05),而加入不同比例珍珠岩于生料中对青花冠幅的生长并无显著差异(P>0.05)。生菜苗在后期叶片生长繁茂,彼时冠幅伸展受限,因此生菜冠幅大小在3个腐熟60 d处理中略有减小,播种后5周和6周冠幅生长无显著差异(P>0.05),但腐熟60 d仍显著优于腐熟40 d和生料(P<0.05)(图5B)。番茄在8号(80%腐熟60 d+20%珍珠岩)与9号处理(70%腐熟60 d+30%珍珠岩)中冠幅生长显著优于泥炭土(P<0.05),且由于番茄苗较生菜苗株型伸展,冠幅生长空间更大,因此冠幅大小并没有受限,此外对于腐熟60 d桉树皮而言,加入不同配比珍珠岩其冠幅生长具有明显差异,其中以加入30%珍珠岩的9号处理效果最佳(P<0.05)(图5C)。分析结果显示,辣木冠幅增长随时间(每周间)具有显著差异(P<0.05),但在同时段各处理间无显著差异(P>0.05)(图5D)。不同处理对烟草冠幅的影响表明(图5E),生料和腐熟40 d桉树皮对烟草冠幅生长的影响没有明显差异(P>0.05),而9号(70%腐熟60 d+30%珍珠岩)与8号处理(80%腐熟60 d+20%珍珠岩)显著优于7号处理(90%腐熟60 d+10%珍珠岩)以及其他处理(P<0.05),9号与8号处理间无显著差异(P>0.05),泥炭土显著优于所有桉树皮基质(P<0.05)。生石花的冠幅在腐熟60 d桉树皮中显著低于泥炭土(P<0.05),而复配30%珍珠岩的9号处理显著优于复配10%与20%珍珠岩的7号和8号处理以及其他处理(P<0.05),此外,加入不同比例珍珠岩于腐熟40 d桉树皮中对生石花冠幅的生长并无显著差异(P>0.05)(图5F)。
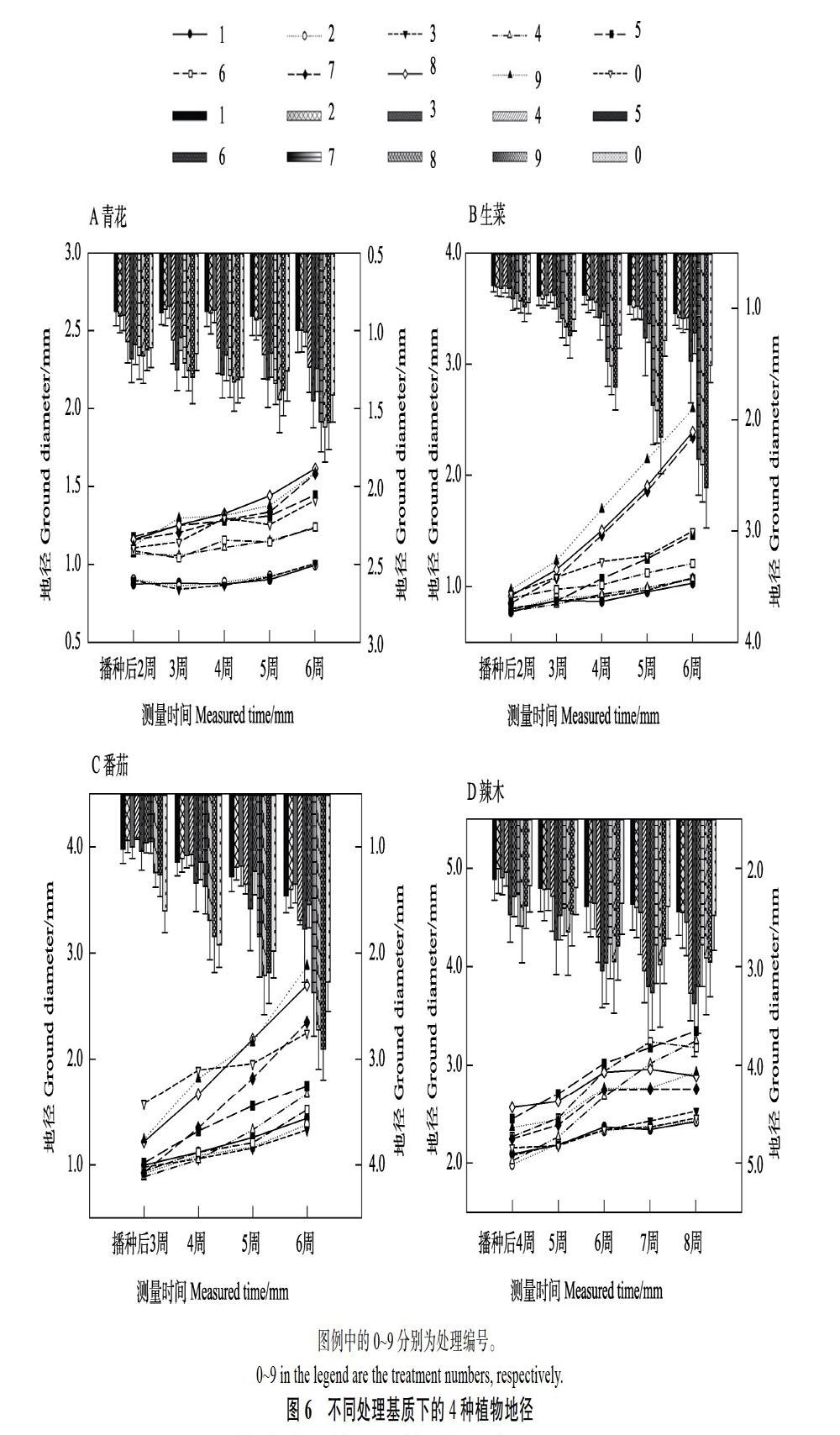
2.2.5 不同腐熟程度桉树皮作为基质对植物地径的影响 不同处理对青花、生菜、番茄、辣木和烟草地径的影响与其对冠幅的影响基本一致(图6),不同点主要表现在:青花在5号处理(80%腐熟40 d+20%珍珠岩)中地径生长显著优于泥炭土(P<0.05),且与7号处理(90%腐熟60 d+10%珍珠岩)无显著差异(P>0.05),但显著低于8号(80%腐熟60 d+20%珍珠岩)与9号处理(70%腐熟60 d+30%珍珠岩)(P<0.05)(图6A)。生菜在3个腐熟60 d桉树皮处理中的地径生长都显著优于泥炭土(P<0.05),在腐熟40 d桉树皮中的地径生长表现也明显优于生料(P<0.05),且每周地径生长差异显著(P<0.05)(图6B)。番茄在8号处理(80%腐熟60 d+20%珍珠岩)中地径生长表现与泥炭土没有显著差异(P>0.05)(图6C)。不同处理对辣木地径生长在不同测量时间上的影响差异显著(P<0.05),但辣木在5号(80%腐熟40 d+20%珍珠岩)、6号(70%腐熟40 d+30%珍珠岩)和8号处理(80%腐熟60 d+20%珍珠岩)中地径生长无显著差异(P>0.05),但显著优于其他处理(P<0.05)。4号(90%腐熟40 d+10%珍珠岩)、7号(90%腐熟60 d+10%珍珠岩)和9号处理(70%腐熟60 d+30%珍珠岩)对辣木地径生长也无显著差异(P>0.05),但显著优于泥炭土和生料(P<0.05)(图6D)。不同处理对烟草地径生长的影响规律与其对冠幅的影响相同。
2.2.6 不同腐熟程度桉树皮育苗效果综合评价 綜合不同腐熟程度桉树皮的育苗效果,青花在9号处理基质(70%腐熟60 d+30%珍珠岩)中的发芽率高于泥炭土,且冠幅与地径生长显著优于泥炭土(P<0.05),而鲜重与泥炭土差异不显著(P>0.05),因此青花育苗使用9号处理基质比泥炭土更优。生菜在8号处理基质(80%腐熟60 d +20%珍珠岩)中发芽率比泥炭土高,且鲜重显著优于对照泥炭土(P<0.05),干重与对照泥炭土差异不显著(P>0.05),而其冠幅与地径在9号处理基质中都显著优于泥炭土(P<0.05),因此生菜育苗使用8号与9号处理基质效果更佳。番茄除在9号处理基质中发芽率不及泥炭土外,其他各项指标都显示9号处理基质对其的育苗效果比泥炭土更优。辣木在泥炭土内的生物量显著高于其他处理基质(P<0.05),且其株高生长也显著优于其他处理(P<0.05),因此辣木使用泥炭土育苗效果优于桉树皮基质。生石花在桉树皮基质中的发芽率明显不及泥炭土,且其在泥炭土内的生长也显著优于桉树皮基质(P<0.05),因此使用泥炭土对生石花进行育苗效果更好。烟草的各项指标都显示泥炭土对其的育苗效果明显优于桉树皮基质。
3 讨论
以桉树皮为原料,其中含有大量木质素、纤维素以及内源生根抑制物质[18-19],经过堆肥技术以及特殊微生物的有效转化在释放养分的同时也促进了抑制物的分解,其中将桉树皮腐熟至60 d左右时,养分得到充分释放并不至于流失,理化性质与泥炭土相似,且有机质和养分含量不低于泥炭土,可作为具有耐受桉树内源抑制物的经济林木、经济作物育苗的有机基质,从而替代传统不可再生资源泥炭土,达到促进生态与经济双重效益的目的。
经过育苗实验研究发现:(1)供试植物种子在生料中发芽率普遍较高,推测这与生料具有较大孔隙度,未经腐熟作用,其内源抑制物较少释放有关。但除辣木外,幼苗在生料中生长养分明显不足。辣木种子由于具有发达胚乳[20],因此自身提供了较充足养分,在生料中苗期生长没有受到明显影响。(2)3种蔬菜在腐熟60 d桉树皮中的鲜重显著高于泥炭土(P<0.05),但经过对其植株烘干处理后,植株干重差异不显著(P>0.05),且明显低于泥炭土(P<0.05),说明生长于腐熟60 d桉树皮处理中的植株含水率普遍比其在泥炭土中高。此外,供试植物在泥炭土中的株高和冠幅生长在后期减缓,而在腐熟60 d桉树皮中的生长普遍表现良好,且并没有出现徒长与缺养分现象。说明泥炭土的保水性能不及腐熟60 d桉树皮,肥效较之腐熟60 d桉树皮持续性不足。(3)对青花与辣木而言,腐熟40 d桉树皮中的高抑制物含量抑制了其株高与冠幅的生长,营养运输本着就近原则,使其地径增长更明显,尤其在属于木本植物的辣木中尤为明显。(4)腐熟40 d桉树皮对绝大多数试验植物的发芽以及苗期生长都具有严重的抑制作用,表现在种子以及发芽幼苗根部发生腐烂,以及幼苗根际变态发育,推测与抑制物大量释放但未分解有关,这与其他人的研究结果一致[21-22]。(5)与泥炭土相比,腐熟60 d桉树皮对番茄、青花、生菜3种蔬菜以及辣木种子的发芽并没有抑制作用,相反对辣木以及青花种子发芽具有明显的促进作用,这与李思远等[14]的研究结果相似。而腐熟60 d桉树皮内抑制物仍未完全降解,烟草与生石花种子的发芽明显受到其内源抑制物的抑制,且对辣木与烟草幼苗生长也产生了明显抑制作用,但随着烟草幼苗的生长,植株对抑制物的耐受性增加,抑制效果减弱。
有机基质常通过复配无机材料和组分来达到优化基质理化性质的目的[23-26],本研究桉树皮基质通过复配不同比例珍珠岩发现:生料复配不同比例珍珠岩对耐受性较好的3种蔬菜(番茄、青花、生菜)种子的发芽无显著差异,而对抑制物耐受性较差的其他3种植物(辣木、烟草、生石花)有所不同,大多随着珍珠岩比例的增加,生料比例的减少而发芽率增加,这可能与无机材料的加入稀释了抑制物的相对含量有关。腐熟40 d桉树皮内由于抑制物含量相对更高,随着加入珍珠岩比例的增加,抑制物含量的相对减少,供试植物的发芽率也有所增加。腐熟60 d桉树皮内加入30%珍珠岩对大多试验植物种子的发芽率最佳。基质复配不同比例珍珠岩对试验植物苗期生长的影响与对发芽率的影响大致相同,不同在于腐熟40 d桉树皮加入20%珍珠岩对耐受性较好的3种蔬菜的苗期生长显著优于加入10%与30%珍珠岩的基质,这可能是该种处理基质内养分和抑制物对幼苗共同作用相互平衡的结果。
综上所述,腐熟60 d桉树皮可以替代泥炭土应用于番茄、青花、生菜的栽培育苗,其中又以复配30%珍珠岩的效果更佳,而对于辣木与烟草而言,腐熟60 d桉树皮还达不到泥炭土的效果,推测其主要原因在于基质内的抑制物,如果将该基质应用于这两种植物,还应通过优化发酵腐熟工艺进一步降低其内源抑制物含量至适合这一类低耐受性植物。对生石花而言,腐熟60 d桉树皮的效果略低于泥炭土,可通过与泥炭土的成本比较来决定是否采用该基质。本研究初步确定了腐熟桉树皮可作为基质主料应用于一些高耐受性植物育苗,至于各类经济植物对桉树内源抑制物耐受程度的临界值,以及将腐熟桉树皮基质应用于大部分经济植物育苗,还值得进一步深入研究。
参考文献
刘子刚, 王 铭, 马学慧. 世界泥炭地有机碳储量和有机碳密度[J]. 湿地科学, 2014, 12(3): 279-285.
Weiss R, Alm J, Laiho R, et al. Modeling moisture retention in peat soils[J]. Soil Sicence Society of America Journal, 1998, 62(2): 305-313.
Larsfranzén, 朱志辉. 湿地是解开冰期循环之谜的关键吗?[J]. AMBIO-人类环境杂志, 1994, 23(Z1): 300-308.
Xu J M, Bai J Y, Lu Z H. Some sustainable strategies of improvement and breeding for Eucalyptus tree species in southern China[J]. Forest Research, 2001, 14(6): 587-594.
Huang H Y, Zhao C J, Jiang X J, et al. Study on insect diversity in Eucalypt plantations[J]. Guangxi Forestry Science, 2011, 40(4): 292-295.
龙 波. 桉树-山毛豆复合经营种间生长影响及根系空间竞争研究[D]. 长沙: 中南林业科技大学, 2017.
Batish D R, Singh H P, Setia N, et al. Chemical composition and phytotoxicity of volatile essential oil from intact and fallen leaves of Eucalyptus citriodora[J]. Zeitschrift fur Naturforschung C, A Journal of Biosciences, 2006, 61(7-8): 465-471.
Dhakad A K, Pandey V V, Beg S, et al. Biological, medicinal and toxicological significance of Eucalyptus leaf essential oil: a review[J]. Journal of the Science of Food and Agriculture, 2018, 98(3): 833-848.
Ens E J, Bremner J B, French K, et al. Identification of volatile compounds released by roots of an invasive plant, bitou bush (Chrysanthemoides monilifera spp. Rotundata), and their inhibition of native seedling growth[J]. Biological Invasions, 2009, 11(2): 275-287.
Benchaa S, Hazzitb M, Abdelkrima H. Allelopathic effect of Eucalyptus citriodora essential oil and its potential use as bioherbicide[J]. Chemistry & Biodiversity, 2018, 15(8): 522-531.
Barbosa L C A, Filomeno C A, Teixeira R R. Chemical variability and biological activities of Eucalyptus spp. essential oils[J]. Molecules, 2016, 21: 1671.
劉曼红. 林业“三剩物”的开发利用现状和前景概述[J]. 林业调查规划, 2010, 35(3): 62-63, 67.
尚秀华, 谢耀坚, 彭 彦, 等. 腐熟桉树皮基质对桉树育苗效果的影响[J]. 中南林业科技大学学报, 2011, 31(6): 33-38, 43.
李思远, 余雪标, 陈候鑫, 等. 桉树皮对小白菜种子发芽的影响[J]. 热带生物学报, 2018, 9(2): 219-224.
刘纪霜, 亢希然, 李雪枝, 等. 桉树皮栽培秀珍菇配方筛选试验[J]. 中国食用菌, 2016, 35(6): 73-75.
陈丽新, 黄卓忠, 陈振妮, 等. 桉树皮、桉树屑栽培平菇营养成分和重金属分析[J]. 南方农业学报, 2015, 46(10): 1883-1886.
亢希然, 刘纪霜, 谢日禄, 等. 桉树皮栽培姬菇试验[J]. 食用菌, 2013, 35(6): 31-32.
黄卓烈, 林韶湘, 谭绍满. 桉树体内的生根抑制物质研究综述[J]. 林业科学研究, 1994(3): 319-324.
巫光宏, 詹福建, 黄卓烈, 等. 桉树LH21无性系生根抑制物初步研究[J]. 亚热带植物科学, 2002(3): 1-5.
王华宇, 陈丽文, 杨利平, 等. 辣木的种子特性及发芽试验[J]. 黑龙江农业科学, 2018, 291(9): 68-72.
李华君, 周 料, 唐恒孟. 广西东门林场尾巨桉育苗基质的研究[J]. 科技展望, 2014, 301(8): 43.
张沛健. 桉树皮废弃物腐熟技术及其在育苗中应用的研究[D]. 北京: 中国林业科学研究院, 2011.
陈 阳, 林永胜, 周先治, 等. 不同育苗基质对番茄幼苗生长的影响[J]. 热带作物学报, 2015, 36(12): 2149- 2154.
王秀雪, 惠成章, 张 维. 草炭替代物及其复配基质的理化性质研究[J]. 园艺与种苗, 2017, 77(11): 25-27, 34.
苏 飞. 椰糠复合基质在番茄无土栽培上应用与推广[D]. 福州: 福建农林大学, 2014.
杨敬华, 武占会, 刘明池, 等. 无机基质配比及沼渣施肥水平对无土栽培苗用型大白菜生长的影响[J]. 中国蔬菜, 2013, 280(6): 44-49.
责任编辑:沈德发
