遮阴对茶叶品质和产量影响研究进展
2021-09-10陈家铭吴淑华曾兰亭东方辜大川
陈家铭 吴淑华 曾兰亭 东方 辜大川

摘要:茶树在生长过程中常受到高光和高温等不良环境因素的影响,进而影响茶叶的品质与产量。遮阴作为茶园管理中常用的农艺措施,可以通过改善茶园微环境,尤其是光环境,从色泽、香气和滋味等方面提高茶叶的品质,应用前景广泛。文章综述了近年来关于遮阴對茶叶品质和产量的研究进展,从代谢物水平和基因调控水平阐释遮阴对茶叶色泽、香气和滋味的影响,及遮阴环境下茶树的生理响应与产量变化,并探讨了茶树遮阴操作中存在的问题及未来的研究方向,为进一步的研究与应用提供参考。
关键词:茶树;遮阴;品质;产量
Research Progress on the Influence of Shading on
Tea (Camellia sinensis) Quality and Yield
CHEN Jiaming1,2, WU Shuhua1,2, ZENG Lanting1, DONG Fang3, GU Dachuan1*
1. South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China;
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. Guangdong Food and Drug Vocational College, Guangzhou 510520, China
Abstract: Tea plants (Camellia sinensis) are often affected by adverse environmental factors such as high light andhigh temperature during the growth process, which in turn affect the tea quality and yield. Shading is a commonly agronomic measure in the management of tea gardens. It could improve the quality of tea in terms of color, aroma andtaste by enhancing the microenvironment of the tea garden, especially the light environment, which has a broadapplication prospect. This article reviewed the research progress of shading on tea quality and yield in recent years,discussed the effects of shading on tea color, aroma and taste in view of the metabolites and gene regulation, as wellas the physiological response and yield changes of tea plants in a shade environment. In addition, this article also discussed the problems and future research directions in tea plant shading operations, and provided a reference forresearch and application in the future.
Keywords: Camellia sinensis, shading, quality, yield
茶树(Camellia sinensis)属山茶科山茶属,是我国重要的木本经济作物之一,同时也属耐荫植物,喜阴、喜湿、喜漫射光。《大观茶论》中写道:“植茶之地,崖必阳,圃必阴……阴阳相济,则茶之滋长得其宜”。茶是以茶树上的芽叶嫩梢为原料加工制成的饮品,是世界上流行范围最为广泛的非酒精型饮料,仅次于水。茶叶的品质和产量与外界环境条件密切相关,其中光照是影响茶叶品质和产量的重要环境因素之一。春季,茶树体内氮代谢旺盛,进而影响春茶的滋味物质,例如具有鲜爽味的氨基酸含量较高,而苦涩味重的茶多酚和花青素等物质的含量较低,因而春茶是制作绿茶的极佳材料。但是,南方春季常发生“倒春寒”,茶嫩梢易受冻害而减产。因此,如何增强春季茶园抗霜冻能力从而提高春季茶叶产量和经济效益,是茶产业关注的问题之一[1-2]。与春季相反,茶树在夏秋季的高温干燥环境下碳代谢旺盛,氮代谢受抑制,不利于品质的形成。但茶树在夏秋季生长迅速,茶叶产量占全年总产量的60%,然而因品质差,每年都有大量的夏秋茶未被采摘利用而造成资源浪费。因此,如何提高夏秋茶品质从而增加茶叶利用率及经济效益,也是茶产业亟需解决的问题之一[3-4]。长期生产实践发现,对茶树进行遮阴处理可有效解决以上两个产业问题。大量研究发现,遮阴不仅能增强茶树抗霜冻能力,还可明显提高夏秋茶的色泽、香气和滋味等品质[5-7]。但也有研究发现,长期遮阴和重度遮阴会引起茶叶减产,表现为新梢鲜重、干重及百芽质量降低,叶片变薄等[8-9],因此遮阴时期及遮阴程度的控制尤为重要。
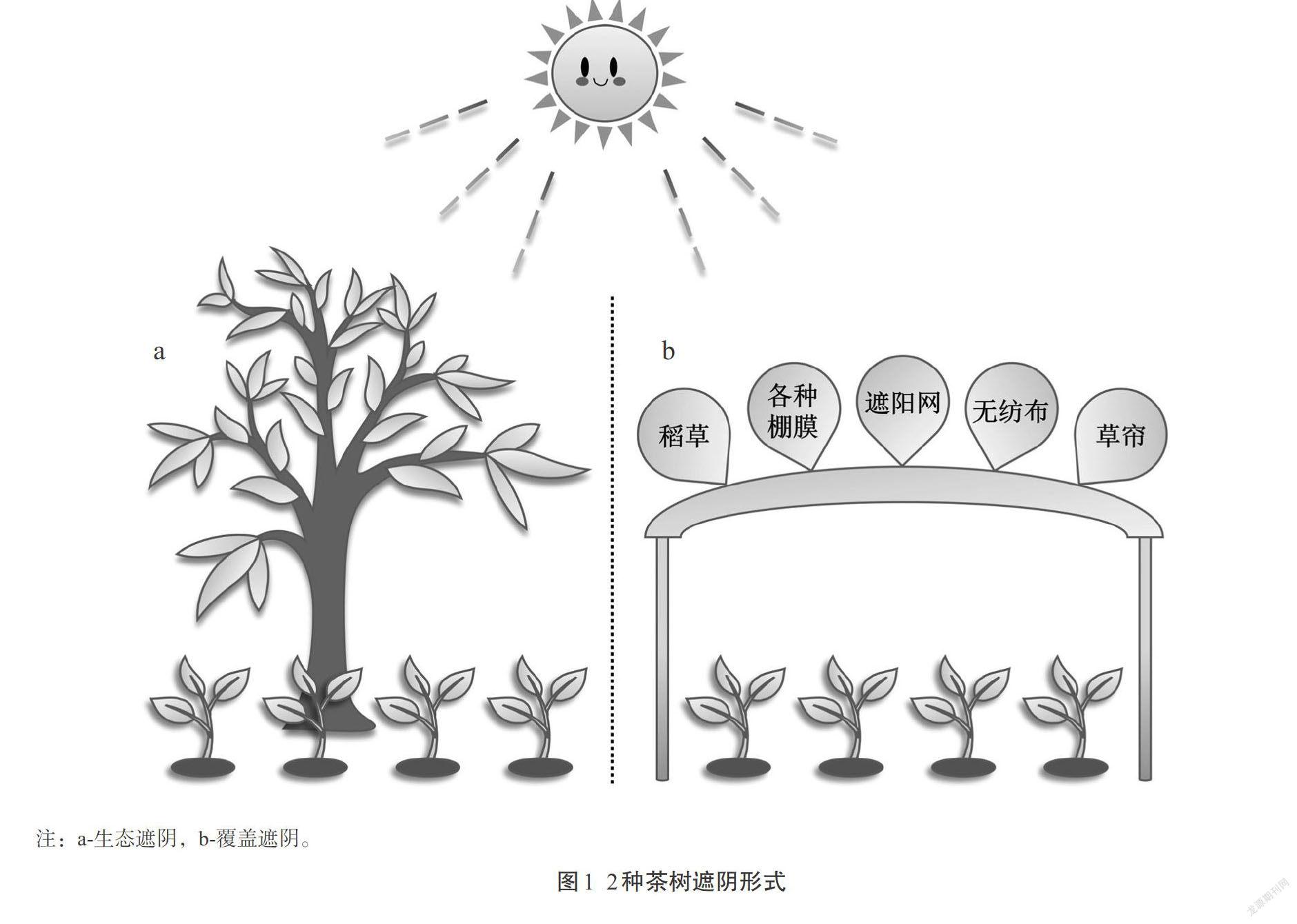
茶树遮阴有2种形式:生态遮阴和覆盖遮阴(图1)。生态遮阴指在茶园内种植遮阴树或与经济林木和高大适生树种间作,包括林茶间作、果茶间作及胶茶间作等;覆盖遮阴主要包括塑料大棚覆盖和遮阳网覆盖,所用材料较为广泛,例如稻草、小麦秸秆、草帘、各种树枝、无纺布(如帆布等)、各种棚膜、遮阳网等[7,10]。已有报道研究指出,遮阴可调节茶园微生态环境,如:降低茶树冠层温度、土壤温度及有效光合辐射强度,提高散射辐射的比例、改善光质、增加冠层湿度和土壤湿度及孔隙度[11-12]。遮阴还可增强土壤各种酶活性,促进土壤营养元素的循环利用,提高有机质和有效态C、N、K含量,从而提高土壤肥力[13-14]。目前,遮阴已成为一项重要农艺措施,在茶叶生产中发挥愈来愈重要的作用。为此,本文综述遮阴措施对茶叶色泽、香气和滋味等品质和产量影响的研究进展,以期为今后进一步研究及应用茶园遮阴提供参考。
一、遮阴对茶叶色泽的影响
茶叶的色泽品质包括干茶色泽、叶底色泽与茶汤色泽,不同茶类对于色泽的要求各不相同,如在名优绿茶的感官评审中,要求茶叶具有外形色泽绿、汤色绿、叶底绿的“三绿”品质[6]。大量研究發现,遮阴会引起茶叶叶绿素蓄积,具体表现在叶绿素a、叶绿素b的含量升高和叶绿素a/b值的显著降低[8]。叶绿素a/b值是衡量植物耐阴性的指标之一,比值越低,光捕获能力越强,有利于茶树吸收环境中的散射光,这种化合物变化是茶树适应遮阴环境的结果[15]。此外,遮阴还能调控叶绿体发育。Liu等[16]发现遮阴14 d后茶树叶绿体数量变多、形态变圆润、类囊体数量更多、堆叠更紧密,造成细胞内淀粉粒增加。对叶绿素合成前体物质进一步研究发现,遮阴处理后,短期内镁原卟啉IX(My-Proto IX)积累量迅速增加,而长期处于遮阴环境下,二乙烯原叶绿素酸酯(DPP)明显增加[6,16]。除了叶绿素,类胡萝卜素也是影响叶片颜色的主要色素。在黄化种和正常种中均发现遮阴会引起类胡萝卜素的蓄积,然而涨幅不及叶绿素变化幅度[17-18]。此外,叶黄素也会响应遮阴而发生积累[19]。
除植物光合色素外,茶叶中的茶黄素(TFs)对茶叶色泽品质也有贡献,其颜色为橙色或橙红色,一般认为它是茶叶多酚氧化聚合的产物,是儿茶素在多酚氧化酶(PPO)的作用下转化成的二聚物。Yu等[20]在采摘前对茶树进行遮阴处理,发现遮阴降低了大部分儿茶素的含量,却增加了茶黄素及其衍生物的含量。这可能是因为遮阴环境激发了PPO的酶活性和基因表达,尤其是CsPPO3的表达水平被显著提高。
综上,遮阴对茶叶色泽的影响是多方面的,不仅通过影响叶绿素和类胡萝卜素等光合色素的代谢水平,甚至还能通过酚类化合物的代谢途径影响色泽品质。近年来,越来越多的研究人员聚焦遮阴下茶叶色泽提升背后的科学问题。Liu等[16]通过结合转录组和代谢组学分析发现光信号途径的受体蛋白及转录因子可能通过植物激素共同参与茶树在遮阴环境下叶绿素蓄积的过程,研究结果为进一步解析遮阴对茶叶色泽形成的调控机制提供了前期基础。
二、遮阴对茶叶香气的影响
香气(挥发性化合物)是指干茶或茶叶冲泡时散发出的芳香气味。一般来说,酚类和氨基酸分别占总化合物的18%~36%和1%~4%,而香气物质仅占不到0.03%[21]。目前,利用气相色谱(GC)、气质联用色谱(GC-MS)、核磁共振(NMR)等技术手段从茶中已经鉴别和分离了700多种香气物质[22]。尽管茶叶中含有丰富的香气物质,但是只有浓度超过嗅觉阈值的物质才能贡献茶香的品质,且人类对不同香气物质的嗅觉阈值存在差异。常见的茶叶香气物质可以分为挥发性脂肪酸衍生物(VFADs)、挥发性萜烯类(VTs)、挥发性苯丙烷类/苯环类(VPBs)。香气作为茶叶的主要感官特性之一,影响着人们对茶叶的选择,高品质的茶叶香气呈现持久、浓烈、纯正的特点[22-23]。早在上个世纪,沈生荣等[24]就利用气相色谱技术发现遮阴后茶叶中香气成分有不同程度的明显提高,包括芳樟醇、橙花醇、香草醇、香叶醇、茉莉酮及其甲酯,以及百里香酚等物质。Yang等[25]利用代谢组学技术发现遮阴可以显著提高VPBs的含量,如苯甲醛、苯甲醇和2-苯乙醇,这些物质贡献了茶叶的花香味;该研究也进一步发现,这些物质在遮阴条件下增加的原因是由于它们共有的上游前体L-苯丙氨酸含量的上升。此外,其他研究中也发现遮阴可以通过增加香气物质及其前体的种类和含量来改善茶叶的香气品质[26-27]。
遮阴不仅会影响光强,还会改变环境中的光质,近年来的研究发现光质的改变也会显著影响茶叶香气组成。Fu等[28]发现蓝光(470 nm)和红光(660 nm)能够显著提高采前茶叶中包括VFADs、VPBs和VTs在内的大多数挥发物质的产生,主要是通过诱导相关合成基因的表达,而这2种光质处理对采后茶叶的影响却不大。虽然有别于传统遮阴,但是这一发现也为实际生产中提高茶叶品质提供了新的思路和方法。
三、遮阴对茶叶滋味的影响
滋味是指人的味觉感官对茶叶呈味物质的综合反映,茶叶滋味包括茶汤的苦、涩、酸、甜、鲜等因子,与香气共同构成茶叶品质中的核心要素[29]。影响茶叶滋味的物质有茶多酚、咖啡碱、氨基酸等,其中茶多酚是茶叶中酚类化合物的总称,包括儿茶素类、花青素、黄酮类和酚酸四大类化合物。儿茶素类是茶多酚中最主要的成分,占其总量的70%~80%[30]。儿茶素具有较强的苦味,是茶味中生津回甘和涩味的主要来源[23]。咖啡碱则贡献了茶汤中的苦味和鲜爽味[31],而氨基酸是茶叶中鲜爽味和甜味的主要物质。茶叶中的茶氨酸含量最高,占游离氨基酸的50%以上。氨基酸不仅在口味上中和了茶多酚和咖啡碱的苦涩味,还能演化出多种香气[32]。
研究发现,遮阴能改善茶叶的内含成分。顾辰辰等[33]试验发现,遮阴后的茶叶中茶多酚含量下降,儿茶素类化合物总量也随着遮阴程度的加重和遮阴时间的延长呈现下降趋势。对遮阴后8种茶叶中的儿茶素含量进行分析后发现,酯型儿茶素EGCG和ECG有升高的趋势,而大部分非酯型儿茶素如EGC和EC均发生不同程度的下降。该研究结果与孙京京等[34]的试验结果一致,即遮阴后儿茶素品质指数明显提高。儿茶素品质指数能反映鲜叶的嫩度和品质,是茶叶品质鉴定的早期生化指标,它的增加表明芽叶持嫩性增强,茶青品质高[31]。除了影响儿茶素类化合物总量,遮阴还会降低紫叶茶树总花青苷和黄酮组分(如槲皮素、山奈酚和杨梅素)含量[35],引起大部分黄酮(醇)糖苷含量显著下降[36],并且下调编码类黄酮生物合成途径关键酶的基因转录水平,包括苯丙氨酸解氨酶(PAL)、二氢黄酮醇还原酶(DFR)、花青素还原酶(ANR)和查尔酮合成酶(CHS)等关键限速酶基因[37]。有趣的是,在黄化品种中还发现遮阴会引起儿茶素和黄酮醇的空间分布发生改变[38]。
遮阴不仅可以通过降低茶多酚来减弱茶中的苦涩味,还可以通过提高游离氨基酸的含量提高茶叶鲜爽味和甜味,丰富茶的滋味与口感。茶氨酸是茶叶中的主体氨基酸成分,遮阴后茶氨酸含量显著提高,茶氨酸代谢途径的合成前体谷氨酸也明显蓄积,然而合成关键基因茶氨酸合成酶基因(CsTS)的表达量随着遮阴时间的延长变化不大,具体调控机制仍不清晰,尚需进一步研究[33]。而其他游离氨基酸如苏氨酸、组氨酸、天冬酰胺、天冬氨酸、精氨酸、苯丙氨酸等都有不同程度的提高[19,39],游离氨基酸含量升高可显著降低酚氨比。酚氨比是评价绿茶滋味的主要因子,遮阴后的鲜叶酚氨比低,更适合制作绿茶。此外,遮阴还可提高鲜叶中咖啡碱和可溶性糖的含量,从而提高茶汤的滋味品质[8,40]。其中,遮阴对于咖啡碱的影响可能受物种影响较大,在部分品种中咖啡碱变化不明显,甚至有遮阴后咖啡碱含量降低的报道[17]。
遮阴作为一种茶叶生产中的重要农艺措施,其提质效果已有大量的数据和文献支撑。在我国,由于夏秋茶品质差,大量的茶叶资源未能得到及时采摘和利用,造成巨大的经济损失。在夏季高温时节,遮阴措施在茶园管理中的应用可以很大程度提升茶叶的滋味,改善茶叶品质,具有直接的经济效益,然而遮阴在实际生产上的应用常常受限于人力成本而未能得到充分推广。此外,遮阴措施对春茶品质的提高作用十分有限,有研究发现春季遮阴引起茶叶氨基酸含量减少,咖啡碱含量水平基本不变甚至减少[41],这可能与产区气候以及春茶特性(酚氨比低)有關。不同产区地形、气候、季节、栽培品种耐阴特性、新梢萌育情况和生产需要等因素对茶树遮阴的程度与方式都有着不同的要求,这也进一步限制了遮阴措施应用推广。因此,解析遮阴对茶树提质的具体机制,对于指导实际生产具有深远的意义。然而,对机制的阐明往往受限于茶树遗传转化体系的不完善,至今仍有大量的研究空白。
四、遮阴对茶叶产量的影响
关于遮阴对茶树产量的影响受产地、气候、品种、遮阴程度与方式等因素的制约,至今未有统一的结论。目前存在2种观点:适度遮阴改善了茶园小气候、优化生长条件进而促进茶树生长;遮阴对品质的提升效应随着遮阴程度的加重而逐渐加强,在重度遮阴下茶叶品质更好,然而产量降低。其中,比较统一的结论是遮阴会提高新梢的含水率,造成茶叶干物率下降,影响产量。单武雄等[39]对秋茶进行遮阴处理,发现遮阴会引起茶树百芽质量、芽密度和单位面积产量呈现下降趋势;而秦志敏等[4]对春茶和夏茶进行遮阴处理却得到相反的结果:百芽质量和芽密度都显著提高。造成这种截然不同的结果可能是因为季节、气候、遮阴方式和品种耐阴特性的差异,甚至可能是统计标准的原因。翁伯琦等[42]在同一产区的不同月份进行遮阴处理也得到2种相反的结果,这暗示着遮阴对于茶树产量的影响可能受到多种因素的调控,因此,在实际生产中需要因地制宜。以日本的玉露为例,玉露茶主要用于抹茶的加工,由于产品对叶片叶绿素含量有较高的要求,在玉露种植过程中遮阴处理是必不可少的环节。然而,为了追求更好的品质,实际生产中往往采用遮阴率85%以上的重度遮阴,在生产中常常表现为产量降低、休眠芽比例升高、叶片变薄、单位面积叶片干质量降低等特征[8]。这种减产效应还会因反复遮阴而加剧,但该现象并不会影响第二年的产量[40]。
除了直接影响新梢的密度和质量,遮阴还可通过改变叶片结构影响新梢形态,间接影响产量。研究发现,遮阴会引起叶片及其表皮层、角质层和栅栏组织厚度变薄;新梢变短;叶面积增大;栅栏组织层数变少,排列疏松;海绵组织中间隙变多;栅栏组织/海绵组织比值变小[8-9,15,43]。这种形态结构的变化也与植物在荫蔽环境下表型变化相一致。遮阴后,茶树叶片除了形态上的变化,叶片质地也变得更柔软,持嫩性强。持嫩性是指茶叶保持嫩度的能力,在茶树品种选育过程中“持嫩性”是重要的衡量标准之一[44]。研究发现遮阴引起的茶叶粗纤维含量降低、木质素减少和含水量升高,可能是影响茶叶持嫩性的直接原因[41,45]。叶片嫩度高可能会影响茶叶的防御能力,然而对叶片表皮角质层成分进行分析后发现,遮阴不会影响角质合成,这侧面反映出茶树对低光强的高度适应性[46]。
遮阴引起产量变化的根本原因是遮阴改变了茶树的生长环境。茶园遮阴能显著降温增湿,使茶树免遭不良天气对生长的抑制。大量研究证实,遮阴会降低茶树冠层空气温度、叶片温度和土壤温度,提高茶树冠层空气湿度;降低茶树呼吸速率,提高净光合速率;气孔导度的增加加速了叶片内外的气体和水分交流,使胞间CO2浓度和蒸腾速率都呈上升趋势。此外,不同于露天环境下茶树的“午休”现象,中度遮阴环境下茶树净光合速率日变化进程呈现单峰曲线,还降低茶树光补偿点和光饱和点,暗示茶树在遮阴环境下更适合有机物的积累[43,47]。对茶树体内碳氮化合物的分析发现,遮阴后总氮含量增加,总碳含量减少,说明遮阴促进茶树氮循环,抑制碳循环[40]。然而,也有人提出不一样的看法,Li等[17]通过对碳氮代谢中间代谢产物的分析发现,遮阴可以抑制叶片的糖酵解途径和三羧酸循环,短期遮阴会一定程度促进叶片的氮循环,而长期遮阴无论对叶片还是整个新梢的氮循环都是抑制的,这与一般认为的遮阴可以促进氮循环的观点有所不同。
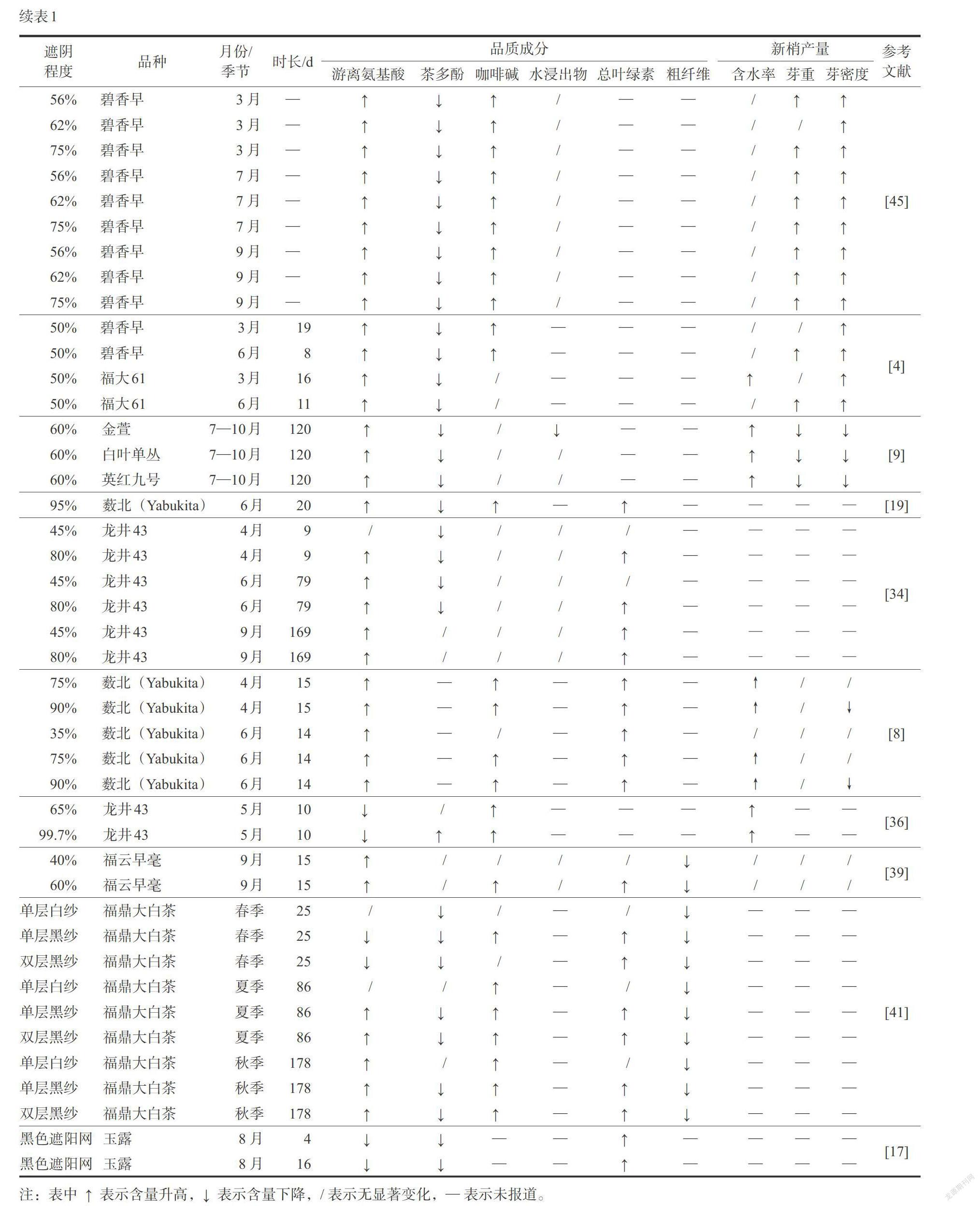
为了更加清晰地认识遮阴措施对茶叶品质和新梢产量的综合效益,整理前期研究中不同遮阴程度、不同品种、不同时期和处理时长下,遮阴处理对茶叶品质和新梢产量影响如表1。虽然遮阴对茶叶部分品质的提升受环境和茶树品种的影响较大,但整体效果仍呈现随遮阴程度的提高提质效果越好的趋势,具体表现在遮阴显著提高了茶叶中叶绿素和游离氨基酸水平,降低茶多酚含量,改善茶叶的色泽、香气和滋味品质。此外,遮阴对茶树产量的影响存在争议,部分文章报道遮阴提高了新梢产量,然而总体上随着遮阴程度的加剧新梢产量降低。
五、遮阴影响茶树生理响应
上游调控机制探讨
遮阴是茶树长期生产实践中一直沿用的农艺措施,关于遮阴对茶树色泽、香气、滋味和产量的影响已经开展了大量的研究,但这些研究多集中于对色泽、香气、滋味和产量改变的描述,鲜少涉及机理方面的研究。近年来,随着茶树基因组的公布[48-49],分子生物学技术的进步,通过代谢组[37]、蛋白质组[26]、转录组[37]和DNA甲基化组[50]等组学手段初步解析了遮阴措施如何影响茶叶品质或功能相关代谢物形成及其基因调控网络,例如:遮阴引起的游离氨基酸蓄积主要来自于叶绿体蛋白的蛋白水解,而非来自游离氨基酸的合成[26];遮阴通过影响UVR8介导的信号途径中MYB12/MYB4下游因子调控类黄酮的蓄积[37];使用互补的非靶向代谢组学方法,在幼嫩的茶苗组织(茎、第一叶和第四叶)中检测到1 419种代谢物响应两种不同的遮阴处理。遮阴处理后,茶氨酸在第一叶和第四叶中均显著增加,而茎中的茶氨酸则显著下降,并且遮阴处理对氨基酸/儿茶素比的积极影响在第四叶中比第一叶中更为明显[44];全基因组DNA甲基化分析表明,遮阴处理会引起全基因组DNA甲基化模式发生显著变化,联合转录组的分析则表明,差异甲基化区域相关的差异表达基因是主要次生代谢产物生物合成的关键参与者[50]。
尽管多组学技术联用已初步描绘了遮阴措施对于茶树的综合影响,但是目前针对某一性状或者代谢物的调控研究仍较少,大量的上游调控因子,如转录因子等,有待发掘。此外,这些调控因子响应遮阴措施的具体机制也有待进一步研究解析。
对于植物而言,光是光合作用和正常生长发育所需的环境因子,但是相邻的植株冠层或遮阴材料会造成R∶FR(红光∶远红光)比例以及光合有效辐射强度的降低,从而影响植物的生长发育[51]。长期以来,植物自身发展出多种光受体来感知外部光环境的变化,检测竞争者的存在,从而相应地调整其生长发育策略。一般而言,根据叶片对遮阴的不同反应,植物可分为两类:避阴和耐阴[51]。植物的避阴反应(SAS)是指当植物感知到周围环境遮阴时,会表现出节间伸长、分支减少、叶面积减小、开花加快等特征,以更快的生长速度突破遮阴环境,获得更多光资源,这是生长相对密集的植物群体在长期的演变进化过程产生的一种竞争光资源的策略[52]。模式植物拟南芥就属于典型的避阴植物,当拟南芥遭受遮阴胁迫后,叶绿素含量减少并且参与光合作用的基因表达下调[53]。与此相反,当茶树遭受遮阴胁迫后,叶绿素a和b含量增加但a/b比值减小,相应的光合作用基因表达上调[16]。茶树遮阴后的这些表現与植物耐阴反应(STR)的表型相一致[54],也与长期以来认为茶树是耐阴植物的观点相符。耐阴植物往往通过其他不同于避阴反应的生理和代谢变化来适应有限资源的环境。如生活在热带树冠下的秋海棠属植物具有特化的表皮叶绿体,这些特化叶绿体一方面增强了植物在主要为绿光的弱光环境下的光能捕获效率,另一方面提高5%~10%的量子产率,进而提高光合作用产量[55]。近年通过比较来自不同生境的野生天竺葵发现,即便是亲缘关系较近的物种,遮阴与避阴物种在防御基因的调控、生长和基因表达模式上都存在着明显的差异[56]。
近期关于不耐阴的苏格兰松树和耐阴的挪威云杉的研究则表明,R∶FR比值触发了挪威云杉对遮阴的响应,而在苏格兰松树中则观察到R∶FR比值和光强的相互作用,进一步的RNA-seq数据则显示,SAS和STR响应包括主要参与激素信号转导和色素生物合成的基因表达变化[53]。同年发表的另一项研究则表明,与近亲拟南芥不同,在遭受遮阴胁迫时,耐阴的碎米芥通过进化出一种更高效的phyA依赖途径,抑制下胚轴的伸长[57]。尽管近年来对于植物耐阴的机制有了以上初步的探索,但是相比于避阴植物中的研究,耐阴植物的研究基础仍相对薄弱,而以茶树作为耐阴植物所开展的研究目前尚未有报道。今后,对于茶树耐阴机制的解析,不仅有助于理解遮阴措施对于茶树的影响,更能为理解耐阴植物的适应性机制提供经典案例。
六、小结与展望
一般而言,各种遮阴措施均能在一定程度上降低树冠面光强和叶片温度,提高空气和土壤的相对湿度,从而改善茶树生长的微环境。遮阴后,在不同季节不同茶树品种中,色泽、香气和滋味等品质性状均有一定程度的改良,主要表现为叶绿素和β-胡萝卜素含量显著提高,挥发性脂肪酸衍生物和挥发性苯丙烷/苯甲酸酯显著提高,茶氨酸等氨基酸和咖啡碱等组分含量显著增加,茶多酚和总儿茶素含量显著降低。在产量与性状方面,遮阴对茶树的影响则呈现出不统一的规律,推测可能的原因与遮阴材料、遮阴强度、茶树品种、遮阴季节和时长均有一定关联性。鉴于遮阴对于茶叶品质的显著提升作用和对于茶叶产量影响的不确定性,未来的工作应重点解析遮阴影响新梢产量和茶叶品质的上游因子,为达到茶叶“保产提质”或“增产提质”目标提供理论基础。
参考文献
[1] 陶汉之, 王镇恒. 我国茶树光合作用研究进展及发展趋势[J]. 茶叶科学, 1995, 15(1): 1-8.
[2] 胡永光, 江丰, ASHRAF M, 等. 春茶采摘末期遮荫对其生长和品质的影响[J]. 农业机械学报, 2018, 49(1): 283-289.
[3] 师大亮, 郭敏明. 提高夏秋名优绿茶品质的技术措施[J]. 浙江农业科学, 2006 (6): 710-711.
[4] 秦志敏, JOHN TANUI, 冯卫英, 等. 遮光对丘陵茶园茶叶产量指标和内含生化成分的影响[J]. 南京农业大学学报, 2011, 34(5): 47-52.
[5] 黄海涛, 许永妙, 张伟, 等. 遮阳网覆盖方式对茶园霜冻害防御效果的比较研究[J]. 杭州农业与科技, 2014 (6): 38-39.
[6] 舒华, 王盈峰, 张士康, 等. 遮荫对茶树新梢叶绿素及其生物合成前体的影响[J]. 茶叶科学, 2012, 32(2): 115-121.
[7] 张文锦, 林春莲, 熊明民. 茶树遮荫效应研究进展[J]. 福建农业学报, 2007, 22(4): 457-460.
[8] SANO T, HORIE H, MATSUNAGA A, et al. Effect of shading intensity on morphological and color traits and on chemical components of new tea (Camellia sinensis L. ) shoots under direct covering cultivation[J]. Journal of Agricultural and Food Chemistry, 2018, 98: 5666-5676.
[9] 唐颢, 唐劲驰, 黎健龙. 高温干旱季节茶园覆盖遮荫的综合效应研究[J]. 广东农业科学, 2008(8): 26-29.
[10] 宋晓蕾, 杨红玉, 曾黎琼, 等. 植物遮荫效应的研究进展[J]. 北方园艺, 2009(5): 129-133.
[11] 陈佩, 杨知建, 肖润林. 遮阴对茶园生态环境及其茶树光合作用和产量的影响研究[J]. 安徽农业科学, 2010, 38(11): 5604-5605,
5639.
[12] 王雪萍, 龔自明. 夏秋季茶树遮阴效应研究进展[J]. 湖北农业科学, 2017, 56(23): 4447-4449, 4453.
[13] 段建真, 郭素英. 遮荫与覆盖对茶园生态环境的影响[J]. 安徽农学院学报, 1992, 19(3): 189-195.
[14] 王国夫, 孙小红, 方逸, 等. 遮阴对抹茶茶园土壤微生物特性及土壤酶活性的影响[J]. 茶叶科学, 2019, 39(3): 355-363.
[15] 张文锦, 梁月荣, 张方舟, 等. 覆盖遮荫对乌龙茶产量、品质的影响[J]. 茶叶科学, 2004, 24(4): 276-282.
[16] LIU L, LIN N, LIU X, et al. From chloroplast biogenesis to chlorophyll accumulation: the interplay of light and hormones on gene
expression in Camellia sinensis cv. Shuchazao leaves[J]. Frontiers in
Plant Science, 2020, 11: 256.
[17] LI Y, JEYARAJ A, YU H, et al. Metabolic regulation profiling of carbon and nitrogen in tea plants [Camellia sinensis (L.) O. Kuntze] in response to shading[J]. Journal of Agricultural and Food Chemistry, 2020, 68: 961-974.
[18] 田月月, 张丽霞, 张正群, 等. 夏秋季遮光对山东黄金芽茶树生理生化特性的影响[J]. 应用生态学报, 2017, 28(3): 789-796.
[19] LEE L S, CHOI J H, SON N, et al. Metabolomic analysis of the
effect of shade treatment on the nutritional and sensory qualities of
green tea[J]. Journal of Agricultural and Food Chemistry, 2013, 61:
332-338.
[20] YU Z M, LIAO Y Y, ZENG L T, et al. Transformation of catechins into theaflavins by upregulation of CsPPO3 in preharvest tea (Camellia sinensis) leaves exposed to shading treatment[J]. Food Research International, 2020, 129: 108842.
[21] 宛晓春. 茶叶生物化学[M]. 北京: 中国农业出版社, 2003.
[22] ZENG L T, WATANABE N, YANG Z Y. Understanding the biosyntheses and stress response mechanisms of aroma compounds in tea (Camellia sinensis) to safely and effectively improve tea aroma[J]. Critical Reviews in Food Science and Nutrition, 2019, 59: 2321-2334.
[23] 张欣然. 茶叶审评技术研究进展[J]. 中国野生植物资源, 2020, 39(12): 46-51.
[24] 沈生荣, 杨贤强, 陈席卿. 遮荫对蒸青绿茶香气成分的影响[J]. 浙江农业大学学报, 1990, 16(1): 98-101.
[25] YANG Z Y, KOBAYASHI E, KATSUNO T, et al. Characterisation
of volatile and non-volatile metabolites in etiolated leaves of tea
(Camellia sinensis) plants in the dark[J]. Food Chemistry, 2012, 135:
2268-2276.
[26] CHEN Y Y, FU X M, MEI X, et al. Proteolysis of chloroplast proteins
is responsible for accumulation of free amino acids in dark-treated
tea (Camellia sinensis) leaves[J]. Journal of Proteomics, 2017, 157:
10-17.
[27] ZENG L T, ZHOU X, SU X G, et al. Chinese oolong tea: an aromatic
beverage produced under multiple stresses[J]. Trends in Food
Science & Technology, 2020, 106: 242-253.
[28] FU X M, CHEN Y Y, MEI X, et al. Regulation of formation of vola-
tile compounds of tea (Camellia sinensis) leaves by single light
wavelength[J]. Scientific Reports, 2015, 5: 16858-16864.
[29] 凌彩金, 王秋霜, 卓敏, 等. 茶叶审评技术研究进展[J]. 广东农业科学, 2010, 37(3): 68-71.
[30] 彭静, 孙威江. 茶叶苦味研究进展[J]. 食品工业, 2017, 38(1): 244-248.
[31] 叶乃兴. 茶叶品质性状的构成与评价[J]. 中国茶叶, 2010, 32(8): 10-11.
[32] 蔡翔, 李延升, 杨普香, 等. 茶氨酸呈味特征及应用前景[J]. 蚕桑茶叶通讯, 2017 (6): 31-33.
[33] 顾辰辰, 王荣秀, 江丽娜, 等. 短期遮阴对茶树嘌呤碱、氨基酸和儿茶素生物合成的影响[J]. 安徽农业大学学报, 2017, 44(1): 1-6.
[34] 孙京京, 朱小元, 罗贤静丽, 等. 不同遮荫处理对绿茶品质的影响[J]. 安徽农业大学学报, 2015, 42(3): 387-390.
[35] 金琦芳, 孙威江, 王仲. 遮阴处理对茶树叶色表型及生化成分的影响[J]. 食品工业科技, 2018, 39(23): 1-6.
[36] 陈勤操, 戴伟东, 蔺志远, 等. 代谢组学解析遮阴对茶叶主要品质成分的影响[J]. 中国农业科学, 2019, 52(6): 1066-1077.
[37] LIU L L, LI Y Y, SHE G G, et al. Metabolite profiling and transcriptomic analyses reveal an essential role of UVR8-mediated signal transduction pathway in regulating flavonoid biosynthesis in tea plants (Camellia sinensis) in response to shading[J]. BMC Plant Biology, 2018, 18: 233-238.
[38] ZHANG Q F, LIU M Y, RUAN J Y. Metabolomics analysis reveals
the metabolic and functional roles of flavonoids in light-sensitive
tea leaves[J]. BMC Plant Biology, 2017, 17: 64-69.
[39] 單武雄, 肖润林, 王久荣, 等. 遮光对丘陵茶园白露毛尖茶产量和品质的影响[J]. 农业现代化研究, 2010, 31(3): 368-372.
[40] YAMASHITA H, TANAKA Y, UMETSU K, et al. Phenotypic
markers reflecting the status of overstressed tea plants subjected to
repeated shade cultivation[J/OL]. Frontiers in Plant Science, 2020,
11: 1-15[2021-04-10]. https://www.researchgate.net/publication/
346735991_Phenotypic_Markers_Reflecting_the_Status_of_Over-
stressed_Tea_Plants_Subjected_to_Repeated_Shade_Cultivation.
[41] 刘建军, 袁丁, 司辉清, 等. 遮荫对不同季节茶树新梢的内含成分影响研究[J]. 西南农业学报, 2013, 26(1): 115-118.
[42] 翁伯琦, 张文锦. 乌龙茶覆盖遮荫技术的研究[J]. 厦门大学学报(自然科学版), 2005(s1): 16-21.
[43] 付晓青, 陈佩, 秦志敏, 等. 遮荫处理对丘陵茶园生态环境及茶树气体交换的影响[J]. 中国农学通报, 2011, 27(8): 40-46.
[44] 黄艳, 成浩. 茶树新梢“持嫩性”研究进展[J]. 安徽农业科学, 2011, 39(17): 10260-10262.
[45] 王玉花, 秦志敏, 肖润林, 等. 遮光水平对丘陵茶园茶叶生长指标和品质的影响[J]. 经济林研究, 2011, 29(2): 48-53.
[46] TSUBAKIS, SAKUMOTO S, UEMURA N, et al. Compositional
analysis of leaf cuticular membranes isolated from tea plants (Camellia sinensis L.)[J]. Food Chemistry, 2013, 138: 286-290.
[47] 趙甜甜, 蔡新. 不同遮荫度下茶树生理生化特性的研究[J]. 湖南农业科学, 2010 (5): 38-41.
[48] XIA E H, ZHANG H B, SHENG J, et al. The tea tree genome pro-
vides insights into tea flavor and independent evolution of caffeine
biosynthesis[J]. Molecular Plant, 2017, 10(6): 866-877.
[49] WEI C L, YANG H, WANG S B, et al. Draft genome sequence of
Camellia sinensis var. sinensis provides insights into the evolution of
the tea genome and tea quality[J]. Proceedings of the National A
cademy of Sciences, 2018, 115(18): 4151-4158.
[50] XU P, SU H, JIN R, et al. Shading effects on leaf color conversion and biosynthesis of the major secondary metabolites in the albino tea cultivar “Yujinxiang”[J]. Journal of Agricultural and Food Chemistry, 2020, 68(8): 2528-2538.
[51] FIORUCCI A S, FANKHAUSER C. Plant strategies for enhancing access to sunlight[J]. Current Biology, 2017, 27(17): 931-940.
[52] CASAL J J. Photoreceptor signaling networks in plant responses to shade[J]. Annual Review of Plant Biology, 2013, 64: 403-427.
[53] RANADE S S, DELHOMME N, GARCIAGIL M R. Transcriptome analysis of shade avoidance and shade tolerance in conifers[J]. Planta, 2019, 250(1): 299-318.
[54] GOMMERS C M, VISSER E J W, STONGE K R, et al. Shade
tolerance: when growing tall is not an option[J]. Trends in Plant
Science, 2013, 18(2): 65-71.
[55] JACOBSS M, LOPEZ-GARCIA M, PHRATHEP O P, et al. Photonic multilayer structure of Begonia chloroplasts enhances photosynthetic efficiency[J]. Nature Plants, 2016, 2(11): 1-6.
[56] GOMMERS C M, KEUSKAMP D H, BUTI S, et al. Molecular
profiles of contrasting shade response strategies in wild plants:
differential control of immunity and shoot elongation[J]. The Plant
Cell, 2017, 29(2): 331-344.
[57] MOLINA-CONTRERAS M J, PAULISIC S, THEN C, et al. Photoreceptor activity contributes to contrasting responses to shade in
Cardamine and Arabidopsis seedlings[J]. The Plant Cell, 2019, 31(11): 2649-2663.