湿润地区3种松属植物枝和根导水系统的效率-安全关系*
2021-09-06叶琳峰王忠媛陆世通潘天天谢江波
叶琳峰 李 彦,2,3 王忠媛,2,3 陆世通 潘天天 陈 森 谢江波,2,3
(1.浙江农林大学 省部共建亚热带森林培育国家重点实验室 杭州 311300;2.中国科学院新疆生态与地理研究所 荒漠与绿洲生态国家重点实验室 乌鲁木齐 830011;3.浙江农林大学林业与生物技术学院 杭州 311300)
过去几十年间,干旱引起的森林枯萎和死亡被广泛报道(Youngetal.,2017)。在干旱区(Doughtyetal.,2015)、温带森林(Andereggetal.,2015)和热带雨林(Truebaetal.,2017),植物生长和死亡都对干旱高度敏感。可见树木干旱致死并不局限于干旱地区,通常认为不存在水分限制的区域也可能会发生(Allenetal.,2010)。实际上,干旱区的树木能够适应一定的干旱胁迫(Andereggetal.,2015),而湿润区的树木因缺乏抗旱适应驯化或能力而对干旱胁迫更敏感(Jumpetal.,2017),更易发生水力失效(Choatetal.,2012)。一般来说,裸子植物比被子植物抗栓塞性更强(Maheralietal.,2004),但干旱同样也会导致它们死亡(Hartmannetal.,2013)。全球范围内,裸子植物干旱死亡及其导水系统栓塞抗性的研究多集中在干旱区(Larteretal.,2017),而且侧重植物单一器官(Boucheetal.,2014),如叶(Schumannetal.,2019)、茎(David-Schwartzetal.,2016)或根(Lossoetal.,2015)。而栓塞抗性具有器官间差异,因此理解水力效率在不同器官的协调关系就显得十分必要(Petersetal.,2020)。虽然同步检验茎、叶水力性状权衡关系的研究逐渐增多(Rosasetal.,2019),但将地上、地下部分结合考虑的研究依然非常有限。
植物木质部在有效运输水分的同时,自身也面临气穴栓塞的风险(陈志成等,2017),栓塞引起的水力失效可直接决定植物的生死(Brodribbetal.,2010;Choatetal.,2012)。木质部栓塞是木本植物在遭受水分胁迫时产生的导管输水功能障碍,可用“脆弱曲线”(vulnerability curves,VC)来描述其栓塞抗性(Tyreeetal.,1989)。该曲线根据木质部导水率损失的百分数(percent loss of conductivity,PLC)与相对应水势绘制而成(Domecetal.,2001),P50(导水率损失50%时的木质部水势)是脆弱曲线最陡处,常用来表征植物木质部栓塞抗性(Choatetal.,2012)。在植物水分关系研究中(Gleasonetal.,2016),一般用比导率(Ks,枝条或根单位边材面积的导水率)来定义水力效率(以下简称“效率”),同时将其水力安全性(以下简称“安全”)定义为栓塞抗性(P50)。效率-安全权衡假说认为,木质部的安全性(抵抗栓塞形成和扩散的能力)应与木质部的效率(输送水分的能力)相权衡,即水分运输效率高的物种易发生栓塞,导致其安全性低;而水分运输效率低的物种则更安全,栓塞抗性高(Hackeetal.,2006)。这一经典假说自其提出后一直被讨论和验证。Maherali等(2004)发现,相较于被子植物,裸子植物的效率-安全权衡更明显。但由于裸子植物具有独特的纹孔塞模式,这种权衡又可能被解偶联(Burgessetal.,2006),也确有研究发现二者并不存在权衡(Buccietal.,2013)。
气候或生境对整个植物的影响可以改变效率或安全性的相对优势,但不一定能改变木质部本身的结构特性带来的权衡(Gleasonetal.,2016)。木质部作为维管植物的水分运输组织,其结构是水力效率、水力安全的物质基础(Scholzetal.,2013),管胞和纹孔结构更是评估木质部功能性状的重要依据(Brodersenetal.,2014)。已有研究表明,除管胞直径外,管壁厚度(Hackeetal.,2001)、管胞密度(Frouxetal.,2005)和木材密度(Rosneretal.,2011)均与水力效率密切相关。此外,越来越多的证据表明,纹孔控制着栓塞在相邻导管间的扩散,是决定木质部水力导度和栓塞抗性的关键结构(Schulteetal.,2015)。因此,木质部结构与效率-安全的关系,是揭示树木干旱致死机制的基础。
本文聚焦于湿润区常见的3种松属植物的枝与根的水力性状,以确认是否存在地上、地下器官的木质部效率-安全权衡,及其存在是否与解剖结构有关。以便更深入认识植物干旱响应机制,为预测气候变化导致的树木干旱致死奠定基础。
1 材料与方法
1.1 研究区概况 试验地位于浙江省西北的天目山区(30°24′55″N,119°24′47″E)。受海洋暖湿气流与北方寒冷气流的交互影响,该地四季分明、雨水相对充沛但干旱(尤其伏旱)频发。年均气温14 ℃,最冷月平均气温-2.6~3.4 ℃,最热月平均气温19.9~28.1 ℃,年降水量1 390~1 870 mm,年均相对湿度76%~81%,年太阳辐射3 270~4 460 MJ·m-2。
1.2 研究材料 选取3种当地常见的松属树种:马尾松(Pinusmassoniana)、日本五针松(P.parviflora)和湿地松(P.elliottii),样本采集时间为2019年2月。每个树种随机选取立地一致、生长良好的3株样树,其株高、胸径、冠幅相似。在8:00—11:00间,用枝剪采集树冠中南部外层的基径6~8 mm、长40~50 cm的2年生枝条。在对应树冠方位下取根,即在10~20 cm土层选择直径3~5 mm、长20~40 cm侧根,小心剥离土壤避免根损伤,迅速放入装有水的黑箱中(以防止水分散失和外界空气进入切开的导管内),立即带回实验室。
1.3 研究方法 1)枝条(根)导水率及脆弱曲线测定 用木质部导水率与栓塞测量系统(XYL’EM-PLUS,Bronkhorst,Montigny-les-Cormeilles,France)测定导水率和导水损失率(样品其余部分用于木质部解剖结构、木材密度的测定,以确保结构性状与功能性状指标来自同一枝条)。
(1)将采回的样品在水下切割,用刀片把茎段两端的切口修平(去除充满空气的导管或管胞),剪取的枝干长度在10~20 cm。将茎段放入压力套(Model 1505D-EXP Pressure Chamber,PMS Instrument Company,OR,USA)中且两端露出,压力套密封后连接压力表,茎段近轴端与XYL’EM-PLUS连接(Kavanaghetal.,1999)。
(2)在水下切割茎段后立即用低压(3~5 kPa)测定茎段初始导水率(Kstand,kg m·s-1MPa-1),所用溶液为20 mmol·L-1KCl+1 mmol·L-1CaCl2(陈志成等,2018)。
(3)初始导水率(Kstand)测定后,用0.15~0.2 MPa的压力冲刷枝条,直至枝条管胞内无气泡溢出为止,然后测量最大导水率(Kmax,kg m·s-1MPa-1)(周洪华等,2015)。
(4)最大导水率(Kmax)测定后,使用空气注入法(Schumannetal.,2019),加压10 min以诱导栓塞形成,停止加压至压力充分释放后,测定相应导水率(Khi,kg m·s-1MPa-1),依此类推,直至导水损失率PLC=(1-Khi/Kmax)×100%达90%以上。
(5)紧接着,用数显卡尺测量枝条内径(去除树皮后枝条的直径)、长度。比导率(Ks):Kmax与单位边材面积(无髓无树皮)的比值(Schuldtetal.,2016)。
(6)所用压力和相应的PLC之间关系的曲线即为枝干的脆弱性曲线,通过Weibull函数拟合。用R软件中的“fitplc”包计算导水率损失50%时所对应的木质部水势P50,以此代表木质部栓塞抗性(Duursmaetal.,2017)。
对根的相关操作同上。
此外,本研究收集了2000—2019年间发表的松属植物效率-安全权衡的数据(这些研究均同时测量了植物输水效率和栓塞抗性指标),以甄别本试验结果的普适性。这些文献中研究对象多为自然生长的松属植物,少数为人工栽培或室内盆栽。
2)木质部解剖结构 从测定脆弱曲线的枝条上截取3~5 cm枝段,切成0.5 cm长的条状,放入FAA固定液保存。处理过的样品依次经过软化、脱水、包埋、切片、染色、粘片、胶封等步骤后,制成永久切片。使用Leica DM3000(Leica Microsystems CMS Gmbh,Germany)显微镜拍摄切片照片,观察木质部解剖结构。
(1)水力直径(Dh)计算 在Leica DM3000显微镜50×镜下获取管胞切面照片,每个切面上选取相对均匀的3个扇面(每个扇面30°),每个扇面沿射线细胞、由木质部外侧向内侧髓心方向拍照,用ImageJ软件直接从图片上获取D,每个扇面约有1 000~1 500个管胞(根每个扇面包含150~300个管胞)。Dh计算公式(党维等,2017):Dh(μm)=ΣD5/ΣD4。
(2)管胞密度(N)测量 在测量管胞直径的照片上,用ImageJ软件统计出单位面积内管胞数目和横截面的面积,每根枝条5个重复(Caietal.,2014)。N(103个·μm-2)=横截面所有管胞的数量/横截面的面积。
(3)单管胞壁厚度(T)测量 依据Hacke等(2001)的研究方法,在400×镜下拍摄管胞照片,测量T(μm)值,每根枝条5个重复。
(4)木材密度(WD)测量 每个枝条取2.5 cm枝段,测定枝段干质量和鲜体积,每根枝条5个重复(Hackeetal.,2000)。WD(g·cm-3)=干质量/鲜体积。
(5)纹孔特征测定 用Leica DM3000显微镜观察拍照,将径切面和弦切面切片放在400×镜下观察(Pittermannetal.,2006),每根枝条选取5个视野,每个视野10~15个重复。利用ImageJ软件进行测量,获得纹孔塞面积(pit torus area,Ato,μm2)、纹孔膜面积(pit membrane area,Apm,μm2),并计算塞缘面积(pit margo area,Am,μm2)(Boucheetal.,2014):Am=Apm-Ato。
对根的操作方法同上。
1.4 数据分析 性状单因素方差分析(one-way ANOVA,用LSD进行差异显著性检验)、结构性状和功能性状相关分析以及冗余分析(RDA)、图表制作等,均在R(版本3.5.3;R Core Team 2019)中完成。
2 结果与分析
2.1 水力功能性状分析 1)输水效率 3个树种枝条比导率(Kss)大小顺序为:日本五针松>马尾松>湿地松,日本五针松Kss显著大于马尾松和湿地松(图1A);3个树种根比导率(Ksr)有显著差异,大小顺序也为:日本五针松>马尾松>湿地松(图1B);3个树种根输水效率极显著大于枝条输水效率(图1)。此外,相关分析(图2)表明:各树种Kss和Ksr正相关。

图1 3个树种枝条和根的比导率Fig.1 Specific hydraulic conductivity of branch and root of the three Pinus species不同小写字母表示各树种枝条或根比导率差异显著(P <0.05)。Different lowercase letters indicate significant differences of specific hydraulic conductivity in branches and roots among species(P <0.05).

图2 3个树种枝条和根功能性状的相关分析Fig.2 Correlation analysis of functional traits of branch/root for the three speciesKss:枝条比导率;Ksr:根比导率;P50s:枝条栓塞抗性;P50r:根栓塞抗性。相关系数的显著性:*,P <0.10;**,P <0.05;***,P <0.01。下同。Kss:Branch specific hydraulic conductivity;Ksr:Root specific hydraulic conductivity;P50s:Branch embolism resistance;P50r:Root embolism resistance.Significance of correlation coefficient:*,P <0.10;**,P <0.05;***,P <0.01.The same below.
2)栓塞抗性 3个树种枝条P50差异极显著(图3A-C),表现为马尾松<湿地松<日本五针松。根P50大小为马尾松<日本五针松<湿地松,但3个树种根P50无显著差异(图3D-F)。3个树种枝条P50显著小于根P50。相关性分析后发现枝条P50和根P50正相关(图2)。
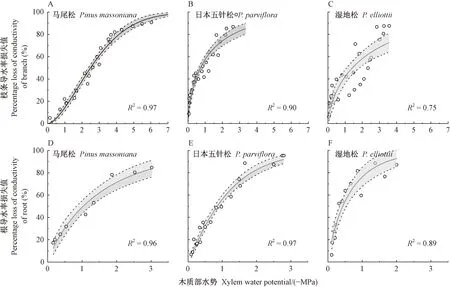
图3 3个树种枝条和根的脆弱曲线Fig.3 Branch and root vulnerability curves of the three Pinus species
另外,本研究整合了已发表数据与实测数据,分别对Kss与枝条P50、Ksr与根P50进行相关分析。结果(图4)表明:枝条水力效率与水力安全极显著负相关(R2=0.15,P<0.01),根水力效率与水力安全相关系数趋近于0(R2=0.01,P=0.94),即:生长于湿润区的松属树种枝条效率与安全有权衡,但根无权衡。
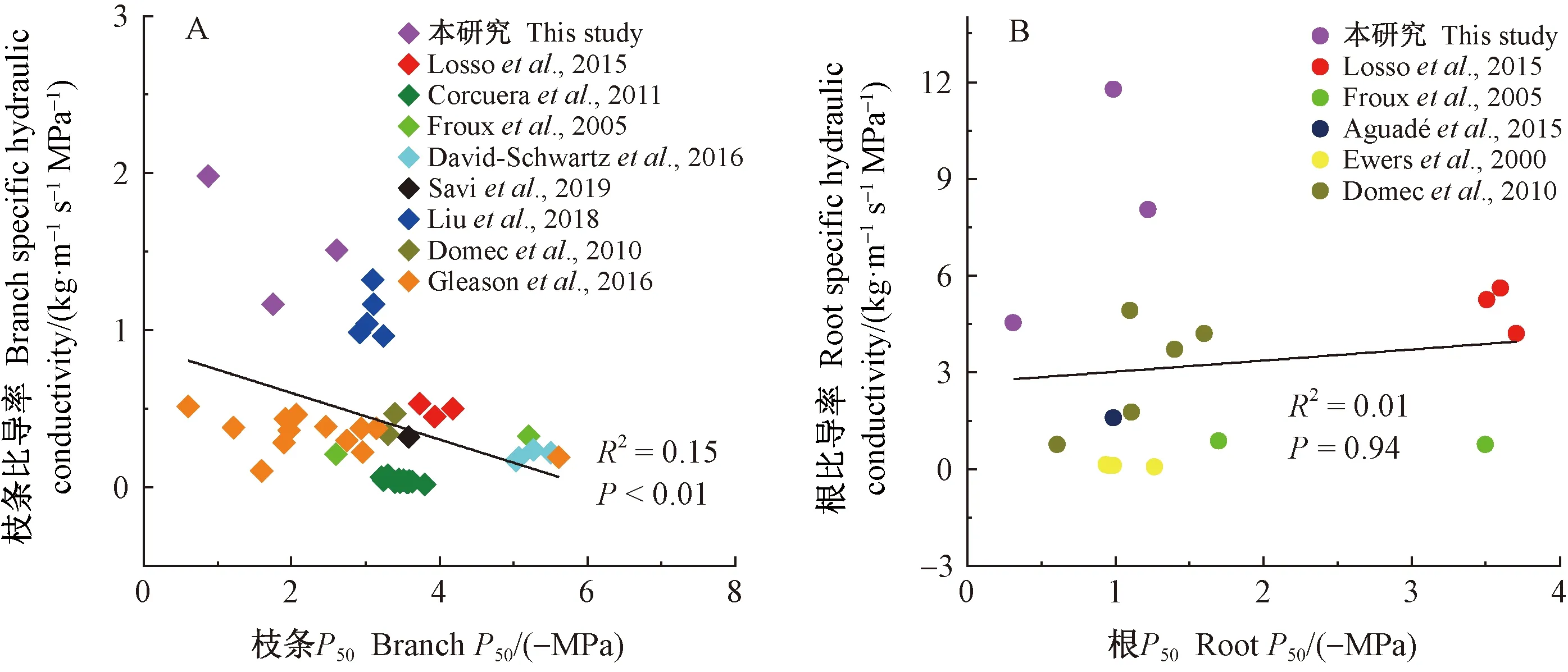
图4 松属树种枝条和根的比导率与P50的相关关系Fig.4 The correlation between branch and root specific hydraulic conductivity and P50 in Pinus species图中“本研究”是指数据点为自测所得,注明文献数据点来自对应文献。In the figure,the data points of “this study”are obtained by self-measurement,and the other data points are from corresponding literature.
2.2 木质部解剖结构分析 1)水力直径 3个树种根水力直径(Dh)极显著大于枝条Dh(图5)。各树种枝条Dh无显著差异,大小顺序为湿地松>马尾松>日本五针松;各树种根Dh约为枝条的2倍且亦无显著差异,大小顺序为马尾松>湿地松>日本五针松(图6A)。

图5 3个树种枝条和根的木质部横切面(标尺:100 μm)Fig.5 Cross sections of branches and roots for the three Pinus species(Scale:100 μm)
2)管胞密度 3个树种枝条管胞密度(N)极显著大于根的N(图5)。各树种枝条N无显著差异,大小顺序为日本五针松>湿地松>马尾松;各树种根N也无显著差异,大小顺序为马尾松>湿地松>日本五针松(图6B)。
3)单管胞壁厚度 3个树种枝条单管胞壁厚度(T)之间无显著差异,大小顺序为日本五针松>马尾松>湿地松;各树种根T之间也无显著差异,大小顺序为湿地松>马尾松>日本五针松(图6C)。

图6 3个树种枝条和根的木质部结构性状Fig.6 The xylem structural traits of branches and roots for the three Pinus species不同大写字母表示各树种枝条木质部结构差异显著(P <0.05),不同小写字母表示各树种根木质部结构差异显著(P <0.05)。下同。Different capital letters indicate significant differences of xylem structure in branches among species(P <0.05),and different lowercase letters indicate the significant differences of xylem structure in roots among species(P <0.05).The same below.
4)木材密度 各树种枝条木材密度(WD)大小无显著差异,大小顺序为日本五针松>湿地松>马尾松;各树种根WD大小为湿地松>马尾松>日本五针松,其中湿地松和马尾松根WD显著大于日本五针松(图6D)。
5)纹孔特征 3个树种中日本五针松枝条纹孔塞面积(Ato)显著大于马尾松和湿地松;马尾松根Ato在3个树种中最大,但与日本五针松和湿地松根Ato差异不显著(图7、图8A)。3个树种中日本五针松枝条纹孔膜面积(Apm)显著大于马尾松和湿地松;马尾松和日本五针松根Apm显著大于湿地松(图7、图8B)。3个树种中日本五针松枝条塞缘面积(Am)显著大于马尾松和湿地松;马尾松根Am在3个树种中最大,但与日本五针松和湿地松根Am无显著差异(图7、图8C)。

图8 3个树种枝条和根的木质部纹孔结构比较Fig.8 Comparison of xylem pit structure in branches and roots of the three Pinus species
2.3 不同器官木质部解剖结构与水力功能性状的关系 枝条水力功能性状与枝条木质部解剖结构的相关分析(图9A)发现:枝条输水效率Kss与枝条P50之间的结构需求总体上呈对立关系,即Kss与水力直径Dh负相关,而枝条P50与Dh正相关;Kss与管胞密度N、单管胞壁厚度T、纹孔塞面积Ato、纹孔膜面积Apm、塞缘面积Am正相关,而枝条P50正好相反,即上述结构指标均与枝条P50负相关。所有结构指标中:枝条Am与Kss相关性最显著,枝条Ato与枝条P50相关性最显著。
此外,以枝条水力功能性状为响应变量,木质部解剖结构为解释变量做冗余分析(图10)发现:解剖结构对Kss解释度从大到小依次为Apm(解释度32.50%,P=0.06)>Am(解释度27.30%,P=0.17)>N(解释度22.30%,P=0.20)>WD(解释度13.70%,P=0.12);Ato对枝条P50解释度最大,为85.80%(P=0.04),其次是Apm。
根水力功能性状与根木质部解剖结构的相关性分析(图9B)表明:输水效率Ksr与根P50之间的结构需求总体一致,即N、WD与Ksr、根P50均是负相关关系,T、Apm、Am与Ksr、根P50均正相关。所有结构指标中:根WD与Ksr相关性最显著,根Am与根P50相关性最显著。
冗余分析结果(图10)表明:根解剖结构对Ksr解释度从大到小依次为 WD(解释度62.00%,P=0.18)>Am>T;对根P50解释度最大的是Am(解释度50.70%,P=0.15),其次是WD(解释度26.10%,P=0.22)。
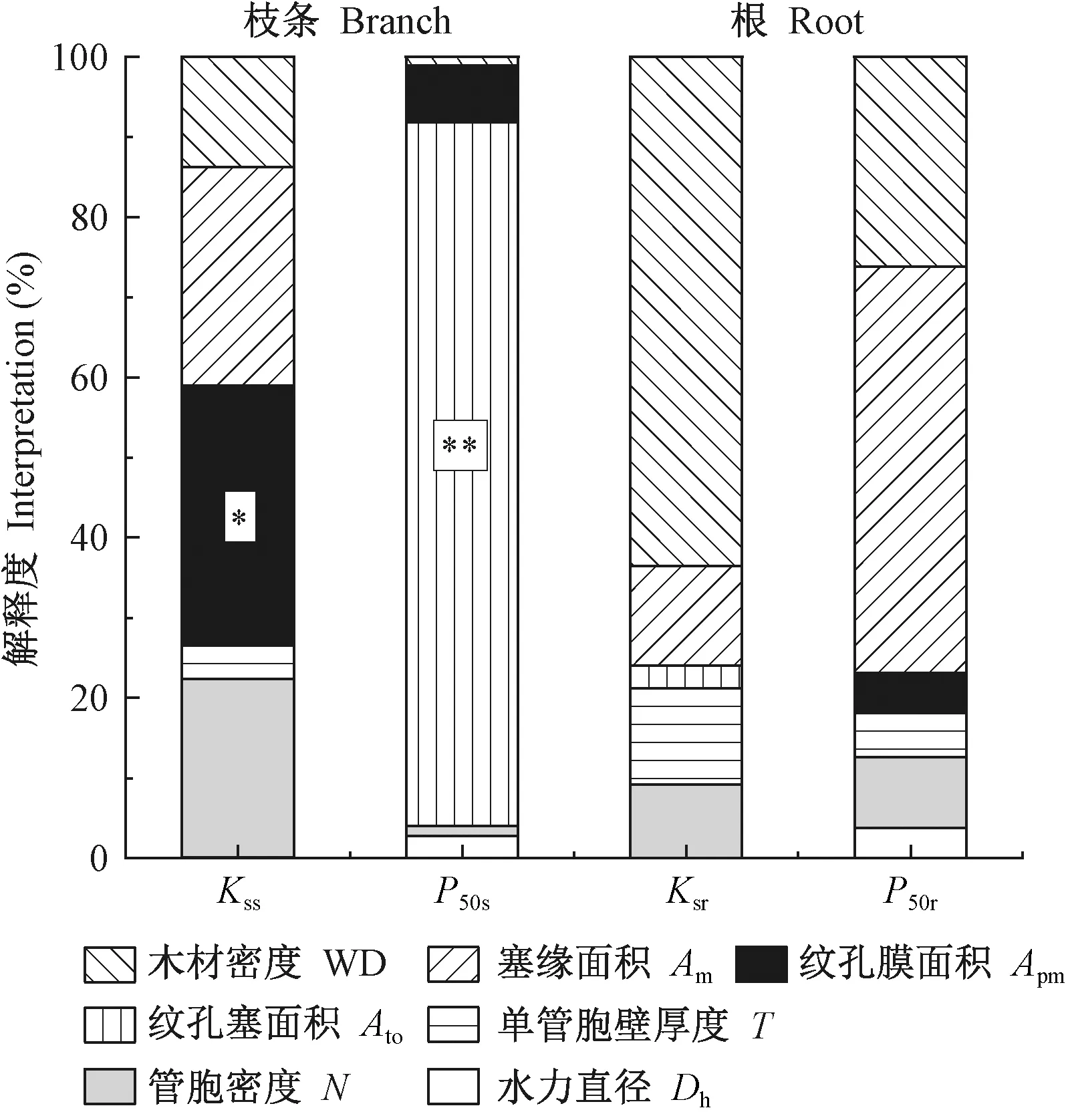
图10 枝条和根的木质部结构性状和功能性状冗余分析的解释度Fig.10 The interpretation of structural traits on functional traits by redundancy analysis(RDA)
3 讨论
3.1 地上、地下器官间效率和安全性比较 本研究涉及树种的根均比枝条更易发生栓塞(图1)。首先,根水力直径大、管胞密度小(图5,图6A、B)的特点使得根比枝条对干旱更敏感(Frouxetal.,2005)。其次,研究区水分相对充足,根较少遇到干旱,由于缺失驯化和适应,因而一旦遭遇干旱就会更脆弱,这一结果与Cuneo等(2016)的结论一致。更早的研究也表明,在针叶树种中,根比下游器官更易栓塞(Sperryetal.,1994)。一项针对全球木本植物死亡原因的研究表明,旱生树种比湿生树种更能抵抗栓塞(Choatetal.,2012),随后Zhang等(2016)发现干旱条件下根能比枝条存活更久。在干旱地区,植物受干旱胁迫时会选择牺牲地上部分(茎、叶)(Rodriguez-Dominguezetal.,2018),将非结构性碳水化合物转移到根系(Zhangetal.,2016),以维持地下部分(根)水力功能的完整性,保证其在水分状况改善后再萌发。而本研究显示,湿润区松属树种的根栓塞抗性弱于枝条。因此,干旱侵袭发生时,根更易因水力失效而死亡;这种死亡将导致植物无法再次萌发,因而更具毁灭性(Zhangetal.,2016)。
此外,Kss与Ksr、枝P50与根P50的正相关关系(图2)表明植物体内部输水效率和栓塞抗性的彼此协调:根输水效率越高的树种,其枝条输水效率也越高;根栓塞抗性强的树种,其枝条栓塞抗性也强。这与近期研究结果(Rodriguez-Dominguezetal.,2018)一致。Gleason等(2016)的研究,将输水效率区间在0~2 kg·m-1s-1MPa-1、栓塞抗性区间在-6~0 MPa的裸子植物定义为低效率低安全性。本文3个树种枝条木质部水力效率和水力安全值总体处于这一区间内,属低效率低安全性物种。
3.2 器官内效率-安全权衡 松属植物枝条木质部输水效率和栓塞抗性之间存在效率-安全权衡(图4A)。木质部解剖结构数据表明,枝条Kss和P50的结构需求趋异(图9A),这是枝条木质部效率-安全权衡关系存在的结构基础。但根木质部无效率-安全权衡(图4B),根Ksr与P50对结构需求趋同(图9B)。也就是说,结构需求趋异或趋同是效率-安全权衡存在与否的解剖结构基础。然而,效率-安全权衡不仅与木质部结构有关,也受其生境影响:根系的生境有效地保护根,这是因为在同一植物体内从根到叶水势逐渐降低,因此根系受到水分胁迫的程度最低(Tyreeetal.,2002),枝条和叶片受到的水分胁迫程度最高。这一观点也与本研究结论——根无权衡、枝条有权衡相契合:根系所处环境水分状况好,不需要通过权衡来保障自身水力安全,只需获取足够的水分满足植物体正常需求即可;反之,枝条受水分胁迫程度高,需要在保证安全的同时输送足够的水分供给叶片进行光合作用。
Cochard(1992)的研究表明,在水分供应充足情况下,7种针叶树枝条不存在效率-安全权衡。也有证据表明,不论在干旱或湿润环境下,花旗松(Pseudotsugamenziesii)枝条和根均不存在效率-安全权衡(Kavanaghetal.,1999)。上述研究及本研究结果都表明,木质部效率-安全权衡仍存在争议。源于不同生境或属于不同分类单元(被子植物、裸子植物甚至不同科属)的植物,效率-安全权衡很可能具有物种(或更大分类单元)特异性。
3.3 导水系统结构与功能相关关系的复杂性 木质部内部结构不是简单的导管或管胞的叠加,而是由无数个导管或管胞以及内部的纹孔和穿孔板相互连通所构成的三维拓扑结构(张红霞等,2017)。这一结构使木质部具有系统的属性和网络连通性,进而影响整个维管植物的输水能力(Loepfeetal.,2007)。大量的解剖结构研究已经阐明了木质部栓塞抗性与输水效率之间的关系,但管胞大小和据此计算的参数只是对输水效率和栓塞抗性的粗略估计,因为与栓塞抗性相关性最强的是管胞壁上的纹孔结构(Hackeetal.,2004)。裸子植物管胞中的具缘纹孔是影响其输水效率和栓塞抗性的重要结构指标(Boucheetal.,2014)。本研究发现:枝条纹孔膜面积(Apm)、塞缘面积(Am)对枝条比导率(Kss)的解释度最高;枝条纹孔塞面积(Ato)、Apm对枝条P50解释度最高;根Am对根比导率(Ksr)和P50解释度均突出(图10)。这表明木质部管胞间的纹孔结构不仅是决定栓塞抗性的关键结构基础,也是影响植物木质部水力导度的关键要素(Brodersenetal.,2014)。除纹孔结构外,木材密度(WD)、单管胞壁厚度(T)和管胞密度(N)也是对植物输水效率有重要影响的结构特征(图9、图10),这些指标又与木质部机械强度紧密相关,在植物水分运输过程中充当了重要角色(Lensetal.,2011;Rosasetal.,2019)。这是因为水力效率由木质部中“最脆弱”的那部分决定,增强径向抗压强度可防止管道内爆,有利于水力传输(Rosneretal.,2011)。
4 结论
本研究表明,松属树种器官间输水效率和栓塞抗性是协调变化的,枝条输水效率高,根输水效率也高;枝条栓塞抗性低,根栓塞抗性也低。马尾松、日本五针松和湿地松的枝和根的木质部水力系统效率-安全关系表现为:枝条有效率-安全权衡,根无权衡。木质部结构和功能相关性网络分析发现,结构需求趋异是枝条存在效率-安全权衡的基础,根结构需求趋同则无效率-安全权衡。植物水力系统对水分胁迫的反应是具有综合反馈机制的调节系统,因此,未来应对木质部维管网络作进一步研究,从而更深层地探究植物木质部功能与结构关系。解剖结构在器官间的差异也为根栓塞抗性低于枝条提供了解释。栓塞抗性更脆弱的根可能代表了干旱期间植物水力通道的脆弱环节,在气候变化导致干旱加剧的背景下,需要做更多的工作来深入理解根的水力功能,以提高植株水平上对植物耐旱能力的理解。
