湖北通城繁殖季节鸟类多样性年际动态及生境差异
2021-09-06杨晓菁伊剑锋楚禄建汪海兵刘彬帅李继红
张 菁 杨晓菁伊剑锋 楚禄建 汪海兵 刘彬帅李继红
(1.中国地质大学(武汉)环境学院 武汉 430078;2.生态环境部南京环境科学研究所 南京 210042)
物种多样性是生物多样性的重要组成,其变化能较准确地反映自然环境的变化。鸟类因其分布范围广,对栖息地的变化较为敏感且易于观察,常作为生态环境监测的指示物种[1-2]。对一个地区进行长期的鸟类多样性监测,有助于切实地评价该地区自然生态环境质量的优劣,并进一步了解当地自然生态环境的变化及其所面临的威胁,以期为环境保护政策的制定和自然保护区的建设和管理提供科学依据。
通城县位于湖北省东部最南缘,地处幕阜山脉北麓的湘鄂赣三省交界处,地理位置较为特殊。在中国动物地理区系划分和鸟类区系分布上,通城县属于东洋界的华中区[3],位于东亚—澳大利西亚候鸟迁徙区西侧[4],境内有大溪国家湿地公园和药姑山省级自然保护区。目前有关通城县鸟类多样性现状的研究报道较为缺乏,仅有邻近的咸宁城市春季鸟类群落调查报道[5]。本研究于2016~2019每年繁殖季节对通城县鸟类群落进行系统调查,以掌握该区域繁殖季鸟类群落组成及其多样性特征;并通过对当地不同生境鸟类多样性及其年际动态进行比较分析,评估通城县生态环境质量及其受胁因素。
1 研究方法
1.1 研究区域概况
湖北省通城县地处幕阜山脉北麓(113°36′~114°4′E,29°2′~29°24′N),总面积1 140.7 km2。海拔跨度为78~1 528.30 m,一般海拔在1 000 m以下。地形为丘陵起伏的盆地,东、南、西被群山环绕。通城县属于亚热带湿润季风气候,四季分明,雨量充沛,县内水资源丰富,河流、水库、湖泊众多。通城县植被繁茂,植物组成丰富,以壳斗科Fagaceae、樟科Lauraceae、冬青科Aquiforliaceae等和竹亚科Bambusoideae、亚热带松柏类植物等为主组成常绿阔叶林、常绿落叶阔叶混交林、亚热带竹林和针叶林等植被类型[6]。因人类活动影响,通城县的自然植被仅在高海拔区域及保护区内局部残存少量原始次生林,调查区域内大部分现生植被是以常绿阔叶林为主的、混有人工杉木、马尾松林的半自然林或人工林[6-7]。该区域还广泛种植毛竹Phyllostachysedulis,竹林与乔木林呈斑块状混杂,是当地植被的重要组成部分。
1.2 调查方法
分别于2016~2019每年4~7月对通城县繁殖季鸟类群落进行调查,每年调查两次,两次调查间隔至少30 d。调查采用样线法,样线设置遵循有效性、代表性、可行性和固定性的选择原则[8],考虑不同海拔、生境、人为活动干扰类型、交通条件等因素,在通城县域内共设置样线10条,涵盖了大溪国家湿地公园、药姑山省级自然保护区、通城县城及周边村镇等各区域(图1)。样线长度1.0~3.0 km,总长18.5 km。主要涉及生境有乔木林、竹林、乡村和内陆水体4大类。因当地竹林生境显著,比重大,故将其与乔木林生境并列。乔木林下灌草丛并入乔木林统计。乡村生境中除村舍建筑物外,间杂旱田、水田、树丛、沟渠等多种小生境斑块。内陆水体包括湖塘、水库、河流、溪流等。每条样线出现的生境类型1~3种,调查时记录样线上各类生境所占比例。
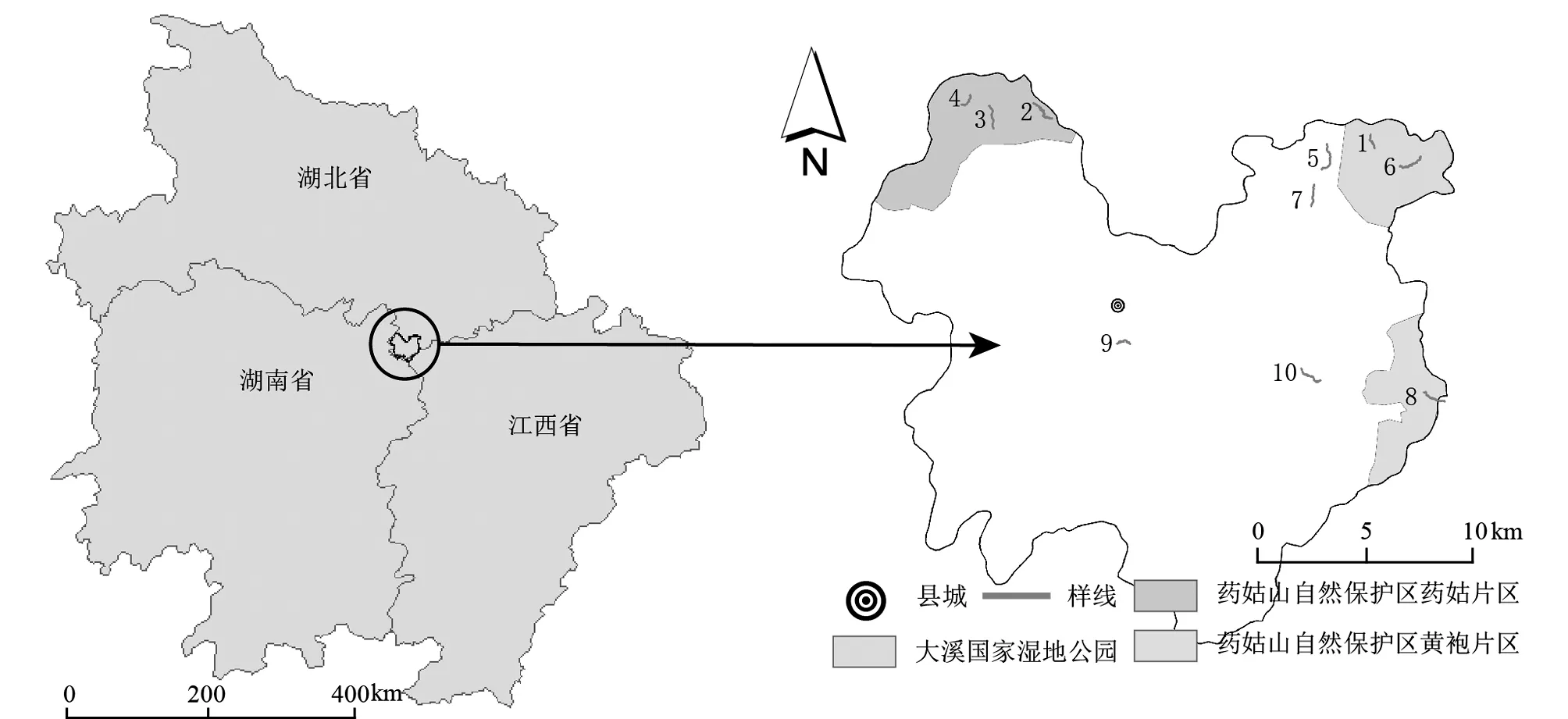
图1 湖北省通城县地理位置及样线分布图(编号1~10为样线编号)
选择晴好天气,在鸟类活动高峰期的早上(05∶30~09∶30)和下午(16∶00~19∶00),以1.5~3.0 km·h-1的步行速度按照设置好的固定样线进行调查,记录沿线看到和听到的鸟类种类、个体数量和所处生境类型,记录范围为样线两侧各约100 m。观测设备为双筒望远镜(Olympus 10×42,Optisan EL 8×42WB),使用相机和便携式数码录音机记录鸟类外形、生境及鸣声辅助鸟类识别。
1.3 数据分析与统计
调查名录中鸟类分类及居留型的确定根据《中国鸟类分布与分类名录(第三版)》[9],物种鉴别参考《中国野外鸟类手册》[10],濒危物种等级参考《国家重点保护野生动物名录(2021)》[11]。每次调查后统计的群落特征参数如下:
(1)用重要值(I.V.)评价每种鸟的优势度[12-13]。I.V.=(相对空间成分+相对数量成分)/2×100,其中,相对空间成分=某种鸟出现的样线数/总调查样线数;相对数量成分=某种鸟个体数/最多的那种鸟的个体数。因每次调查持续天数较短,故公式中不计入相对时间成分。50≤I.V.<100的为最重要的鸟类,即每次调查的优势种。
(2)计算总调查区域和各生境的鸟类密度(D)及单位面积物种数(R)。D=N/2LW,N为样线内记录的鸟类个体数量,L为样线长度,W为样线单侧宽度,按100 m计算。R=S/A,S为物种数,A为样线面积。
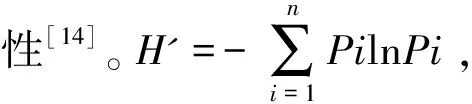
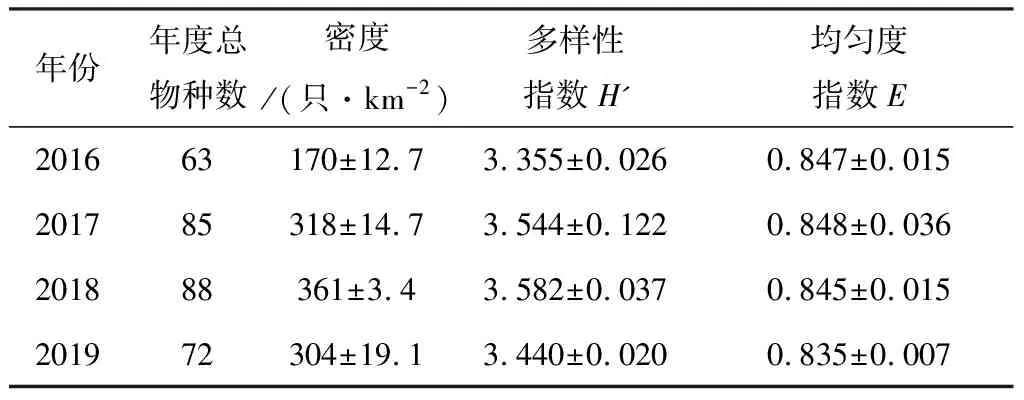
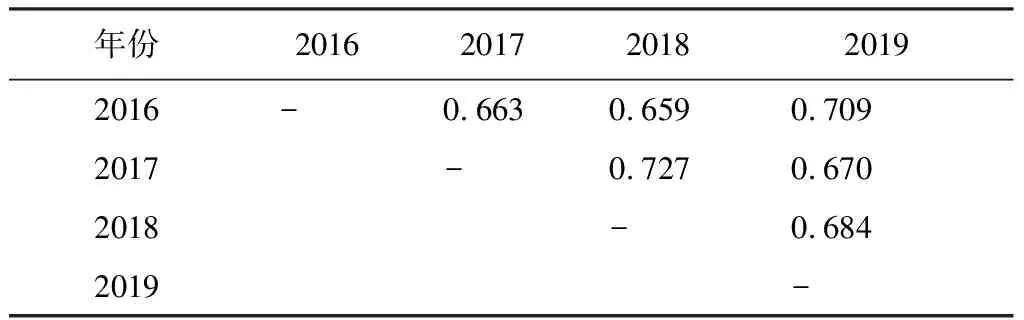
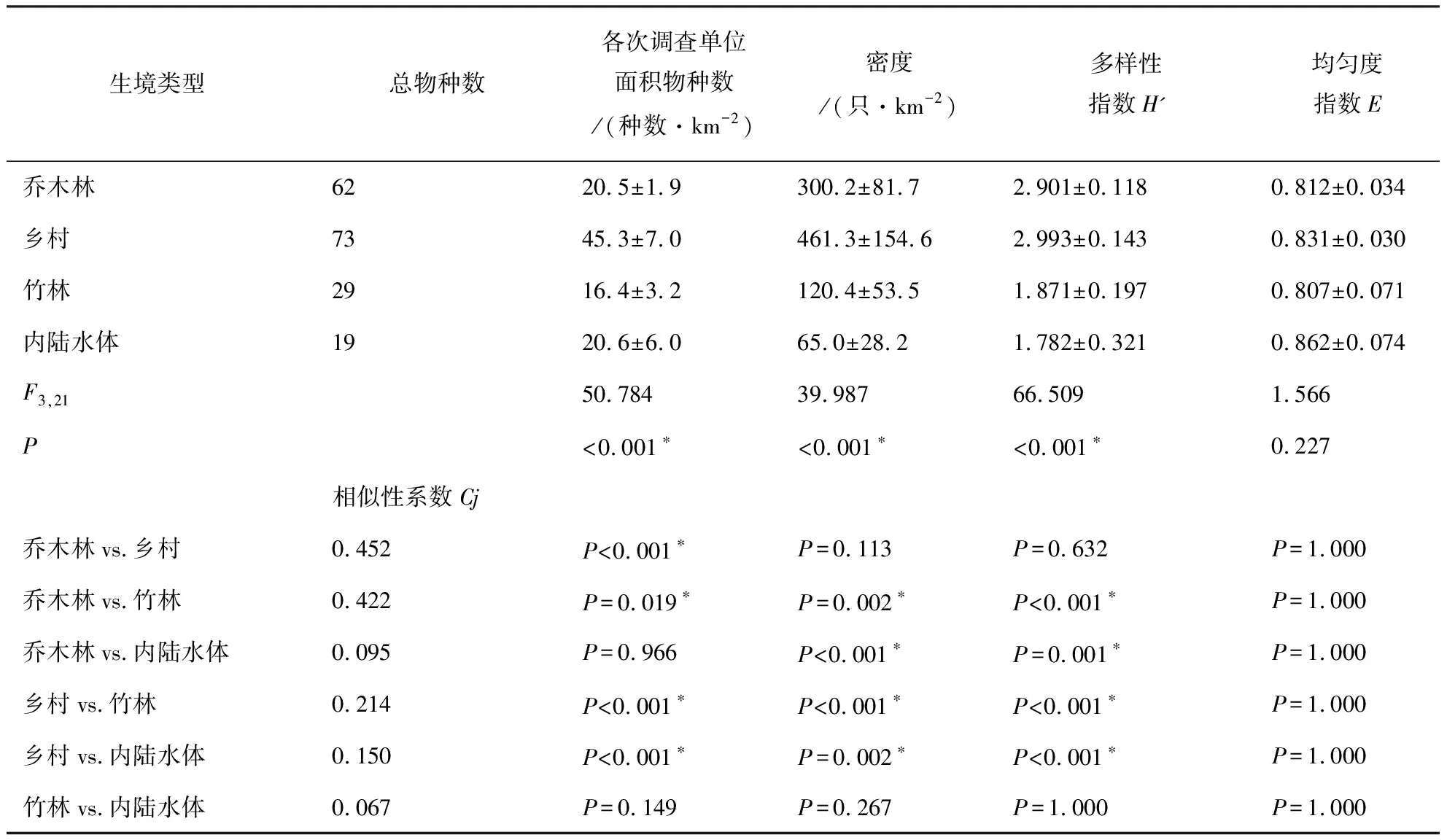
(4)采用Jaccard相似性系数(Cj)对历年以及各生境的群落物种组成进行相似性分析。Cj=j/a+b-j,其中a、b为两个群落分别的物种数,j是两个群落共有的物种数。0.75 年度间比较时,先检验每年内两次调查的数据是否存在显著差异,采用非参数的Wilcoxon符号秩检验(Wilcoxon Signed Ranks test)对2个配对样本进行检验,若无显著差异,则取年内两次调查的数据均值进行年度间比较。进行生境间比较时,采用各生境的4年8次调查数据进行单因素重复测量方差分析(One-way repeated measures ANOVA)。所有的统计分析在SPSS 16.0软件中进行。显著性差异水平设为P<0.05。 2016~2019年4年8次繁殖季节调查共记录到鸟类105种8 626只次,隶属于13目40科。其中雀形目Passeriformes种类最多,共计26科70种,占总种数的66.7%,其次为鹃形目Cuculiformes,有1科6种,占总种数的6.17%(见附录)。雀形目中又以鹟科Muscicapidae鸟类最多,共有10种,其次为鹎科Pycnonotidae,有6种。 共记录到国家II级重点保护鸟类15种,分别为白鹇Lophuranycthemera、小鸦鹃Centropusbengalensis、蛇雕Spilornischeela、白腹隼雕Aquilafasciata、赤腹鹰Accipitersoloensis、凤头鹰A.trivirgatus、灰脸鵟鹰Butasturindicus、斑头鸺鹠Glaucidiumcuculoides、领鸺鹠G.brodiei、蓝喉蜂虎Meropsviridis、白胸翡翠Halcyonsmyrnensis、红隼Falcotinnunculus、画眉Garrulaxcanorus、红嘴相思鸟Leiothrixlutea、白喉林鹟Cyornisbrunneatus。此外,还记录到湖北省新分布纪录2种——丽星鹩鹛Elachuraformosa[16]和岩燕Ptyonoprognerupestris、湖北省新分布亚种1种——灰眶雀鹛东南亚种Alcippemorrisoniahueti。 历年调查的优势种组成和种数各不尽相同,但全部为雀形目鸟类。白头鹎Pycnonotussinensis、强脚树莺Horornisfortipes、大山雀Paruscinereus在8次调查中均为优势种。其他优势种还有画眉、棕头鸦雀Sinosuthorawebbiana、领雀嘴鹎Spizixossemitorques、暗绿绣眼鸟Zosteropsjaponicus、棕颈钩嘴鹛Pomatorhinusruficollis、棕脸鹟莺Abroscopusalbogularis等。 记录到的105种通城繁殖季节鸟类中以东洋界鸟类为主,共有58种,占总物种数的55.24%;广布种27种,占25.71%;古北界鸟类20种,占19.05%。居留型方面,留鸟是通城繁殖季节鸟类群落的主要组成部分,共记录到留鸟68种,占总物种数的64.8%;夏候鸟29种,占27.6%;旅鸟15种,占14.3%;冬候鸟11种,占10.5%(有的鸟类在该分布区不止一种居留型)。 年内比较:比较2016~2019年每年内相隔一月的两次调查鸟类物种数(Wilcoxon符号秩检验:Z=-1.890,P=0.125>0.05)、密度(Z=-1.826,P=0.125>0.05)、多样性指数(Z=-1.461,P=0.250>0.05)和均匀度指数(Z=-1.826,P=0.125>0.05),均无显著性差异。2016~2019各年度内两次调查的物种相似性系数分别为0.698、0.624、0.602和0.708,均处于0.5 年际间比较:由表1可知,2016年记录的总物种数、密度和群落多样性指数均最低,2018年的最高。均匀度指数则是2019年最低,2017年最高。中间两年的群落多样性略高于头尾两年,但4年的群落多样性指数和均匀度指数实际波动并不剧烈,多样性指数在3.3~3.6范围内波动,均匀度指数则在0.83~0.85范围内波动(表1)。不同年份间的鸟类群落相似性系数均处于0.5 表1 2016~2019年通城繁殖季节鸟类群落特征(均值±标准差,n=2次调查) 表2 2016~2019不同年份间通城繁殖季节鸟类群落相似性系数 统计并比较通城4大类生境(乔木林、竹林、乡村和内陆水体)中的各群落特征参数,结果见表3。各生境中,乡村生境的总物种数、单位面积物种数、密度和多样性指数都最高;乔木林生境的总物种数、密度和多样性指数均仅次于乡村生境,但其单位面积物种数低于内陆水体生境,排在第三;竹林生境的单位面积物种数最低。密度和多样性指数最低的都是内陆水体生境。均匀度指数则是水体最高,竹林最低。 除均匀度指数外,4类生境的单位面积物种数、密度、多样性指数均存在显著性差异(P<0.001)。两两比较结果显示,乡村生境的单位面积物种数显著高于其他三类生境(P<0.001),乔木林和竹林间也存在显著差异(P=0.019<0.05),其余则不存在显著差异(P>0.05)。密度和多样性指数是乔木林与乡村都显著高于竹林和内陆水体(P<0.05),但两个参数在乔木林与乡村之间、以及竹林与水体之间都不存在显著差异(P>0.05)。均匀度指数在各生境间均不存在显著差异(P=0.227>0.05)(详见表3)。 乔木林与乡村、乔木林与竹林的相似性系数属于0.25 表3 通城不同生境繁殖季节鸟类群落特征(均值±标准差,n=8次调查)、单因素重复测量方差分析结果及不同生境群落相似性系数 通城繁殖季节鸟类群落区系组成以东洋界鸟类为主,这与通城县在中国地理区系划分中属于东洋界华中区密不可分[3]。此外,通城县所处区域位于东亚—澳大利西亚候鸟迁徙区西侧[4],春季迁徙季节有大量鸟类途经该区域北迁,这是在该区域记录到19.05%古北界鸟类的主要原因。通城县繁殖季节鸟类居留型组成以留鸟为主,占总调查种数的64.7%,其次为夏候鸟(27.6%)。该结果与咸宁市市区(留鸟53.4%,夏候鸟26.0%)[5]、安徽鹞落坪(留鸟60.8%,夏候鸟30.4%)[17]、南京紫金山(留鸟61%,夏候鸟28%)[15]、河南杞县、淮阳县黄淮平原(留鸟>60%)[18]等地区的繁殖季节鸟类群落构成类似。而纬度较高的地区,如甘肃碌曲(夏季,留鸟44.8%,夏候鸟40.8%)[19]、辽宁省(夏季,留鸟26.4%,夏候鸟45.5%)[20]、新疆塔城(留鸟28.7%,夏候鸟71.3%)[21]的繁殖季鸟类中夏候鸟比例明显增加。纬度相对更低的广西大明山(留鸟81.7%,夏候鸟18.3%)繁殖季节留鸟占比较通城更高[22]。总体呈现出随纬度越高,当地繁殖季节鸟类群落中留鸟比例越低,夏候鸟比例越高的趋势[23]。 每年年度内两次调查的鸟类群落相似性分析显示均为中等相似群落。两次调查相隔至少30 d,因此物种组成不完全一样。部分旅鸟或冬候鸟如鹀科鸟类、黄眉柳莺Phylloscopusinornatus、红尾歌鸲Larvivorasibilans、白腰草鹬Tringaochropus等仅见于第一次调查(4月底),到达较晚的蓝喉蜂虎等夏候鸟仅见于第二次调查(5月底、6月初)。当然也不排除有观察的偶然性等因素。 比较2016~2019年的通城繁殖季节鸟类群落特征,2016年鸟类密度明显较低的原因不排除首年调查记录方法的不完善。2019年多样性较前两年略有下降,有可能与当年通城大溪国家湿地公园内公路整修拓宽、内冲瑶族村旅游开发项目施工相关,部分调查样线受施工干扰较严重。尽管如此,4年的群落多样性指数和均匀度指数实际波动却并不剧烈。此外,相似性分析结果显示通城繁殖季节鸟类群落组成在2016~2019各年度间均属于中等相似,说明这期间鸟类群落组成基本稳定。 调查区域内各生境的群落组成相似度均不高,为中等不相似至极不相似,说明了各类生境的独特性,不同生境对于当地鸟类群落的构成和多样性的维持都有其不可替代的作用。乡村生境的鸟类总物种数、单位面积物种数、密度、多样性指数均高于其他生境,尤其单位面积物种数显著最高,说明乡村生境鸟类是通城繁殖季节鸟类群落的重要组成部分。生境内景观的异质性和层次丰富性是影响物种多样性的重要因素[24]。包含有房屋、树丛、灌丛、农田和沟渠的传统乡村生境是调查区域内层次最丰富、景观异质性最高的生境类型,其间各种小生境斑块交错存在。尽管乡村生境存在一定的人为活动干扰,但该生境仍具有最为丰富的鸟类种类、数量及最高的生物多样性。一些伴人物种如八哥Acridotherescristatellus、乌鸫Turdusmandarinus、鹊鸲Copsychussaularis、金腰燕Cecropisdaurica等只见于居民点附近。池鹭Ardeolabacchus、灰头麦鸡Vanelluscinereus等湿地鸟类也依赖乡村里的水田进行觅食,因此乡村能为更多的对生境有不同需求的鸟类提供合适的栖息、觅食和繁殖场所。此外,居民点和农田等开阔生境中的鸟类也较森林中更容易被观察到,且不少鸟种还常在这类生境中集群分布。故而尽管调查的乔木林生境所占比例最大,几乎是乡村生境面积的2倍,但在通城这样人类活动频繁、植被以半自然林为主的区域,乡村生境往往能比山区林地记录到更多的鸟类种类和数量。当然,也有不少物种,特别是一些非常见鸟类,如白鹇、白喉林鹟、丽星鹩鹛等仅见于人类干扰较少的自然林地。 类似情况也出现在其他地区,在与通城县临近的咸宁城市春季鸟类群落中,农田生境因分布于丘陵与乡村建筑群之间,具有边缘效应,加上农作物可为鸟类提供丰富的食物,相比于当地的常绿阔叶林、常绿落叶阔叶混交林、竹林等生境,农田生境具有最高的鸟类密度与多样性[3]。江西武夷山-黄岗山西北坡的农林营作干扰区生境同样也是该调查区域内景观多样性和微环境结构最复杂的生境类型,相较于林区和灌丛草甸区生境具有相对较高的G-F多样性指数[25]。卢训令等认为人类活动(村庄和农田耕作等)不一定对鸟类种群和生态系统服务呈现负效应。农业景观对于生物多样性的保护具有举足轻重的作用,它为众多生物(包括鸟类)提供了食物、栖息地和庇护所,尤其是农林复合系统常能吸引更多的鸟类[18,26]。因此,间杂的人类村舍和树林、灌丛、旱田、水田、池塘、沟渠等不同生境类型形成良好的交错平衡能保证丰富的鸟类多样性。保持这样具有高度空间异质性的乡村生境对维持整个区域的鸟类群落多样性,乃至其它类群的生物多样性都具有非常重要的意义。 调查区域中的竹林生境和内陆水体生境所占比例较小,且竹林生境相对于其他生境组成较单一,结构简单,因此其鸟类群落构成也相对简单,数量和多样性较低,主要由一些喜活动于竹林生境的林鸟组成,与乔木林生境有一定重叠。当地常见于竹林的鸟类有棕脸鹟莺、暗绿绣眼鸟、红嘴相思鸟等。调查区域内的水体生境主要包括水库、池塘、河流、溪流等,繁殖季节常见鸟类为鹭、、鹡鸰、翠鸟、水鸲、燕尾等水禽或傍水鸟类,不见或少见于陆地生境,因此与其他生境的相似性最低。 本研究对通过4年8次调查数据的统计分析,系统掌握了通城县范围内的繁殖季节鸟类多样性特征及动态。总体上看,近4年来通城县繁殖季节鸟类群落组成基本稳定,生境尚未发生显著变化。空间异质性较高的乡村生境对当地鸟类多样性贡献较大。然而,一旦人类干扰程度加剧,传统乡村中人为环境和自然环境的比例与分布失衡,将会导致多样性的丧失。目前的通城鸟类群落所面临的主要威胁是大规模的开发建设。2019年调查中即发现当地的乡村旅游开发和道路建设施工对样线上的鸟类多样性有负面影响。随着施工结束,人为干扰应会相应减弱,但开发后成为旅游景区的乡村势必较原来干扰程度加剧。建设美丽乡村,坚持生态优先,在项目规划建设时,要尽量避免或减少对于包括传统乡村生境在内的鸟类栖息地的干扰、破坏和违规占用。 致谢:感谢湖北药姑山省级自然保护区管理局在2016年为本研究调查方案的制定及野外工作中所提供的帮助。感谢参加过野外调查工作的张鹏程、张天、汪文韬、兰希平、张运晨、田尚坤、禹墨涵同学。感谢通城县当地所有为本研究野外调查提供过帮助的社会人士。2 结果
2.1 物种组成
2.2 年内及年际间比较
2.3 生境间比较
3 讨论
3.1 区系、居留型及群落多样性年度分析
3.2 乡村生境对通城鸟类多样性贡献最高
3.3 受胁因素及保护建议
