近自然经营对黔中地区马尾松灌草群落多样性及物种共存的影响
2021-09-05臧丽鹏张广奇何跃军刘庆福陈丹梅
臧丽鹏 张广奇 何跃军 刘庆福 陈丹梅


摘 要:马尾松是我国低山丘陵地区造林的主要用材树种之一,近自然经营是人工林管理的一种有效途径,也是马尾松人工林生态经营的关键技术。为探究近自然经营对马尾松人工林林下物种多样性及共存的影响,研究选取中龄林间伐和近熟林择伐的近自然经营马尾松人工林为对象,通过样地调查、多样性对比及模型拟合等方法探究近自然经营对马尾松人工林林下灌木、草本群落多样性格局及潜在生态过程的影响。结果表明,近自然经营8年后,灌木、草本群落α物种多样性变化显著。灌木群落的物种组成与经营前差异明显,物种共存机制由生态位向中性过程逐渐转化;近自然经营对草本群落的物种组成和物种共存机制无显著影响,地域差异对物种组成及物种共存影响更大。
关键词:物种共存;种多度分布模型;近自然林经营;马尾松人工林
中图分类号:Q948.1
文献标识码:A
文章编号:1008-0457(2021)03-0021-08
国际DOI编码:10.15958/j.cnki.sdnyswxb.2021.03.004
Effects of Close-to-Nature Management on Biodiversityand Species Coexistence of Shrubs and Herbs in Pinus massoniana Monoculture Plantations of Central Guizhou
ZANG Lipeng,ZHANG Guangqi,HE Yuejun*,LIU Qingfu,CHEN Danmei
(Institute Center of Forest Ecology,College of Forestry,Guizhou University,Guiyang,Guizhou 550025,China)
Abstract:
Pinus massoniana is one of the most important afforestation tree species in China.Close-to-nature management is one of the most efficient theories for managing plantations,andit is also a key technique in managing P.massoniana monoculture plantations.The study conducted two dominant managements (intermediate cutting in half-mature forests and selective cutting in near-mature forests) on P.massoniana monoculture plantations.By investigating community,comparing diversity patterns and fitting models,we explored the effects of close-to-nature management on diversities and potential ecological processes of shrubs and herbs communities.Results showed that the α-diversity indexes varied significantly after 8-years close-to-nature managing.The species composition of shrub community was significantly different from that before the management.The mechanism of species coexistence of shrubs gradually changed from niche to neutral process.There was no obvious variation in species composition and mechanisms of species coexistence of herbs communities.However,it seemed that the geographic locationshad a greater impact on species composition and ecological process of herbs compared with management.
Keywords:
mechanisms of species coexistence;Species abundance distribution;Close-to-nature management;Pinus massoniana monoculture plantations
森林作為最重要的陆地生态系统,在调节气候、提供生态系统服务等方面均发挥主导作用。然而,随着人类活动的日益加剧,森林生态系统退化严重[1- 2]。有研究表明,全球森林面积呈现总体下降趋势,但人工林面积却以200~300万公顷/年的速度增加;可见,人工林在生态系统服务等方面发挥的作用日趋重要[3]。马尾松具有耐瘠薄、干旱等特性,是我国南方低山丘陵地区的主要造林树种之一。尽管大面积分布的马尾松人工林有较大的径材产出,但也存在林相单一、养分循环较差、立地退化、生态系统脆弱等缺点[4]。因此,如何对其进行科学经营从而增加系统稳定性,这已成为当下林业面临的关键问题之一。
近自然森林经营是指充分利用森林自然生长发育规律,以森林更新到演替顶级完整的森林全过程为依据规划森林经营活动,从而优化森林结构功能,达到生态与经济平衡持续产出的一种接近自然的森林经营模式[5]。该理论起源于1880年德国林学家Gayer提出的“接近自然的林业理论”,在Moller为代表的自然林业学派的推广下,其理论和技术体系日趋成熟和完善。20世纪70年代以后,世界各国开始逐渐引入近自然经营理论并付诸实践。研究发现,近自然经营在森林抗干扰能力、蓄积量、林地立地质量等方面均有较为显著的成效[6]。森林生态系统稳定性很大程度依赖于群落结构及生物多样性格局,较高的生物多样性有助于生态系统功能稳定性及多功能性发挥[7-8]。因此,辨别近自然经营对生物多样性格局的影响是评估生态系统稳定性的有效途径。贵州省森林资源丰富,主要的森林植被类型包括亚热带常绿阔叶林、常绿落叶阔叶混交林、常绿针叶林、竹林等,其中马尾松人工林所占比例较大,可达35%以上。为探究近自然经营对马尾松人工林生物多样性及生态系统稳定性的影响,本研究以2种近自然经营措施(中龄林间伐和近熟林择伐)的马尾松人工林为研究对象,通过样地调查,对比分析近自然经营实施8年对马尾松人工林林下灌木群落和草本群落的物种组成及多样性格局的影响,以检验近自然经营成效,以期为马尾松人工林后期经营决策与规划提供坚实有效的科学依据。
1 材料与方法
1.1 研究区概况
本研究在贵州省贵阳市开阳县和息烽县完成(110°07′~110°17′E,21°22′~26°11′N),平均海拔1200 m左右,属于典型的亚热带湿润温和型气候,水热资源丰富,年平均气温15.3℃,年均降水量为1200 mm,雨热同期。
1.2 样地设置
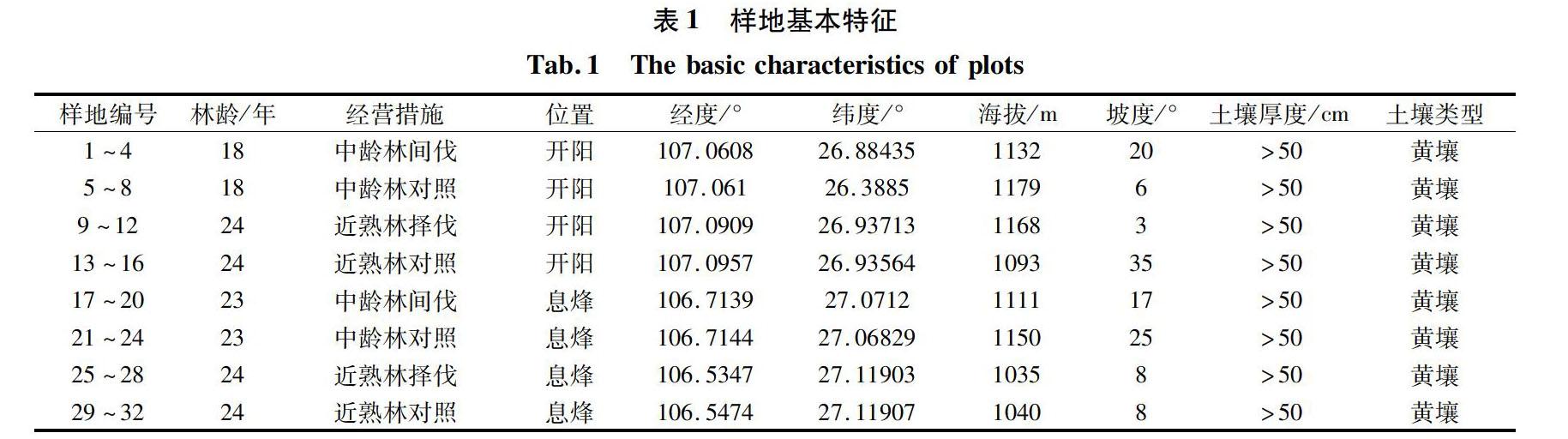
根据不同近自然经营措施,共计设置中龄林间伐样地8个,近熟林择伐样地8块,每个样地旁边设置1个对照样地,共计32个样地。其中息烽县包括中龄林间伐样地4个,近熟林择伐样地4个,及相应对照样地,共16个样地;开阳县包括中龄林间伐样地4个,近熟林择伐样地4个,及相应对照样地,共16个样地。调查样方面积均为20 m×20 m。对每个样地详细记录样地编号、地理位置、样地面积、调查日期,并测定海拔、地形、坡向、坡度、土壤类型、干扰状况、经营利用状况及生境概况等林分情况指标。在样方内开展相应的乔木、灌木、草本调查:乔木调查基于20 m×20 m样方进行,记录物种、胸径、树高、枝下高、冠幅、生长状态等信息;灌木基于5 m×5 m样方进行,记录物种、基径、盖度、数量、高度、生长状态等信息;草本基于1 m×1 m样方进行,记录物种、盖度、生长状态等信息。样地详细信息如表1所示。
1.3 物种多样性指数计算
本文主要通过Patrick指数、Simpson指数、Shannon-Weiner指数、Pielou均匀度指数等来量化林下灌木、草本的α多样性格局。各指数计算公式如下:
Patrick指数:
R=S(式1),
式中,R为物种丰富度,S为每一样方的物种数目;
Simpson指数:
D=1-∑ Pi2(式2),
式中,Pi为i种的个体数占群落中总个体数的比例;
Shannon-Weiner指数:
H=-∑S(i=1)Pi ln Pi(式3),
式中,Pi为i种的个体数占群落中总个体数的比例;
Pielou均匀度指数:
E=H/Hmax(式4),
式中,H为实际观察的物种多样性指数,Hmax为最大的物种多样性指数,Hmax=lnS,S为群落中的总物种数。
1.4 数据分析
根据近自然经营后的样地与对照将样地分为两类,采用非度量多维标度(NMDS,non-metric multidimensional scaling)方法检验近自然经营对灌木群落和草本群落物种组成的影响。模型中样地相似性距离计算参数采用Chao相似性系数,将灌木群落和草本群落分别进行NMDS分析[9]。
本文利用种多度曲线拟合来预测近自然经营下马尾松人工林林下灌木、草本的物种共存机制是否发生变化。本研究的基本流程是作出样地的实际种多度分布曲线,将其与生态位模型和中性模型进行拟合,通过参数检验拟合优度来确定不同生境类型下究竟是什么过程对群落组配起到更重要作用[10]。最常用的生态位模型是由Motomura提出的几何级数模型,它的原理是假设群落中优势度最高的物种占据总生态位的a份,第二优势的树种占据剩余生态位的a份,即总生态位的a(1-a),一直到剩余的生态位不够支撑一个物种为止,第i个物种的预期多度可由公式计算:
Ni=Na×(1-a)i-1 (i=1,2,……)。
而中性模型本文选用Volkov模型[11],模型中包含n个个体物种频度则可用以下公式表示:
δ=x(1-x),
y=μγδθ,
γ= m(J-1)1-m,
Γ(z)=∫∞0 tZ-1 e-t dt,
S(n)=θJ!n!(J-n)! Γ(γ)Γ(J+γ) ∫γ0 Γ(n+y)Γ(1+y) Γ(J+γ-n-y)Γ(γ-y) exp(-yθγ)dy。
式中,x表示由于出生、死亡、迁移等原因造成的每个物种出生率和死亡率之比,x小于1,μ是群落中第k个物种的多度,J是群落中个体数量。γ为迁移到局域群落的个体数。T是在第k个物种含有n个个体的时间。
针对每一块样地的灌木和草本的物種多度分布格局进行生态位模型和中性模型拟合,物种多度分布模拟拟合过程使用R语言中“sads”包中的fitsad功能进行验证。模型拟合优度使用卡方检验,若P<0.05,则模型被拒绝,若P>0.05,则模型被接受。使用赤池信息量准则则(Akarke Information Criterion,AIC)进行模型拟合效果评估,AIC值越小,模型拟合效果越好,如果两个模型AIC值差距小于2,则认为两个模型在统计学上没有显著差异。数据分析主要通过R3.0.2[12]中的“vegan”、“sads”等包进行计算。
2 结果与分析
2.1 近自然经营对林下灌木、草本α物种多样性指数的影响
本研究选用Patrick丰富度指数、Shannon-Weiner指数和Simpson指数表示林分群落各层的植物多样性,用Pielou均匀度指数表示群落物种的分布情况。各林分下灌木和草本的α物种多样性如表2、表3所示。
由表2可以看出,开阳中龄林间伐样地的灌木群落Patrick丰富度指数、Shannon-Weiner指数和Simpson指数均高于对照样地,但Pielou均匀度指数低于对照样地;而开阳近熟林择伐样地Patrick丰富度指数、Shannon-Weiner指数低于对照样地,Simpson指数、Pielou均匀度指数高于对照样地,表明不同经营措施下对于开阳间伐样地的马尾松人工林下的灌木群落物种α多样性产生不一样的影响效应,表现为中龄林间伐增加了物种丰富程度,但是降低了物种的均匀程度;而近熟林择伐则减少了物种丰富程度,增加了物种分布的均匀度。
息烽中龄林间伐样地的灌木群落Patrick丰富度指数和Shannon-Weiner指数低于对照样地,Simpson指数和Pielou均匀度指数与对照样地无显著差异;而息烽近熟林择伐样地Patrick丰富度指数、Shannon-Weiner指数、Simpson指数、Pielou均匀度指数均显著高于对照样地。
由表3可以看出,开阳中龄林间伐样地和近熟林择伐样地的草本群落Patrick丰富度指数、Shannon-Weiner指数、Simpson指数和Pielou均匀度指数均低于对照样地。息烽中龄林间伐样地的草本群落Patrick丰富度指数显著高于对照样地,而Pielou均匀度指数低于对照样地,Shannon-weiner指数和Simpson指数无显著差异;而息烽近熟林择伐样地Patrick丰富度指数、Shannon-Weiner指数、Simpson指数、Pielou均匀度指数均显著低于对照样地。
2.2 近自然经营对林下灌木、草本物种组成的影响
本研究采用NMDS排序表征群落间物种组成相似性。由图1可以看出,近自然经营样地和对照样地的灌木群落物种组成差异显著,而草本群落物种组成差异不显著。
2.3 近自然经营对物种多度模型拟合效果的影响
通过物种多度模型拟合不同经营方式对林下灌木、草本的影响结果如图2、图3所示。
由图2和表4来看,开阳中龄林间伐样地内灌木群落表现为中性模型拟合效果更好,而对照样地也表现为中性模型拟合效果更好;开阳近熟林择伐样地内灌木群落表现为中性模型拟合效果更好,对照样地表现为生态位模型拟合效果更优。息烽中龄林间伐样地内灌木群落表现为中性模型拟合效果更好,而对照样地表现为生态位模型拟合效果更好;开阳近熟林择伐样地内灌木群落表现为中性模型拟合效果更好,对照样地表现为生态位模型拟合效果更优。
由图3和表4来看,开阳中龄林间伐样地内草本群落表现为中性模型拟合效果更好,而对照样地也表现为生态位模型拟合效果更好;开阳近熟林择伐样地内草本群落表现为生态位模型拟合效果更好,对照样地也表现为生态位模型拟合效果更优。息烽中龄林间伐样地内草本群落表现为中性模型拟合效果更好,而对照样地也表现为中性模型拟合效果更好;开阳近熟林择伐样地内草本群落表现为中性模型拟合效果更好,对照样地表现为中性模型拟合效果更优。
3 结论与讨论
近自然经营可改善人工林养分循环,增加生态系统稳定性,实现生态与持续产出的平衡[5]。生物多样性是生态系统功能稳定性和多功能性发挥的基础[8]。本研究发现近自然经营8年后马尾松人工林林下物种多样性格局及潜在生态过程发生较大的变化,但是不同地区、不同群落对不同经营方式响应机制存在一定差异。具体来看,近熟林择伐导致灌木群落物种均匀度增加,而中龄林间伐有降低灌木群落均匀度趋势;与之相对应的是,两种经营模式均導致草本群落的物种均匀度降低,且物种丰富度也有降低的趋势。从群落排序的结果来看,近自然经营显著改变了灌木群落的物种组成,但对草本的物种组成影响不显著。通过物种多度格局拟合结果来看,近自然经营导致灌木由生态位过程逐渐转向中性过程,而对草本影响不显著,影响草本物种共存更关键的因素是地域限制。
影响物种多样性格局的因素包括扩散限制、环境筛效应及生物间相互作用等多方面[13]。在相对郁闭的马尾松近熟林内,较差的林下光环境作为环境筛很大程度上会限制林下物种(尤其是喜光的阳性物种)建成。相对狭窄的环境条件造成林下的灌木在生态位上存在高度重叠的现象,加剧种间竞争,竞争的不对称性容易造成一部分物种占绝绝对优势地位,而另一部分物种的生存空间狭小,物种多度格局就表现出极大的不均匀性[14](表2、图2)。近熟林择伐减少了林分密度,增加了透光度,光照斑块的增加改善林下光环境,削弱了环境筛作用强度,缓解了种间竞争作用。当竞争作用较强时,物种间的共存机制主要由生态位过程主导,而环境筛作用削弱减缓了种间竞争,主导物种共存的潜在生态学过程逐渐转向随机过程[15-16](图2、表4)。而中龄林林下光环境质量较好,光照条件并没有严格限制物种多样性格局及其共存机制,部分样地内物种间竞争作用较弱,通过间伐方式而增加的光环境质量并不能从根本上改变物种间的相互作用形式,因此,中龄林内间伐对物种共存机制的影响相对较小[17-18]。
研究发现,相对灌木群落而言,近自然经营对草本群落的物种组成及其潜在生态学过程影响相对较弱,而地域性差异对其物种组成及潜在生态过程影响更大。物种共存机制的空间尺度依赖特性以及物种本身的习性可能是该现象的主要机理[19-20]。影响物种共存的因素(气候、地形、土壤、小环境、生物间相互作用)有强烈的空间尺度依赖特性。气候往往在较大的地理尺度上对物种组成及共存机制起到绝对支配作用,地形因素是在中等空间尺度上的关键因素,土壤及其它因素构成的微环境在较小的空间尺度上影响较大,在最小的邻体尺度上生物间相互作用对物种多样性格局起到支配作用[21]。而不同生活型的物种所受影响的尺度不尽相同,普遍来看,木本植物所受的影响尺度相对较大,而草本植物同种生态学过程所发生的空间尺度较小。息烽和开阳地理位置有可能导致区域物种库等差异巨大,进而对物种组成起到支配作用而掩盖了近自然经营对其物种多样性格局的影响[22]。此外,草本群落的微环境除受乔木层影响外,灌木群落结构差异所导致的林下光资源二次分配对其影响更为强烈,近自然经营显著影响了灌木群落的物种组成和结构(图1),从而改变了林下光资源的二次分配,但分配的方向具有极大不确定性,这种不确定性导致草本群落潜在生态学过程的不确定性,从而呈现出一种中性格局[23]。另一方面,物种属性的差异也有可能造成现有格局。从生态策略上来看,灌木相对于草本植物而言更倾向于忍耐型生态策略,当资源(例如光资源)从紧张变为充裕时,原有生态位空间会释放,生态过程表现为从竞争趋向中性,而草本植物更倾向于杂草性策略(具有逃避型特征),资源状况并不能从根本上决定物种间的相互作用形式[24],因而潜在的生态过程存在极大的不确定性,这就导致从拟合结果上来看,生态位占优和中性过程占优均有可能,究竟是什么格局占优,并不完全取决于经营模式,而更多取决于影响其物种多样性格局诸多过程中相对作用强弱。
综上所述,近自然经营改变了马尾松人工林林下灌木物种间的相互作用模式,进而影响了灌木群落物种组成和多样性格局。但近自然经营对林下草本群落影响相对较小,存在极大的不确定性,草本群落结构和物种组成更多的是受地域限制和林分生境的影响。
参 考 文 献:
[1] BENAYAS JMR,NEWTON AC,DIAZ A,et al.Enhancement of biodiversity and ecosystem services by ecological restoration:a meta-analysis[J].Science,2009,325:1121-1124.
[2] MORENO-MATEOS D,ALBERDI A,MORRIN E,et al.The long-term restoration of ecosystem complexity[J].Nature Ecology and Evolution,2020,4:676-685.
[3] VERHEYEN K,VANHELLEMONT M,AUGE H,et al.Contributions of a global network of tree diversity experiments to sustainable forest plantations[J].Ambio,2016,45:29-41.
[4] 姜春武,徐庆,张蓓蓓,等.马尾松光合生理特性及资源利用效率研究进展[J].世界林业研究,2017,30(4):24-28.
[5] 陆元昌.近自然森林经营的理论与实践[M].北京:科学出版社,2006.
[6] 陆元昌,张守攻,雷相东,等.人工林近自然化改造的理论基础和实施技术[J].世界林业研究,2009,22(1):20-27.
[7] BONGERS F J,SCHMID B,SUN Z,et al.Growth–trait relationships in subtropical forest are stronger at higher diversity[J].Journal of Ecology,2020,108:256-266.
[8] SCHULDT A,ASSMANN T,BREZZI M,et al.Biodiversity across trophic levels drives multifunctionality in highly diverse forests[J].Nature Communications,2018,9:2989.
[9] NORDEN N,CHAZDON R L,CHAO A,et al.Resilience of tropical rain forests:tree community reassembly in secondary forests[J].Ecology Letters,2009,12:385-394.
[10] PRADO P,MIRANDA M.Package ‘sads:maximum likelihood models for species abundance distributions[EB/OL].cran r-project org/web/packages/sads/index html,2014.
[11] VOLKOV I,BANAVAR J R,HUBBELL S P,et al.Neutral theory and relative species abundance in ecology[J].Nature,2003,424:1035-1037.
[12] R DEVELOPMENT CORE TEAM.R:A language and environment for statistical computing[M].Vienna,Austria:R Foundation for Statistical Computing,2014.
[13] 牛克昌,劉怿宁,沈泽昊,等.群落构建的中性理论和生态位理论[J].生物多样性,2009,17(6):579-593.
[14] WU A,DENG X,HE H,et al.Responses of species abundance distribution patterns to spatial scaling in subtropical secondary forests[J].Ecology and Evolution,2019,9:5338-5347.
[15] SIMONS N K,LEWINSOHN T,BLTHGEN N,et al.Contrasting effects of grassland management modes on species-abundance distributions of multiple groups[J].Agriculture,Ecosystem and Environment,2017,237:143-153.
[16] VILLA P M,MARTINS S V,RODRIGUES A C,et al.Testing species abundance distribution models in tropical forest successions:Implications for fine-scale passive restoration[J].Ecological Engineering,2019,135:28-35.
[17] XU C,WANG Z,LI Z,et al.Grazing intensity and climate factors shape species abundance distribution by influencing different components of plant communities in a desert steppe[J].Ecological Research,2019,34:730-742.
[18] KOMONEN A,ELO M.Ecological response hides behind the species abundance distribution:Community response to low-intensity disturbance in managed grasslands[J].Ecology and Evolution,2017,7:8558-8566.
[19] BROWN C,BURSLEM D F R P,ILLIAN J B,et al.Multispecies coexistence of trees in tropical forests:spatial signals of topographic niche differentiation increase with environmental heterogeneity[J].Proceedings of the Royal Society of London B:Biological Sciences,2013,280(1764):20130502.
[20] GRAVEL D,GUICHARD F,HOCHBERG M E.Species coexistence in a variable world[J].Ecology Letters,2011,14:828-839.
[21] CZORTEK P,ORCZEWSKA A,DYDERSKI M K.Niche differentiation,competition or habitat filtering? Mechanisms explaining co-occurrence of plant species on wet meadows of high conservation value[J].Journal of Vegetation Science,2021,32:e12983.
[22] HELM A,ZOBEL M,MOLES A T,et al.Characteristic and derived diversity:implementing the species pool concept to quantify conservation condition of habitats[J].Diversity and Distributions,2015,21:711-721.
[23] YUAN Z,GAZOL A,WANG X,et al.What happens below the canopy? Direct and indirect influences of the dominant species on forest vertical layers[J].Oikos,2012,121:1145-1153.
[24] RGER N,BERGER U,HUBBELL S P,et al.Growth strategies of tropical tree species:Disentangling light and size effects[J].Plos one,2011,6:e25330.
