典型冰川退缩区铅的来源、累积及历史沉降
——以青藏高原海螺沟冰川退缩区为例
2021-09-03陈霈嘉王定勇西南大学资源环境学院重庆40075重庆市农业资源与环境研究重点实验室重庆40076
陈霈嘉 ,王 训 ,王定勇 ,2* (.西南大学资源环境学院,重庆 40075;2.重庆市农业资源与环境研究重点实验室,重庆40076)
铅(Pb)是联合国环保署发布的全球共同关注的优先控制污染物,在环境中具有持久性、不可降解性和生物累积性,会对生态环境、粮食安全和人类健康产生极大的影响[1].土壤、植被和径流中的异常铅含量水平主要受到来自人为活动产生的铅排放与大气沉降输入的影响[2].青藏高原被称为地球的“第三极”,虽然人类活动较少,但毗邻人口数量众多且发展迅速的中国东部以及南亚,这些区域人为排放的铅可能会吸附在大气颗粒物上,通过长距离迁移传输进入青藏高原生态系统中造成铅的累积,产生系列的环境风险[3-4].
近年来全球气候变暖进程加快,冰川与冻土加速消融,这为青藏高原重金属污染相关的环境问题增添了新的变数.一方面,冰川与冻土累积的重金属随冰川冻土的消融过程快速进入到下游的水生生态系统中,对野生动物与人类健康造成潜在的环境威胁[5];另一方面,快速变化的生态环境也为研究铅等重金属在生态系统中的累积提供一个较好的研究窗口.冰川消融后,使得原本被冰川覆盖多年的区域重新暴露在大气中,随后的植被演替而形成的冰川退缩区会从天然背景值开始,逐渐地累积来自大气中的重金属沉降[6-8].考虑到冰川退缩区植被的良好时空序列,与局地基本一致的气候条件,在连续相邻的样地上同时开展相关研究,比较各自样地研究结果的差异性,建立时间序列的变化情况,从而反演历史大气沉降铅的大致速率(精准速率反演还应考虑铅在环境的迁移)与在环境中的迁移归趋,解析铅在退缩区植被演替序列中土壤~植被系统中的关键生物地球化学过程.这为认识铅在陆地生态系统中累积速率、迁移方式及分配机制提供数据支撑与理论基础;同时也为未来探究全球变化~植被格局改变对铅的生物地球化学循环的影响提供经典案例与理论探索.
本研究通过位于青藏高原东部边缘的贡嘎山冰川退缩区植被演替序列这一天然实验平台,定量评估贡嘎山海螺沟在不同历史时期所形成的冰川退缩区土壤和植物中铅的含量与储量的分布格局差异,阐明铅在土壤和植被中迁移、分配与累积过程,解析铅在退缩区的来源贡献与历史沉降铅的累积速率,为理解全球变化对青藏高原重金属的生物地球化学循环环境影响评估提供经典范例与基础数据,特别是为未来全球变化对铅的环境地球化学过程影响提供了经典案例与数据理论支撑.
1 材料与方法
1.1 样地介绍与样品采集
以青藏高原东部的贡嘎山海螺沟冰川退缩区(29.6°N, 102.0°E,海拔 2600~2920m)为研究对象.海螺沟冰川属于季风海洋性冰川,年平均气温为 4.2℃,年降水量为1800~2000mm[9],水热条件好,冰川消融速度快,土壤有连续成土过程,其冰川退缩区存在自小冰期以来形成的完整植被演替序列.自 19世纪中期以来,海螺沟冰川退缩明显,无冰进过程,在距 2012年底的冰川末端形成了长达2km的完整植被原生演替序列,该小区域气候和水文条件差异不大,来自大型动物的干扰很小.根据前期的样带调查,海螺沟冰川退缩区的植被演替规律是以柳树、沙棘和冬瓜杨的先锋树种向冷杉或云杉顶极树种演替的过程.利用不同样地的优势木(冬瓜杨)与原生裸地的关系,可确定近百年内样地年代,进而确定当时冰川末端的位置;大于 80年的样地,应用原生演替过程中种间替代关系,采用峨眉冷杉优势木确定年代[6-7].详细有关冰川退缩区年代的确定与各时期植被群落关系,见前人研究资料[5-7].简言之,1980~2005年形成的冰川退缩区是各先锋植物发展时期;1958~1980年样地,冬瓜杨在竞争中处于优势,柳树和沙棘逐渐枯萎而亡;1958~1930年形成的冰川退缩区样地处于先锋植被向顶级群落(云冷杉)过渡时期;1890~1860年样地,形成了云冷杉为顶极群落的高山暗针叶林生态系统[10].
于 2016~2018年间采集了冰川退缩年代为1860~2005年的站点土壤和植物样品,其中土壤的采样点为9个(退缩时间分别为1990年、1985年、1980年、1973年、1966年、1958年、1936年、1890年和1860年),植被的采样点为12个(退缩时间分别为2000年、1990年、1985年、1980年、1973年、1970年、1966年、1958年、1936年、1930年、1890年和1860年).在各样地站点设置面积为10m×10m样方,每个样地有3个重复样方.
按照土壤层次分层采集土壤样品.由于退缩区百年尺度的演替,无明显的矿质层土壤发育,土壤层次主要为森林有机土壤(Oi, Oe与Oa)及C层冰碛沉积物.对于植被样品,由于冰川退缩区组成群落的树种简单,将分别采集所有的树种.枝、叶、树皮、树干按不同方位、不同部位按比例采集混合样;按照不同的方位采集细根、中根和粗根.对于地表草本、苔藓和地衣,每个样方取3个1m×1m的代表性小方框采集;此外还采集样方 0~2m 处树上苔藓与地衣.详细记录各样方基本土壤(如pH值、采样体积等)与植被群落参数,并利用之前研究得到的解析木方程[11],结合树胸径与冠层参数估算植被生物量.
1.2 样品处理和测定
土壤样品在黑暗无尘室中风干,然后用玛瑙研磨过 200目(74μm)的筛;植物样品用电动研磨机研磨.在研磨前,记录所有土壤风干样品的重量,以便计算土壤容重.采用电感耦合等离子体质谱法(ICP-MS)测定土壤和植物样品中微量元素的含量,测量的元素为: Pb、Cr、Zn、As、Sr、Mo、Cd、Ba、U、Li、Be、V、Mn、Co、Ni、Cu、Ga、Rb、Cs和Tl共21种,方法步骤参照美国EPA方法6020B(EPA方法6020B)[12].植被和土壤的标准物质均来自国家质量技术监督局(土壤标准物质为 GBW07405,植被标准物质为GBW10020),每10个样品测量一次标准物质,土壤标准物质的平均回收率为 102%~107%,植被标准物质的平均回收率为 88%~103%.此外,每30个样品中有3个空白样品进行了分析,以确保样品在测量过程中没有受到污染,空白样品中微量元素含量均小于被测样品的1‰值.
1.3 统计计算和分析
富集因子可用于分析元素富集程度,为后续铅的源解析结果判别提供一定参考[13].选取土壤中主要来源于风化壳且含量较为恒定的 Li元素作为参比因子,计算土壤和植物体中铅的富集因子[14]:

式中: EF是富集因子,C是植物、土壤和岩石样品中的Pb含量,R是Li元素含量.根据前人的研究结果[15],对Pb的累积程度按以下方法分级:EF<1,无/低累积,元素主要来自地壳源;1≤EF<3,中度累积;3≤EF<6,高度累积;6≤EF,极度累积.结合测量所得的土壤和植被中含量与样地土壤容重和植被生物量信息,估计每个样点铅储量如下:

将铅的来源分为冰碛沉积物风化来源和大气沉降来源,并利用主成分分析(PCA)方法,辨析相应来源贡献的百分比[16].进行PCA分析前,需进行相关统计预分析.根据相关分析的结果,拟选取的元素包括 Cr、As、Sr、Ba、U、Li、V、Mn、Co、Ni、Cu、Ga、Rb、Cs、Tl、Pb、Cd、Mo 及 Zn.其中 KMO统计检验表明,有机土壤(包括Oa, Oe及Oi层)相关重金属元素集合的KMO值为0.882,因而本研究的数据结果十分适合进行PCA分析.在PCA分析方法中,采用凯撒正态化最大方差法进行旋转.最后在PCA 分析基础上,通过载荷因子的权重,估计不同来源的百分比贡献(C%)如下:

式中: j为该元素在成分矩阵中的载荷因子.考虑到铅储量大小几乎与冰川消退时间呈线性关系(1890年样地站点除外),利用冰川退缩时间与铅储量大小的斜率以及大气沉降的贡献比例,进而估算大气来源铅在退缩区的平均累积速率(V):

式中: a为系统中Pb库对时间的一阶微分斜率; C为大气来源的贡献率.
2 结果与讨论
2.1 冰川退缩区中铅的含量变化特征
图1a表明退缩时间为1860年~1990年的样地土壤平均铅含量为(28.46±14.77) mg/kg,各层的铅含量: Oa 层[(41.02±16.58) mg/kg] > Oe 层[(32.88±14.79) mg/kg] > C 层[(22.94±5.11) mg/kg] > Oi层[(17.00±7.30) mg/kg] (P<0.05,单因素方差分析).随着样地退缩时间增加,大气沉降来源的铅的累积效应愈为显著,故而土壤铅含量随时间递增(图1),O层土壤铅含量以1936年样地最高(Oi层29.61 mg/kg,Oe层52.03 mg/kg, Oa层68.52 mg/kg),为1990年样地的 5.2倍;C层土壤铅含量以 1860年样地最高(32.69 mg/kg),为1990年样地的2.1倍.
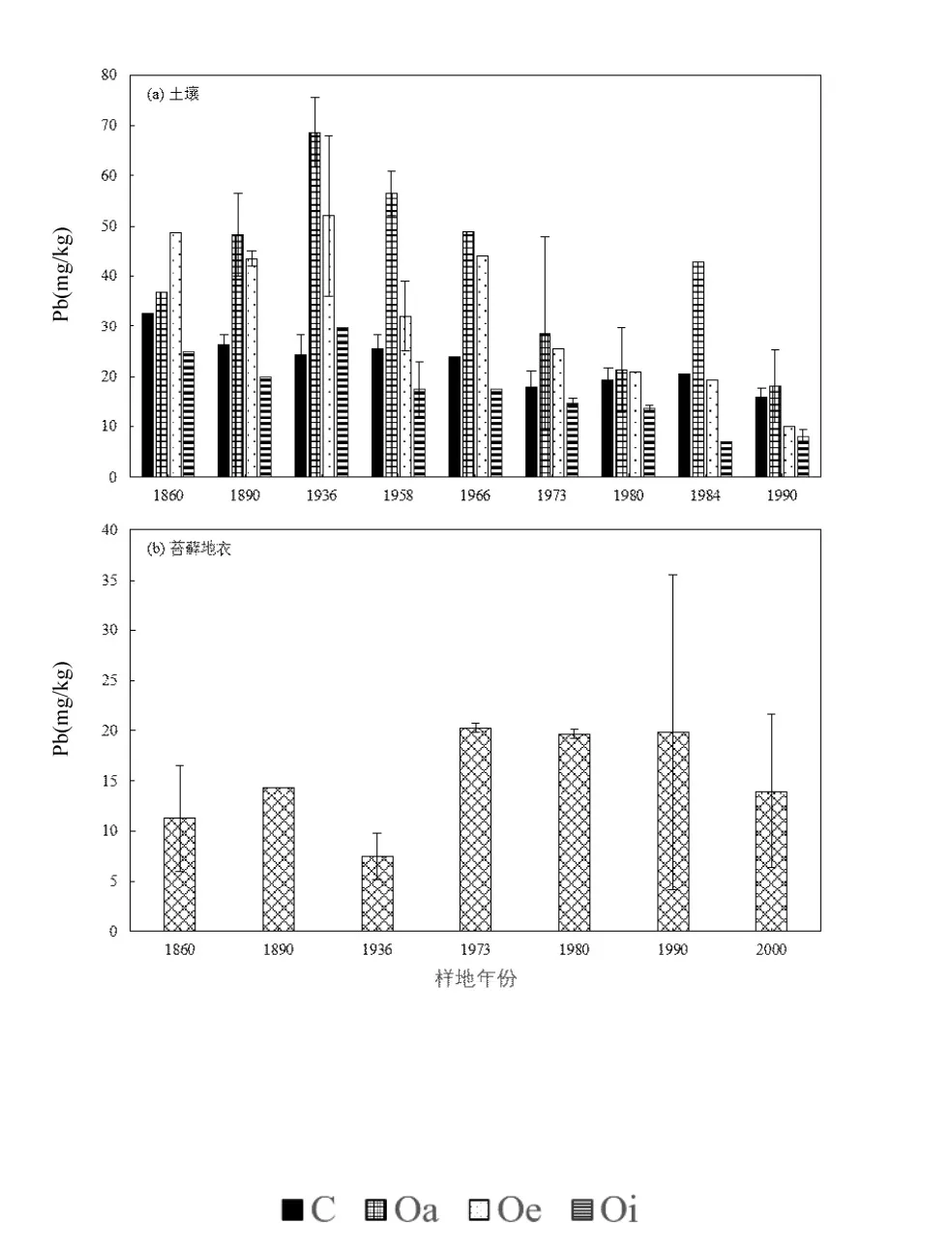
图1 海螺沟冰川退缩区土壤与苔藓地衣的铅含量变化趋势Fig.1 Variation of Pb concentration in organic soil profiles,moss and lichen at Hailuogou glacial retreat area
各样地土壤中铅含量的动态变化是大气~植被~土壤系统相互作用的综合结果[17].冰川消融前,冰川将其下方的基岩和大气隔绝,冰川运动的研磨和粉碎作用,导致基岩形成冰碛物,故冰碛物铅含量受控于基岩中含量.冰川退缩之后,退缩区的土壤开始接受来自大气的铅沉降.大气沉降的铅与土壤中许多的组分发生吸附络合作用,包括铁锰氧化物、碳酸盐、粘土、有机质和硫酸盐等[18].O层为土壤的表层,由枯枝落叶和生物残体堆积分解而成,其铅含量随着土壤腐质化程度的增加而增高.因为土壤有机质对重金属元素具有强吸附络合作用[19],可显著促进铅的累积[20];土壤中 Oa层分解最完全,故有机质与铅含量均最高.此外,退缩区形成的时间越久远,铅与有机质相互作用时间也越明显,其有机土壤累积的大气沉降来源铅也越高.C层土壤不易受外部环境干扰,铅含量显著低于 O 层(P<0.05,单因素方差分析),冰川前缘土壤铅含量在一定程度上反映了当地的土壤背景值,海螺沟冰川前缘土壤铅含量为25.7±2.9 mg/kg[3],与中国大陆土壤的铅含量背景值[(26.0±12.37) mg/kg]相近[21].C层中铅含量随着土壤发育和植被演替而增加,但其含量在整个植被空间序列中的变异性小于 O层土壤的变异性.其相关原因是由于大气沉降来源的铅小部分淋溶至 C层[22],增加C层铅含量.
本研究中云冷杉顶极群落(海拔 2780~2900m)处于退缩时间为 1860~1936年的样地,发育成熟,O层 含 量 为 (41.33±15.19) mg/kg (19.99~68.52mg/kg),C 层含量为(27.76±4.40) mg/kg (24.22~32.69mg/kg),土壤平均铅含量为(37.94±14.45) mg/kg. Luo等[3]2013年在贡嘎山不同海拔处采集的样品中,海拔为2780m的针叶林中O层的铅含量为(64.3±3.8) mg/kg,C层铅含量为(29.4±6.9) mg/kg,海拔为 3200m的针叶林中O层的铅含量为(50.9±3.5) mg/kg, C层铅含量为(31.4±5.1) mg/kg.该采样地为原生针叶林,发育时间更长,土壤铅含量和本研究1860~ 1936年的样地相近,这说明海螺沟冰川退缩区百年尺度的铅沉降累积,可使得植被演替序列中土壤铅浓度与当地的地带性植被区域相当.柏建坤等[23]2007年在可可西里采集的 25个表层土壤样品中,平均铅含量为16.54mg/kg.杨安等[24]2013年 7~8月沿青藏高原东北~西南方向在表层土壤(0~20cm)采集的样品中,平均铅含量为 29.08mg/kg.通过比较可以得出,上述非森林土壤的铅含量与本研究1860年~ 1936年样地的土壤C层土壤相当,但较O层土壤含量偏低.其主要的差异是因为森林土壤具有较高的有机质,对大气沉降的铅具有强累积作用.这也进一步证实了森林有机层土壤是大气铅的重要汇集区[25-27].
在植被系统中(图 2),地下部铅含量:细根[(1.73±1.81) mg/kg] > 中根[(0.69±0.65) mg/kg] >粗根[(0.45±0.35) mg/kg] (P < 0.05,单因素方差分析).之前的研究也表明,细根中有较高的铅含量[28].这是因为细根是功能性根,是植物吸收水分和养分的主要途径[29],根系产生的有机分泌物不但能活化土壤中的铅,还能与铅形成有机络合物,改变铅在土壤中的流动性和生物有效性[28].在本研究中,细根和O层铅含量相关系数为0.89(P < 0.05,T检验),这充分说明 O层土壤中的铅会被细根吸附累积[30].植物地上部铅含量:树枝[(2.10±1.13) mg/kg] > 树皮[(1.19±0.79) mg/kg] > 树叶[(0.78±0.17) mg/kg] > 树干[(0.27±0.32) mg/kg](P < 0.05,单因素方差分析).树干中的铅含量最低,归因于铅在植物体中较难的迁移传输[31].因多孔结构,树皮能直接吸附大气干湿沉降的铅,故树皮铅含量较高.由于植物的解毒机制,大多数重金属倾向于储存在生物活性较弱的组织和代谢不活跃的细胞器中[32].和树叶相比,枝条生物活性较弱,造成枝条具有一定的铅积累特征[3].随着退缩时间的推移,植被的铅含量变化趋势和生长趋势相关,处于生长期的植被吸收铅的能力大,铅含量升高,演替竞争过程中植被逐渐死亡,吸收铅的能力下降,铅含量降低.植物地上部的铅含量变化趋势与土壤不一致,整个时间序列样地中土壤平均铅含量约为植物平均铅含量的28倍,这是由于土壤中的铅生物可利用性较低,难以大量直接被植物吸收[33]所致.

图2 海螺沟冰川退缩区柳树、沙棘的铅含量、冬瓜杨与云冷杉的铅含量Fig.2 Pb concentration in willow, sea buckthorn, big leaf poplar, dragon spruce and fir at Hailuogou glacial retreat area
苔藓地衣没有根部和角质层系统,与土壤等基质接触少,几乎完全从大气干湿沉降中吸收铅[22,27].苔藓地衣铅含量均值为(15.26±4.89) mg/kg (7.49~20.23mg/kg,图1b),是演替序列乔木整体平均铅含量的3.9倍.这是因为苔藓地衣生理结构简单、比表面积大,重金属跨细胞壁较为容易,可在整个表面进行交换吸收[34-36].1860~1936年的样地中苔藓地衣铅含量比 1973年后的样地低 1~2倍,可能的原因是1860~1936年的样地为云冷杉顶极群落,而与晚于1973年样地的阔叶林相比,针叶林的林冠对大气沉降铅颗粒的截留更大[37],致使早于1936年的样地中苔藓地衣铅含量相对较低.
锂被认为是地壳的重要组成部分,背景土壤中的锂含量与硅酸盐风化过程高度相关[38-39],随着冰川的消退,Li库的大小并没有随着时间的增加而明显变化 (C层平均锂含量为(43.32±9.75) mg/kg,其中1936年之前样地铅含量为(34.29±4.90)mg/kg, 2015年样地含量为(34.26±10.91)mg/kg; P > 0.05,独立样本t-检验).因此,表土植被中铅和锂含量的比值可反映是铅相比于冰碛物的累积程度.图 3中,随着冰川退缩时间的推移,有机土壤富集因子增大(2.33~17.45),在1936年样地达到峰值(Oa=17.45,Oe=12.80和Oi=8.68)(EF>6),说明大气沉降的铅在有机土壤中有较大富集.图 4的植被富集因子中,云冷杉(EF=13.04±9.63)>冬瓜杨(EF=13.00±18.00)>沙棘(EF=9.53±13.69)>柳树(EF=5.69±6.71),说明云冷杉和冬瓜杨吸收铅的能力较柳树和沙棘大.不同组织器官之间富集因子相差较大,以1990年样地沙棘中根最小(EF=0.63), 1990年冬瓜杨枝最大(EF=52.98).云冷杉顶极群落时期(早于 1936年的样地),冬瓜杨细根和树干的富集因子是其他样地的 4~5倍.这是因为随着冰川退缩时间的推移,大气累积铅的效应愈发明显,土壤铅的含量显著升高,故而植被树根与树干中铅含量也随之增加.

图3 海螺沟冰川退缩区有机土壤的富集因子Fig.3 Enrichment factor of organic soil at Hailuogou glacial retreat area

图4 海螺沟冰川退缩区柳树、沙棘、冬瓜杨与云冷杉的富集因子Fig.4 Enrichment factor of willow, sea buckthorn big leaf poplar, dragon spruce and fir at Hailuogou glacial retreat area
2.2 冰川退缩区中铅的储量变化
图5描述了贡嘎山1860~2005年样地有机土壤和植被中的铅储量动态变化.有机土壤铅储量:Oa层(6461.25mg/m2)>Oe 层(1388.94mg/m2)>Oi层(400.57mg/m2);植被铅储量:云冷杉(81.64mg/m2)>冬瓜杨(35.66mg/m2)>柳树(7.16mg/m2)>沙棘(4.49mg/m2).有机土壤铅储量于 1936年样地最高(2663.25mg/m2), 植被铅储量于 1890年样地最高(31.91mg/m2).在晚于1936年的样地中,铅储量随着样地年龄的增加而增大,与铅含量在时间序列上的变化规律一致.冰川退缩时间早于1936年的样地铅储量较小,且铅储量随着退缩时间的推移而减少.可能的原因:一是,早于1936年的样地主要为多年生针叶林,其凋落物的分解速度缓慢,不能迅速将铅从树叶或树枝快速释放到土壤中[22],晚于1936年的样地以阔叶林为主,枯枝落叶更多,由于其较快的分解速度,凋落物铅的归还速率较快;二是,早于1936年的样地土壤发育成熟,有机腐殖酸高,使土壤中的 pH 值降低,使得部分惰性铅转化为铅离子,从土壤中流失;三是1936年左右的样地苔藓生物量很大,其固定大气沉降铅的能力显著,致使1936年有机土壤和植被总系统中铅储量最大.

图5 海螺沟冰川退缩区Oi层、Oe层、Oa层、有机土壤、植物与有机土壤和植被总系统中的铅储量动态变化Fig.5 Variations of Pb pool size in Oi, Oe, Oa, organic soil,vegetation, organic soil and vegetation at Hailuogou glacial retreat area
各个时间序列上退缩的样地中(图 6),相应阶段树种铅储量最高值为:1980年样地的柳树(1.04mg/m2),1990年样地的沙棘(1.53mg/m2),1966年样地的冬瓜杨(6.146mg/m2)与 1890年样地的云冷杉(28.66mg/m2).在早于 1930年的样地中,云冷杉是主要储存铅的树种,在 1958年~2005年样地中,冬瓜杨的铅储量达到了植被总系统铅储量的70%.这是因为植被的铅储量变化趋势和生长趋势相关,在植被生长期,铅储量因生物量增加而不断升高,并在植被生长最旺盛的时期达到最大,随后随着演替过程中植被的死亡而降低.在整个时间序列上的不同的树种中,植被整体(包括根、树干、树皮、枝条和叶片)的总铅储量:云冷杉(81.64mg/m2)>冬瓜杨(35.66mg/m2)>柳树(7.16mg/m2)>沙棘(4.49mg/m2).考虑到不同树种间铅的含量变异性远小于生物量的变异性,云冷杉与冬瓜杨较大的总铅储量因为具有显著更高水平的生物量所致.对于不同的植被器官中,植物地下部中:粗根的铅储量较细根和中根大;地上部:枝条、树皮和树干的铅储量较大,叶片的铅储量较少.粗根、树干和叶片的铅储量分布与含量不一致,是因为铅储量受植物组织的铅含量,特别是生物量的共同影响.在整个植物体中,粗根和树干的生物量占比很大,而树叶的生物量相对少.
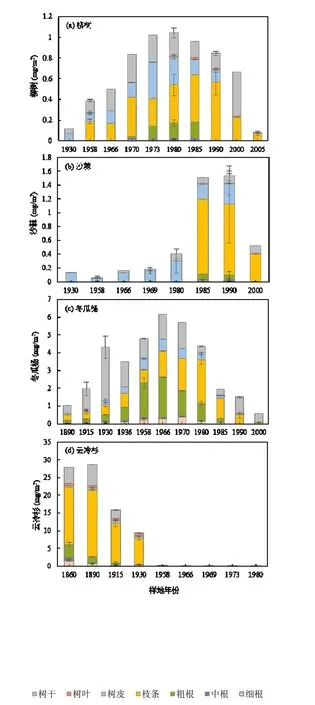
图6 海螺沟冰川退缩区柳树、沙棘、冬瓜杨与云冷杉的铅储量Fig.6 Variations of Pb pool size in willow, sea buckthorn, big leaf poplar, dragon spruce and fir at Hailuogou glacial retreat area
2.3 海螺沟冰川退缩区中铅的来源和累积速率分析
有机土壤 PCA分析结果中,共提取了两个主成分,解释了原始数据89.61%的变异度.其中成分1具有Li、Co、Ni、Cu等高载荷值的元素,成分2具有Cd、Zn、As等高载荷值的元素.考虑到有机土壤中的 Li、Be、V、Co、Ni、Ga、Cs、Sr、Ba 等元素主要为壳源[40],受地壳物质的自然风化和侵蚀等[41]等地质过程的影响更大,所以成分 1反映冰碛沉积物风化过程;而环境中的Cd, As和Zn大部分来自于冶炼、采矿、钢铁铸造厂、化石燃料的燃烧等人为活动排放[40,42-44],所以成分2反映人为排放源的大气沉降过程.通过 Eq.(3)计算,退缩区有机土壤中,冰碛沉积物风化过程和大气沉降过程对铅的贡献百分比分别为2.79%和57.15%. 值得注意的是,铅在有机土壤中有 40.06%的变异性没有得到解释,本研究暂且归于其他自然过程(如冰河相关的水土迁移过程等),并期望未来予以进一步研究. Bing等[4]2012年在海螺沟冰川退缩区采集的样品中,利用铅同位素比值建立的二元混合模型表明,人为源的铅对 O层的贡献率为 45.2%~61.3%,这与本研究的源解析值相近.大气沉降过程的高贡献百分比说明人为排放铅的大气沉降是贡嘎山有机土壤中铅的主要来源.之前的研究[4]通过混合单粒子拉格朗日积分轨迹模型(HYSPLIT)计算后向轨迹发现,贡嘎山的铅沉降会受到来自云南西北部、四川东北部[3]及南亚地区长距离跨境传输的气团影响(印度、孟加拉国等[45-46])[4].
利用冰川退缩时间与铅储量的斜率以及有机土壤来自人为源的铅沉降比值,初步估算了冰川退缩区的有机土壤中铅的大气累积速率(式 4).同时也必须指出,该假设存在较大的不确定性,这是因为基于统计学的源解析方法,其本身存在较为明显的不确定性,会导致估算的大气来源铅累积速率产生未知误差.在冰川退缩时间为 1890~1990年的有机土壤中,大气来源铅的总累积速率为(8.87±3.55)mg/(m2·a),土壤各层的累积速率:Oa 层[(5.01±2.01)mg/(m2·a)] > Oe 层[(1.64±0.66) mg/(m2·a)] > Oi层[(0.56±0.22) mg/(m2·a)].Oa 层最高的累积速率再次反映了有机质对铅的强吸附络合作用.
考虑到铅在土壤中强吸附固定作用,大气来源铅在生态系统中累积速率理论上略小于大气铅沉降速率,可看作是大气铅沉降的近似.通常铅在工业和城市地点的累积速率最高,其次是农业、郊区和偏远地区,这种空间格局与当地排放密切相关[47].例如,铅在中国北方的大气总沉降通量平均为 26.5mg/(m2·a),在城市和工业地区,铅沉降通量可达到45.8mg/(m2·a),农业用地和偏远地区的铅沉降通量分别为 16.2mg/(m2·a)和 14.1mg/(m2·a)[47].贡嘎山为偏远山区,当地无工业排放、人类活动远低于上述区域,所以大气沉降的铅累积速率明显低于中国大部分经济发达地区域.表 1展示了部分世界发达国家地区以及一些欠发达地区,这些地区铅的累积速率普遍<4.2mg/(m2·a),仅有 Tokyo Bay [9.9mg/(m2·a)]高于贡嘎山,主要原因是这些地区产业结构发展完善,以第三产业为主,本国和邻近地区无重工业发展,外源污染的大气沉降少[14].与世界其它地区的比较中可以得出,贡嘎山虽为偏远山区,但存在一定的铅污染.这印证了中国西南地区和南亚的人为排放的铅会通过长距离迁移传输,沉降进入青藏高原东部的高山地区,造成生态系统的铅含量升高.

表1 世界上部分地区总沉降的铅累积速率[47]Table 1 Accumulation rate of total Pb deposition in parts of the world
3 结论
3.1 土壤各层铅含量随着冰川退缩时间增加,O层含量最高.植被铅含量以树枝、细根和树皮最高,树干最低.整个冰川退缩区中,大气沉降的铅都有较大的富集.
3.2 有机土壤铅储量随着样地年龄和土层深度增大,植物对铅的吸收相对有限,不同种类植物累积铅的能力具有差异性;铅储量变化与植被演替规律、植被铅含量及生物量相关.
3.3 人为排放污染的大气沉降是贡嘎山中铅的重要来源.在冰川退缩时间为1890~1990年的有机土壤中,大气沉降铅的平均累积速率为(8.87±3.55)mg/(m2·a),显著低于我国大部分经济发达区域的速率.
