广西弄岗五桠果叶木姜子群落木质藤本空间格局及其与树种间的关系
2021-09-01李先琨李冬兴黄甫昭李健星陆树华郭屹立文淑均向悟生
陆 芳,李先琨,王 斌,李冬兴,黄甫昭,李健星,陈 婷,陆树华,郭屹立,文淑均,向悟生,*
1 广西师范大学生命科学学院, 桂林 541006 2 广西壮族自治区中国科学院广西植物研究所广西喀斯特植物保育与恢复生态学重点实验室, 桂林 541006 3 广西友谊关森林生态系统国家定位观测研究站, 凭祥 532699
我国喀斯特地区占国土面积高达1/3左右,尤其在西南地区,有岩溶发育最强烈、生物多样性丰富的喀斯特景观,在全球生态系统中具有范例级的重要地位[1]。广西弄岗国家级自然保护区保存有生态系统较完整的喀斯特季节性雨林,是西南喀斯特森林的典型代表,该区域森林具有生态结构复杂、物种多样性丰富、特有成分突出等特点[2]。五桠果叶木姜子群落是弄岗喀斯特季节性雨林中具有代表性和典型性的独特森林类型,在保护区仅分布于约2hm2的封闭洼地内,群落保存完整、原生性强,是中国境内唯一在喀斯特地区发现有该群落分布的区域。群落优势种五桠果叶木姜子为狭域分布的珍稀植物,仅零星分布于云南西南部和广西西南部,被IUCN红色名录中列为濒危物种[3]。研究五桠果叶木姜子群落植物种间的相互关系,将有助于探明喀斯特季节性雨林的生态过程,促进该区生物多样性保护和生态系统功能维持,也有利于极小植物种群的保育。
木质藤本是一类不能单独直立、需借助其他植物和支撑物才能直立生长的木本植物[4]。在大多数热带森林中,10%—45%的树木攀附有藤本植物[5-6]。同时,它们的总生物量仅次于树木[7]。因此,木质藤本在森林生物多样性、结构和功能等方面都占据重要地位,影响着森林群落动态与生态过程[8-9]。喀斯特森林中的木质藤本种类丰富,广西喀斯特植被中有较为独特的藤刺灌丛类型,在广西喀斯特植被96个群系中,藤刺灌丛占了28个[10],弄岗国家级自然保护区的藤本种子植物种类丰富[11],藤本植物占种子植物区系的比例为20.8%[12],表明木质藤本在热带亚热带喀斯特植被中的普遍性和重要性。
木质藤本通过影响支持木的生长和更新过程,从而影响很多森林群落的结构和功能,因此开展木质藤本与支持木关系的相关研究,有助于深入了解森林结构及其动态规律。近年来,部分学者对木质藤本与支持木相关性的研究发现,云南干热河谷植被不同径级树木的附藤情况存在差异[13];部分常绿阔叶林中,大径级支持木被木质藤本攀附的比率高于小径级支持木,且木质藤本在支持木上呈集群分布[14];海南岛部分热带雨林中的木质藤本对支持木在科和种的水平上具有选择性[15]。此外,有研究发现木质藤本的攀附对支持木的生长死亡存在一定的影响[16-17]。总体而言,森林中木质藤本与支持木关系的研究仍然较少,很多机制仍缺乏了解,也缺乏长期监测研究,在喀斯特森林中相关研究尚未见到报道。基于此,选择典型喀斯特季节性雨林林中的五桠果叶木姜子群落为对象,通过长期监测,对木质藤本与支持木的关系开展研究,期望回答以下科学问题:(1)该群落木质藤本密度和分布格局有何特征?(2)木质藤本对不同种类、不同径级、不同聚集强度的支持木是否具有选择性攀附?(3)木质藤本对支持木的死亡过程是否存在显著影响。
1 材料与方法
1.1 研究区域概况
弄岗国家级自然保护区位于我国广西壮族自治区龙州县之东及宁明县以北(106°42′28″—107°04′54″E,22°13′56″—22°33′09″N),呈西北-东南长条状分布,分为弄岗、陇呼和陇瑞3个片区,总面积10080hm2。目前仍保存着世界上少有的、生态系统完整的喀斯特季节性雨林。弄岗国家级自然保护区地形复杂,以喀斯特峰丛深切圆洼地(谷地)为主,由多个石山山峰和镶嵌其中的洼地(谷地)组成。雨热充沛生物多样性极其丰富[18]。
五桠果叶木姜子群落在弄岗国家级自然保护区,仅分布于陇呼片一个相对封闭的洼地内,面积约2hm2,林内岩石裸露,洼地底部有季节性水淹,洼地与坡地之间的坡度较大,坡面土被不连贯,以石缝土和石穴土分布为主,土壤为棕色石灰土。
1.2 野外调查方法
1.2.1样地建立与木本植物调查监测方法
在五桠果叶木姜子群落分布集中的地方建立1hm2固定监测样地,采用全站仪划分为100个10m×10m小样方,小样方4个角作永久标记。以样地西南角为坐标原点,测量并记录树木横纵坐标。同时,将每株胸径大于等于1cm的树木高度1.3m处漆上红漆做标记。2014年记录了100个10m×10m样方内胸径大于1cm的所有树木的胸径、冠幅、树高、存活状态,挂牌编号,并鉴定到种。2019年对样地进行复查,调查方法与2014年一致,同时对胸径大于1cm树种的新增个体进行了定位、记录、挂牌和鉴定,对死亡个体也详细记录。
1.2.2木质藤本植物信息调查方法
记录缠绕、攀挂和附着于每株在编树木的木质藤本数量,在距地面高度1.3m处测量并记录木质藤本植物的胸径,并且以主要支持木的坐标为该木质藤本的坐标。
1.3 统计分析方法
1.3.1木质藤本密度及空间分布格局
木质藤本密度即单位面积的木质藤本株数,用公式表示为:
N为样方内某种植物个体的全部数目[19]。
采用点格局双关联函数,分析木质藤本在不同空间尺度上的分布格局。g(r)函数是由K函数推演而来,是K函数的概率密度函数,因其能有效剔除K函数小尺度上的累积效应而得到广泛应用。鉴于g(r)函数在细节尺度上更直观、精确的优势,利用该函数描述具体空间尺度上的种群分布格局。g(r)函数同样是基于所有定位的成对个体之间距离的格局分析,是距离依赖关联性函数,包括单变量和双变量分析,其中单变量分析的计算公式如下:
为精确分析种群的空间分布格局,采用的零模型为完全随机零假设进行了199次随机模拟,产生置信度为99%的包迹线,以检验点格局分析结果的显著性。若实际分布的g(r)值落在包迹线内,则符合零模型,为随机分布;若在包迹线以上,则呈聚集分布;若在包迹线以下,则呈均匀分布[20]。基于完全随机零模型的物种分布格局图中,r表示尺度,obs表示观测值,theo表示理论值,lo表示下包迹线,hi表示上包迹线。
1.3.2木质藤本对不同种类和生长特性支持木的选择性
(1)对不同科属种类支持木的选择性
支持木的科属类型依据《中国植物志》的分类系统划分,拉丁名主要参照《Flora of China》确定。计算不同树种的重要值,公式见参考文献[19]。统计不同科属种支持木的附藤数量和附藤率,分析木质藤本在树木科属种水平上是否存在选择性。
(2)对不同径级支持木的选择性
根据五桠果叶木姜子群落现状及其树木生长特点,参考王斌等[21]的径级划分标准,将不同支持木种群径级大小划分为3类以计算树木的附藤情况:1cm≤DBH<10cm为小径级树木,10cm≤DBH<30cm为中径级树木,DBH≥30cm为大径级树木。此外,为了保证统计模型结果的稳定性,选择个体数在15株及以上的主要树种进行分析。
物种的附藤率为某物种被木质藤本攀附的个体数占该物种总个体数的比例,10m×10m样方的附藤率为样方内被木质藤本攀附的树木个体数占树木总个体数的比例。统计样地内不同径级树木个体的附藤数量和附藤率,采用回归模型分析木质藤本胸径与树木胸径的关系。
(3)对不同聚集强度支持木的选择性
采用点格局双关联函数分析主要树种空间分布格局,方法与前述方法一致。树木的聚集强度以分析尺度内的双关联函数平均值来表示,即物种在0—20m内的g(r)平均值(g1+g2+…+g20)/20,表示为g0—20(r)[20]。采用一元回归分析附藤率、附藤数量与支持木聚集强度g0—20(r)值的关系。
1.3.3木质藤本对树木死亡的影响
根据样地2014年和2019年两次调查数据,统计不同树种的死亡率。死亡率为
M=(lnN0-lnSt)/T
其中,N0为某一种群在第一次(2014年)调查时的个体数,St为第二次(2019年)调查时该种群的存活个体数,T为两次调查的时间间隔[22]。
采用一元回归模型分析木质藤本密度与树木死亡率的关系;采用多元回归分析不同支持木的附藤数量、附藤率与支持木死亡率的关系。
以上数据分析及作图在R 3.6.3和Origin 2018中完成。
2 结果
2.1 木质藤本的密度及其空间分布格局
五桠果叶木姜子1hm2样地中木质藤本的平均密度为0.0913株/m2,在100个10m×10m小样方中,木质藤本的密度最高为0.54株/m2。其中木质藤本的密度低于0.0913株/m2高达68%,大于等于0.3株/m2仅7%(图1)。其中,木质藤本的径级在DBH<3cm占比高达89.79%,而DBH≥3cm仅占10.21%。

图1 10m×10m小样方中木质藤本的密度比例图
基于完全随机零模型分析得出,五桠果叶木姜子样地中的木质藤本所有个体(DBH≥0.1cm)及0.1cm≤DBH<3cm个体的空间分布格局相似,在0—19.5m空间尺度均表现为聚集分布,且随着空间尺度的增大,聚集强度逐渐减弱;其他空间尺度表现为随机分布;虽然两者的空间分布格局相似,但是0.1cm≤DBH<3cm个体的g0—20(r)值稍高一些。DBH≥3cm个体在1.5—9.5m、16—18m空间尺度上表现为聚集分布,聚集强度随空间尺度的增大而呈波浪式下降,且在5—9.5m、16—18m空间尺度上出现两个波峰,其他尺度表现为随机分布,g0—20(r)值为1.892,比DBH≥0.1cm及0.1cm≤DBH<3cm个体的g0—20(r)值高。从木质藤本的个体分布图也可以看出,0.1cm≤DBH<3cm个体在小尺度上表现出“斑块状”集中分布于地势平坦的中部及具有一定坡度的西南和东北部位,而西北部位在一定尺度上也表现为一定的聚集分布;DBH≥3cm个体较集中分布在中部、西南至西北部位,说明两个径级木质藤本的空间分布格局具有一定的差异(图2)。

图2 木质藤本在两个不同径级的点空间分布图、木质藤本所有个体基于完全随机零模型的分布格局图、木质藤本0.1cm≤DBH<3cm基于完全随机零模型的分布格局图、木质藤本DBH≥3cm基于完全随机零模型的分布格局图
2.2 木质藤本的攀附对不同种类和生长特性支持木的选择性
2.2.1木质藤本对不同科属种水平支持木的选择性
支持木在科水平上附藤率有明显差异,紫葳科树木的附藤率最高,而番荔枝科(Annonaceae)的附藤率最低。附藤数量也存在较大变化,紫金牛科的附藤数量最高为649株,葡萄科(Vitaceae)附藤数量仅为2株(表1)。

表1 科的附藤率及附藤数量(附藤率排前10)
在属水平上,附藤率也存在明显差异,最高的东京桐属(Deutzianthus)为最低的澄广花属(Orophea)的10倍。不同属的附藤数量差异也很大,附藤数量最多的紫金牛属超过200株,而蚬木属(Excentrodendron)、火筒树属(Leea)、龙船花属(Ixora)附藤率最低仅为2株(表2)。

表2 属的附藤率及附藤数量(附藤率排前10)
东京桐的附藤率最高达43.75%,平顶紫金牛(Ardisiadepressa)的附藤率最低为2.8%。南方紫金牛的附藤数量最多为137株,平顶紫金牛的附藤数量最少为1株,附藤数量低于5株的有:水东哥(Saurauiatristyla)、蚬木(Excentrodendrontonkinense)、火筒树(Leeaindica)、割舌树(Walsurarobusta)、白花龙船花(Ixorahenryi)、平顶紫金牛(表3)。

表3 树种附藤率及附藤数量(附藤率排前10)
2.2.2木质藤本对不同径级支持木的选择性
样地独立存活的2384株(含分枝共2825株)DBH≥1cm树木中,有431株攀附有木质藤本,每木平均附藤率为15.26%。小树、中树和大树三个径级树木的附藤数量分别为321株、85株、25株,随着树木径级增加,树木被木质藤本攀附的数量虽然降低,但附藤率却增加,且大树的附藤率最高,说明大树更容易被木质藤本攀附(图3)。

图3 树木的径级分布及其附藤百分比
在附藤的树木中,附藤1株的数量最多,共289株,占总附藤树木量的67.05%;附藤3株以下的树木共356株,占总附藤树木的82.60%;附藤最多的树木上攀附有13株藤本。随着每木附藤数的增加,小树、中树的个体数都是先逐渐减少后小幅度增加,大树个体数则是逐渐减少后大幅度增加;此外,小树和中树的附藤数量1株最多,而大树的附藤数量5株以上最多,这说明大树能够容纳更多木质藤本的趋势(图4)。
一元回归分析结果表明,支持木胸径和木质藤本胸径呈极显著正相关(R2=0.1873,P<0.001),随着支持木胸径的增大,木质藤本胸径不断增大的趋势非常明显(图5)。
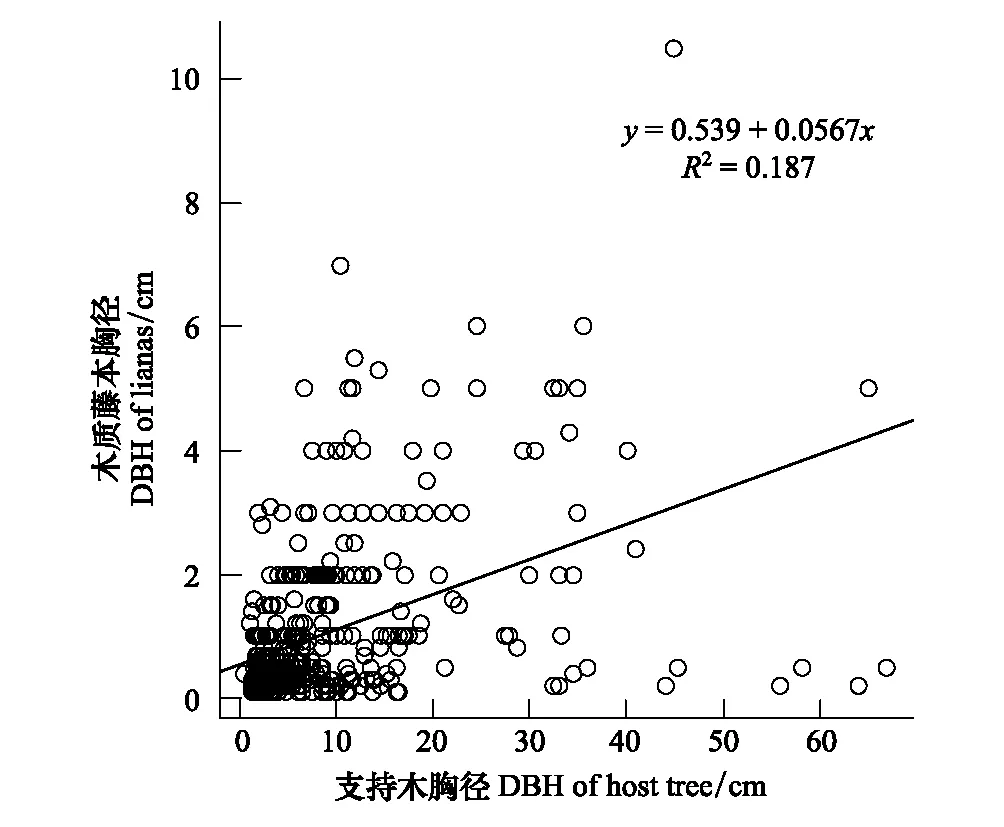
图5 支持木胸径与木质藤本胸径的关系
2.2.3木质藤本对不同聚集强度支持木的选择性
纳入模型分析的22个树种在0—20m尺度的聚集强度呈较大变化,其中苹婆(Sterculiamonosperma)、东京桐在所有空间尺度上均表现为随机分布,对叶榕(Ficushispida)、肥牛树(Cephalomappasinensis)等10个物种在0—20m空间尺度上表现为聚集分布,且随空间尺度的增加聚集强度逐渐降低,典型的3个优势树种的分析结果如图6所示。

图6 五桠果叶木姜子群落样地物种分布散点图、基于完全随机零模型的物种分布格局图
回归模型的分析结果显示,随着物种的聚集强度g0—20(r)值的增加,物种的附藤率显著下降,物种的聚集强度与附藤率呈极显著负相关(F=8.53,P<0.01);随物种聚集强度的增加,物种的附藤数量呈较明显的下降趋势,但物种的聚集强度g0—20(r)与附藤数量的关系不显著(F=2.769,P>0.05)(图7)。

图7 物种的聚集强度与树种的附藤率、附藤数量的一元回归分析
2.3 木质藤本对树木死亡的影响
2.3.1木质藤本密度与树木死亡率的关系
在100个10m×10m小样方中,随着木质藤本密度的增加,树木的死亡率几乎没有发生改变,树木的死亡率与木质藤本的密度的相关关系不显著(F=0.029,P>0.05)(图8)。
2.3.2不同支持木的附藤数量、附藤率与死亡率的多元线性回归分析
不同物种的附藤数量(x1)、附藤率(x2)与树木死亡率(y)的回归方程为y=7.521-0.024x1-0.068x2(R2=0.142,P<0.01),模型在统计学上有意义(F=7.601,P<0.001)。由标准化偏回归系数结果可见,附藤率对死亡率的影响最大。t检验结果表明,附藤率对树木死亡率的影响极显著(P<0.01),且两者为显著的负相关关系,而附藤数量对树木死亡率的相关关系不显著(见表4)。

表4 参数估计及检验
3 讨论与结论
3.1 木质藤本的密度及空间分布特征
喀斯特季节性雨林五桠果叶木姜子群落中木质藤本的平均密度为0.0913株/m2,低于澳大利亚热带雨林(0.1072株/m2)和亚马孙玻利维亚热带森林(0.2471株/m2)[23-24]。样地中木质藤本径级在DBH<3cm占比例高达89.79%,表明群落中小径级木质藤本密度远高于大径级木质藤本的密度。整体来看,五桠果叶木姜子群落大部分生境中木质藤本的密度较低,但局部生境中木质藤本的密度很高,暗示木质藤本的空间格局存在聚集分布特征。
有研究发现,在喀斯特森林藤本植物系统发育结构趋向于聚集[25]。点格局分析结果表明,在0—20m空间尺度内,五桠果叶木姜子样地中的木质藤本整体表现为聚集分布,不同径级的木质藤本在不同空间尺度上的分布格局不同。一般认为,小尺度(0—20m)上的物种聚集分布形成的机制有两种:扩散限制和生态位分化。扩散限制机制认为物种的散布等过程存在距离限制,种子的扩散局限于母株周围,从而导致大部分个体呈现聚集分布;生态位分化则认为物种的分布受生境异质性的影响,不同物种的空间分布对不同生境偏好[26-27],因此会导致一些物种在某些生境中聚集生长。群落中木质藤本的聚集分布应该与这两种机制均有关系。此外,部分原因可能是当树木被一株木质藤本攀附后,其他木质藤本可顺势进行攀附而呈聚集分布,加之种群的聚集分布更有利于抵抗不良环境。不同径级木质藤本在不同空间尺度上的分布格局不同,这可能与群落的自然稀疏过程、环境变化等相关,且种群的空间分布格局可能会随着种群的不断生长发育而出现动态变化。
3.2 木质藤本对树木进行攀附具有选择性
大径级的树木更有可能被木质藤本攀附并容纳较多的木质藤本,这与之前的研究结果一致[14,24,28]。此外木质藤本倾向于选择与自己径级相匹配的树木进行攀附[29]。有研究发现,木质藤本在幼苗时期选择了径级相当的树种进行攀附,到达该支持木的树冠之后就会对相同径级甚至是更高径级树木进行攀附[30]。从另一方面看,大径级树木不仅给已攀附的木质藤本提供便利,而且所占的生存空间更优,即树高更高,树冠更大,木质藤本攀附到树顶后有更多的机会进行光合作用等优势。
有研究表明,木质藤本对支持木在科和种水平都具有选择性[15],这种偏向性选择主要与不同科、树种的径级大小、树冠高度、树皮粗糙度、喜光性、叶片大小等生态习性有关。树种径级大[28]、树冠高大[31]、树皮粗糙[13-14,32]有利于木质藤本进行攀附;耐阴树种一般生长在荫蔽的林下不利于木质藤本的攀附[15],由此推测喜光树种一般生长在光照充足的林冠层而利于木质藤本的攀附;阔叶树种树干分布的木质藤本显著高于针叶树种[33],由此推测叶片大利于木质藤本搭靠。
样地中在科、种水平上附藤率最高的为紫葳科种类和东京桐,紫葳科有木蝴蝶(Oroxylumindicum)、羽叶楸(Stereospermumcolais)、毛叶猫尾木(Markhamiastipulatavar.Kerrii)、菜豆树(Radermacherasinica)、美叶菜豆树(Radermacherafrondosa)5个树种,这5个树种和东京桐都为喜光乔木,一般生长在光照充足的林冠层,冠大荫浓,且东京桐分枝多叶片大,推测方便木质藤本在树冠和枝叶搭靠而不易掉落;虽然这些物种的个体数较少,零散分布于样地,但是紫葳科树种和东京桐大部分个体为中大径级树种,成为木质藤本攀附小径级到中径级支持木、中径级到大径级支持木的攀登梯。番荔枝科和平顶紫金牛在科、种水平的附藤率最低,番荔枝科主要是澄广花属广西澄广花(Oropheapolycarpa),广西澄广花和平顶紫金牛都是径级较小的灌木,分布在光照较弱的林冠下层,且枝条细小而不利于木质藤本的搭靠和攀附。
紫金牛科、紫金牛属、南方紫金牛在科、属、种水平的附藤数量最多。该样地中紫金牛科有杜茎山属中越杜茎山(Maesabalansae),紫金牛属南方紫金牛、紫金牛和平顶紫金牛4个树种;中越杜茎山为多枝灌木,枝条具细条纹增加粗糙度而利于木质藤本的攀附;而南方紫金牛、紫金牛和平顶紫金牛都为灌木或者小乔木,一般生长在荫蔽的林下,因此附藤率都较低。南方紫金牛是紫金牛科数量最多的种,该种径级分布连贯,为不同径级木质藤本进行攀附提供便利,因此该种的附藤数量较多;紫金牛的个体数相对较多,但是紫金牛为不分枝的小灌木或者亚灌木,喜日照不强的林地,不利于木质藤本的攀附;平顶紫金牛附藤率和附藤数量都是最低;同科不同种的附藤情况差异如此之大,这说明除了受树木本身的特征影响外,可能还与其他因素有关,如树种生存环境的差异,也可能与藤本的种类、攀爬习性等相关。附藤数量低于5株的科属种有:楝科割舌树属割舌树、猕猴桃科水东哥属水东哥、葡萄科火筒树属火筒树、茜草科(Rubiaceae)龙船花属白花龙船花、椴树科(Tiliaceae)蚬木属蚬木。水东哥体表被甲状鳞片或钻状刺毛,木质藤本不易在其上攀附生长;火筒树直立的树干不利于木质藤本攀附,而且都是小径级的灌木,在森林中获取光照进行光合的能力较弱;白花龙船花为低矮的常绿灌木,处于光照很弱的森林底层不利于被木质藤本进行攀附。蚬木为常绿乔木,但是平滑的树皮不利于木质藤本攀附,且老时树皮脱落导致木质藤本攀附不稳甚至是跟树皮一起脱落;割舌树虽为喜光树种,但是都为小径级乔木,因此附藤数量较少。当然,木质藤本除受树木本身特征的影响之外,可能还与森林类型、微环境、藤本的分布类型、树木的年龄组成等相关。综上可知,五桠果叶木姜子群落中木质藤本对不同分类群的树木具有选择性攀附的倾向。
五桠果叶木姜子群落中木质藤本对不同聚集强度物种具有偏向性选择。该群落22个主要支持木中,仅苹婆、东京桐在0—20m空间尺度表现为随机分布,其余物种在一定空间尺度内都表现为聚集分布。聚集强度实际代表密度,g(r)值越高表示物种的密度就越大,可能会影响林下的光环境和水环境等生境条件,进而影响木质藤本的生长和发育。如:木质藤本攀附到树冠与树木竞争光资源[34];木质藤本通常能够利用比树木更深层的土壤水,从而与树木竞争地下水分的吸收[35]。广西弄岗喀斯特季节性雨林15hm2监测样地中优势树种强烈聚集分布与生境密切相关[21]。喀斯特地貌中,植物能不同程度地利用各种类型小生境资源[36],植物在其适合的生境内处于竞争优势,光照、水分、土壤养分等非生物因子潜在地影响物种分布[26],而木质藤本作为森林中的重要组分对不同生境也具有偏好。研究发现,哀牢山亚热带常绿阔叶林中木质藤本叶片解剖结构表现出对生境光强具有很强的适应性,而且喜光或适应高强光的木质藤本对异质光生境的适应能力更强[37];在滇南勐宋山地雨林、哀牢山中山湿性常绿阔叶林以及台湾东南部亚热带次生林沟谷中木质藤本的多样性大于坡地[29,38-39];鸡公山落叶阔叶林中藤本植物在树干上的分布受立地坡位的影响,表现为基径处树干坡下位附生的藤本数量高于坡上位,然而在胸高处树干坡下位藤本数量则低于坡上位[33];天目山随着海拔的升高,藤本植物种类下降[40];尼日利亚次生低地热带雨林中土壤变量对藤本植物丰度的影响程度不同[41]。由此可见,不同树种在适宜生境条件下聚集强度越强,密度越大,抵抗不利因素就越强,树种的附藤率、附藤数量就出现越低的趋势,即木质藤本倾向于选择聚集强度较低的树种进行攀附。
3.3 木质藤本对树木死亡的影响
五桠果叶木姜子群落中树木的死亡率与木质藤本密度的相关关系不显著,即随着木质藤本密度的增加,树木的死亡率几乎没有发生改变。从树种水平来看,不同树种的附藤率对死亡率呈极显著负相关,即在一定范围内随着物种附藤率的增加,物种的死亡率减少;而且在一定范围内随着物种附藤数量的增加,物种的死亡率呈下降的趋势。该研究结果部分原因可能是附藤数量多的支持木一般是大径级个体,这样的个体对木质藤本攀附的敏感性和抗性更强,不容易死亡。小径级支持木的枝叶比较少,对环境的适应性没有大径级支持木强,木质藤本攀附可能对它们的光合生理等作用影响更大,更容易死亡。有研究发现在成熟林型会失去被木质藤本牢牢控制的作用[30],且木质藤本对森林生物量的负面影响随着森林年龄的增加而减少,但并未消失[34]。该群落处于保护区核心范围内,原生性保存完好,受人为干扰较少,群落外貌、结构已分化成熟[3]。由此判断,木质藤本对弄岗喀斯特季节性雨林树木的死亡过程可能存在一定程度影响,但并不显著。
4 结论与展望
综上所述,弄岗喀斯特季节性雨林五桠果叶木姜子群落中木质藤本密度较低,以小径级占主体,在0—20m空间尺度表现为聚集分布,随着尺度增大,聚集强度呈下降趋势,不同径级的木质藤本在不同尺度的分布格局不同。木质藤本对不同种类、不同径级、不同聚集强度的支持木具有选择性攀附行为。木质藤本对五桠果叶木姜子群落树木的死亡过程可能存在一定程度的影响,但不显著。对木质藤本及其与树木关系的深入研究,还需对木质藤本的种类、攀援类型、不同树种的生态学特征等进行深入调查,未来将开展长期监测,全面认识喀斯特森林群落的结构、更新动态,从而揭示生物多样性维持机制、阐明生态过程和生态系统功能提升的途径。
