湖南壶瓶山国家级自然保护区鸟类多样性的时空格局
2021-08-30蒋博文贺春容廖庆义康祖杰田书荣
蒋博文,贺春容,廖庆义,康祖杰,文 哲,田书荣
(1.东北林业大学野生动物与自然保护地学院,黑龙江 哈尔滨 150040;2.国家林业局野生动物保护学重点开放实验室,黑龙江 哈尔滨 150040;3.湖南省壶瓶山国家级自然保护区管理局,湖南 常德 415300;4.湖南省林业科学院,湖南 长沙 410004)
鸟类在生物多样性中占据重要地位,是生态系统中不可分割的一部分。它们吞食害虫、给花授粉、散布种子、清除腐肉、循环营养,对于维持生态系统功能起着重要的作用[1-2]。鸟类的物种组成在一定程度上反映了鸟类栖息地环境的综合利用情况。居留型是反映鸟类生态习性与栖息环境关联的有效方式,不同居留型鸟类的物种丰富度和多度在一定程度上随栖息环境的变化有所差别。留鸟活动范围较小,终年生活在出生的区域内,不因季节变化而迁徙;旅鸟在迁徙过程中会途经某一地区停歇和觅食;候鸟通常在一个地方产卵孵化,在另一个地方越冬,受栖息地的安全性、食物的丰富度和可及性影响较大[3]。鸟类物种多样性是鸟类与其他生物及环境相互作用形成的生态组合,它能够表征鸟类群落的组成结构和鸟类栖息环境的优劣,对于了解生态系统的稳定性具有重要意义。自然保护地对生物多样性保护起着重要作用,生物多样性具有多维度性和尺度效应,因此需要从不同的时间和空间尺度进行分析。用多维度、多尺度的视角来研究生物多样性及其变化规律可以更好地了解生物多样性在面对自然和人为驱动因素时如何变化,把握鸟类分布的时空格局,可以为生物多样性的监测和保护提供理论依据[6]。20世纪90年代至今,国际上鸟类研究主要包括鸟类群落的丰富度、均匀度、多样性、密度、多度、相似性、生物量和鸟类群落的季节动态及年动态等方面[7-9]。国内鸟类研究在以上方面也都有了一定的进展,但我国国土面积大,相关研究还需要在部分地区进一步展开[10]。
壶瓶山保护区位于我国云贵高原向东部低山丘陵的过渡地带,是热带植物区系与温带植物区系的重要交汇地带,为武陵地区山脉北支的太清山系山脉所分隔,境内山峰众多,不同地区的海拔高度和生境不同,受第四纪冰川的影响较少。壶瓶山保存了良好的生态环境和大量的古老珍稀濒危孑遗动植物物种及其群落,已成为我国生物的避难所和特有物种的集中分布区,是华中地区生物多样性最丰富和生态环境保护最完好的区域之一[11]。笔者在壶瓶山国家级自然保护区(以下简称“壶瓶山保护区”)设立8个监测站,在监测范围内进行了样线调查,分析壶瓶山保护区鸟类组成及其在不同时间与空间尺度上的动态变化,以期为保护地生物多样性监测和管理提供依据。
1 研究区概况
壶瓶山保护区(北纬29°50′~30°09′,东经110°29′~110°59′)地处武陵山脉东北端,主要位于湖南省石门县境内,总面积66 568 hm2(图1),其北部核心区紧邻湖北后河国家级自然保护区。
境内山峰连绵,多数为单面峻脊,地势较为陡峭。最低海拔220 m,最高海拔2 098.7 m。属亚热带山地气候区,受太平洋暖流的影响明显,年均温度9.2 ℃,年均降水量1 898.5 mm。壶瓶山保护区森林植被是目前我国亚热带地区少数保存较完好的地区之一,森林覆盖率达86.5%,植被覆盖度达98.7%,森林资源类型主要包括低山针叶林、中山针叶林、中山针阔混交林、常绿阔叶林、常绿落叶阔叶混交林、落叶阔叶林等。壶瓶山国家级自然保护区现已记录179种鸟类,共15目39科。其中金雕(Aquilachrysaetos)属国家Ⅰ级重点保护动物,国家Ⅱ级保护鸟类有34种,占整个保护区内已知鸟类物种数的18.99%[11]。
野外调查在8个常设监测站点监测范围内进行(图1)。将8个站点的监测范围根据海拔划分为5个梯度:≤1 000、>1 000~1 200、>1 200~1 400、>1 400~1 600、>1 600 m。其中≤1 000 m海拔段包括土湾站点,>1 000~1 200 m海拔段包括神景洞和惜粮坪站点,>1 200~1 400 m海拔段包括野猫岔和黄莲棚站点,>1 400~1 600 m海拔段包括壶瓶山站点,>1 600 m包括顶坪和联防站点。
2 研究方法
2.1 数据收集
2016年1月至2017年12月在壶瓶山保护区8个监测站点的监测范围内采用固定样线法进行鸟类调查[12]。结合各监测站点的植被和地形特点,在每个监测站点布设2~4条沿山脉走向的固定样线(表1),共计23条,平均海拔1 418.95 m,样线单侧宽25 m,平均长度7 km(5~9 km),各样线间的平均距离为1 km。每条样线每个月进行1~4次调查,核心区西组4个站点的样线基本呈南北走向,核心区东组的2个站点及黄莲棚站点的样线基本呈东西走向,实验区土湾站点的样线基本呈西南走向,各样线间没有交叉或重复。为减少调查次数及样线长度不同带来的影响,每个站点每月鸟类调查数据求和后,分别除以相应站点当月总调查次数和样线总长度,以对数据进行标准化处理。
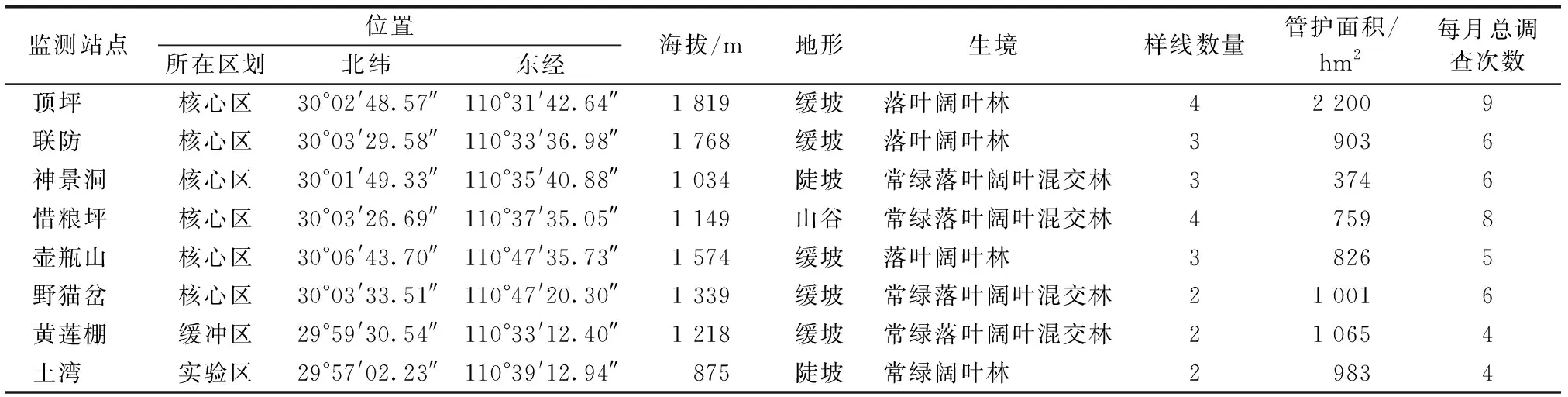
表1 各监测点所在位置基本生境和样线信息
每次调查的平均速度为2 km·h-1,根据气象条件在调查日的06:30—09:30或14:30—17:30进行。采用双筒望远镜(蔡司8×30BT)进行观测;采用索尼ICD-SI813录音笔记录鸟类鸣叫声;采用尼康D90、尼康D800带400 mm长焦距镜头进行拍摄;鸟种鉴别参照文献[13],居留型、分布型和保护级别参照文献[14] ,记录样带两侧鸟类的种类、数量以及与样带中心线的垂直距离。用GPS(Garmin 60csx)记录行进速度、样线长度以及起点和终点的经纬度、海拔。
2.2 数据处理与分析
2.2.1物种相对丰富度
分别计算8个监测站点各物种的相对丰度(relative species abundance,AR)[15]:
AR=Ni/N。
(1)
式(1)中,Ni为第i个物种在某监测站点的数量;N为某监测站点鸟类总个体数。得到各物种的相对丰度后,按科分别将2 a的物种相对丰度数据以求和的方式合并,获得各站点监测范围内以科为单位的相对丰度数值,从科阶元的尺度剖析鸟类组成情况。
2.2.2α多样性
采用香农-威纳多样性指数(Shannon-Wiener index,H′)[16]和Pielou均匀度指数(Pielou′s evenness index,J′)[17],以种为单元衡量各站点监测范围的α多样性:
(2)
J′=H′/lnS。
(3)
式(2)~(3)中,pi为第i个物种的个体数与物种个体总数的比值;S为总物种数。
以标准化后的调查数据为基础,按月份分别计算每个站点范围内的α多样性指数来比较α多样性在不同时间尺度上的变化。采用方差分析(ANOVA, TukeyHSD)分别比较α多样性指数在不同站点、不同海拔梯度、不同年份、不同季节和不同月份间是否存在显著差异,然后用t检验计算站点间的差异显著性。
2.2.3β多样性
采用惠特克指数(Whittaker′s index)按月计算整个壶瓶山保护区的鸟类群落多样性,以衡量β多样性指数[18]。以标准化的数据为基础,使用R语言环境中Vegetarian包[19]的d函数计算各月份和各季节的惠特克指数。以方差分析来比较β多样性指数在不同时间是否存在显著差异。
2.2.4监测站点间物种组成的相异性
采用Bray-Curtis相异性矩阵比较不同站点间物种组成的差异[18]。将每个监测站点2 a的调查数据按季节以求和的方式合并,使用R语言环境中Vegan包[20]的Vegdist函数获得站点间的Bray-Curtis相异性矩阵。为了消除数据中零值的影响,将得到的相异系数分别对数转化后加一。采用方差分析比较Bray-Curtis相异系数在不同海拔段间是否存在显著差异。所有的计算都在R Version 4.0.2软件下完成,统计检验显著性水平为 0.05。
3 结果与分析
3.1 鸟类组成
2016—2017年共记录鸟类110种,隶属13目37科(表2),调查到的物种数占整个湖南省鸟类资源总数的24.66%。

表2 调查到的鸟类科组成及其相对丰度
其中中华秋沙鸭(Mergussquamatus)和金雕(Aquilachrysaetos)为国家Ⅰ级保护物种,而国家Ⅱ级保护物种包括红腹角雉(Tragopantemminckii)、勺鸡(Pucrasiamacrolopha)、红腹锦鸡(Chrysolophuspictus)、蛇雕(Spilornischeela)、凤头鹰(Accipitertrivirgatus)、白腹鹞(Circusspilonotus)、游隼(Falcoperegrinus)、红翅绿鸠(Treronsieboldii)、红角鸮(Otussunia)、灰林鸮(Strixnivicolum)、领鸺鹠(Glaucidiumbrodiei)、斑头鸺鹠(Glaucidiumcuculoides)、鹰鸮(Ninoxscutulata)、长耳鸮(Asiootus)。
从科水平的分布来看,雉科(Phasianidae)、鹰科(Accipitridae)、鸠鸽科(Columbidae)、鸦科(Corvidae)、噪鹛科(Leiothrichidae)在所有站点均有分布,其中雉科和噪鹛科丰度较高,且雉科在神景洞、惜粮坪、黄莲棚站点为优势科,而噪鹛科在顶坪、联防和野猫岔站点为优势科。与相毗邻的湖北后河国家级保护区相比,壶瓶山保护区丰度较高的除了噪鹛科和鹰科外还有雉科、鸠鸽科和鸦科[21]。水鸟只在顶坪和土湾站点有记录,且土湾站点的鸭科(Anatidae)丰度较高。从居留型来看,这110种记录到的鸟类中,留鸟(R)共计81种,占所记录鸟类的73.64%;夏候鸟(S)共计14种,占所记录鸟类的12.73%;冬候鸟(W)共计11种,占所记录鸟类的10.00%;旅鸟(P)共计4种,占所记录鸟类的3.64%。可见壶瓶山保护区鸟类以留鸟为主,其次是夏候鸟、冬候鸟,旅鸟较少。从分布型来看,在这83种留鸟中,东洋种鸟类最多,共计48种,占59.26%;广布种次之,共计26种,占32.10%;古北种最少,共计7种,占8.64%。可见鸟类区系组成以东洋界种类占明显优势,这一结果与保护区动物区系属东洋界华中区西部山地高原亚区的特点相一致。
3.2 鸟类多样性特征
由于年际间鸟类群落α多样性没有显著差异,后续分析中将2 a的数据合并。各站点监测范围内α多样性指数存在显著差异(图2)。与顶坪站点相比,联防和野猫岔站点的α多样性水平显著较低,而神景洞、惜粮坪和土湾站点显著较高。各海拔梯度间α多样性存在显著差异(P<0.05)。与最高海拔段相比,其他海拔段的α多样性显著较低,最低海拔段的多样性也最低,基本上表现出随海拔单调递增的趋势。α多样性在月际间没有显著差异,但2月显著低于3月(图3),而在季节间也有显著差异,春季较高。整个壶瓶山保护区鸟类群落的β多样性指数在年际间、月际间均没有显著差异。但在季节间存在显著差异,夏季高于其他季节(图4)。通过相异性矩阵的Bray-Curtis相异系数比较各监测站点物种组成的差异,其值越大表示站点间差异越大,结果显示各海拔段在季节上的物种组成没有显著差异(图5)。
4 讨论
4.1 壶瓶山保护区鸟类多样性特点
壶瓶山保护区鸟类的组成、区系和居留型结果与湖南省其他地区以及湖北省后河保护区结果相似[21-24]。东洋界处于热带与亚热带地区,其物种主要由华中区与华南区共有种以及华中区物种组成,是古北界东部候鸟的越冬地。鸟类飞翔能力强,存在迁徙行为,故鸟类区系表现出东洋界物种和古北界物种相混杂的现象。
鸫科鸟类在大多数站点中为优势物种(表2),这是因为鸫科行动能力较强而且活动较频繁,被发现的概率较高,可见鸟类的行动能力对于生物多样性监测和评估有直接影响[25]。壶瓶山保护区境内峰峦叠嶂,雨量充足,地貌形态丰富多样,各区域均存在适合水鸟生活的环境。然而与以往的调查相比,这2 a调查到的水鸟较少[11],仅在实验区的土湾站点记录到丰度较高的鸭科鸟类,说明保护区对缓冲区与核心区的生境管理有待加强,而对实验区的管理有效,需继续保持。
4.2 鸟类多样性的时空格局
由于鸟类对生境的使用方式和适应能力不同,保护区各区域和各海拔段的生物多样性水平高低不等,各监测站点鸟类组成存在显著区别(表2)。保护区工作人员的日常巡护监测在一定程度上影响着生物多样性水平[5]。α多样性是认识群落物种数、个体数和分布均匀程度的综合指标,可以揭示群落内物种丰富程度,并为群落环境评价提供依据。壶瓶山保护区不同区域的生物多样性水平有显著差异,从α多样性指数看,位于保护区核心区西侧的顶坪和联防站点鸟类多样性水平较高,其次是位于核心区东侧的野猫岔站点,这3个站点的α多样性指数显著高于缓冲区的黄莲棚站点和实验区的土湾站点(图2)。壶瓶山保护区北侧紧邻湖北后河国家级自然保护区,顶坪、联防和野猫岔站点这3个站点的监测范围既在壶瓶山保护区的核心区,又邻近后河保护区,保护水平较高。相反,土湾的监测范围位于实验区,人类干扰相对比较剧烈,α多样性指数均显著较低。这些结果充分反映了保护水平对生物多样性的影响[26]。
从垂直角度来看,各海拔段之间的物种组成相异性与季节没有显著关系(图5)。最高海拔段多样性水平显著高于其他站点,最低海拔段多样性水平也最低。壶瓶山保护区属于亚热带中低海拔地区,鸟类多样性基本呈随海拔递增的趋势,符合物种垂直分布结构中的单调递增模式[27]。而且,处于最高海拔段的顶坪和联防站点的生境均属落叶阔叶林,雨量丰富,温度适宜,林下植被盖度很高,茂盛的草本植物为鸟类提供了丰富的食物资源,故鸟类数量和种类较多。此外,Bray-Curtis相异系数说明,不论季节如何,每个海拔段都有相对稳定的鸟类群落组成。
动物对生境的选择和利用是对栖息地环境的一种适应性行为。鸟类除了要选择合适的海拔段外,还需要选择温度适宜的地区,因此它们会在相对固定的时间开始迁飞。壶瓶山保护区处于中部候鸟迁徙区。鸟类多样性在3月达到最高值之后开始降低,这是因为夏候鸟开始往更偏北的地方迁飞,寻找更适宜的繁殖地。而在11月,鸟类多样性出现了第2个高峰之后又接着降低,因为11月正值冬候鸟越冬迁飞时节,在北部繁殖的候鸟会在迁徙期途经华中地区往更偏南的地区越冬。从月际变化基本可以推断出壶瓶山保护区鸟类迁徙的动向。
从季节上来看,α多样性结果显示,各站点春季鸟类多样性水平均较高而冬季较低,这是由于春季气候刚变暖,树木刚开始发芽,这些条件为鸟类营造了更多的筑巢机会;植被郁闭度较低,有许多幼虫出生,鱼类和两栖类增加,使鸟类有了充足的取食机会,吸引了大量鸟类来此觅食。β多样性体现的是群落组成变化的程度,不仅可以反映整个保护区各群落间的多样性距离关系,还可以反映群落之间的物种分化程度。整个保护区的β多样性在年际和月际上都没有显著差异,但是各站点鸟类α多样性在月际上存在显著差异,一定程度上说明了保护区内各监测站点的鸟类数量变化较小,导致整个保护区每个月的物种多样性变化没有明显差别,各海拔段Bray-Curtis相异性在季节上没有显著差异。夏季鸟类β多样性水平相比其他季节要高,因为保护区夏季多雨潮湿且植被郁闭度高,适合不同昆虫增殖,吸引了更多夏候鸟来此繁殖,在某种程度上增加了鸟类的数量。而冬季不论是α多样性还是β多样性均较低,这是因为冬季没有充足的食物,且温度较低,部分鸟类无法适应,需要迁徙到更暖和的地方过冬[28-29]。此外,季节内也存在月份间差异,2月比12月和1月多样性更低,这是因为可供取食的食物逐渐消耗,从而出现一个“饥饿间隔”,直到3月春季来临食物量开始增加,鸟类多样性突然增长[30-31]。
4.3 监测与保护建议
生物多样性监测是生物多样性保护中非常重要的基础性工作[32],但是监测工作也要消耗大量的资源,如何有效利用资源,提高监测效率,是生物多样性保护工作者需要思考的问题。笔者建议保护区各站点统一样线数量、调查次数及样线长度,以减少统计误差;人类社会活动对鸟类有或多或少的影响,应该在调查中加入人为干扰强度的测评,限制保护区内的车流与人类活动;利用爱鸟周活动,加大鸟类资源保护宣传力度,提高全民爱鸟、护鸟意识;加强河流湿地生态系统的保护,为迁徙鸟类和越冬水鸟以及提供一个良好的栖息生活环境;人工改造或恢复局部区域的生境,为部分分布区域狭窄的鸟类繁衍提供适宜的生境;加强监测人员培训,及时整理和备份数据,进一步规范监测程序,缩小人为影响因素对监测分析结果造成的误差。
