香稻Badh2 基因单倍型及香气成分2-乙酰-1-吡咯啉代谢通路的研究进展
2021-08-25潘阳阳黄道强王重荣周德贵王志东陈宜波周少川
潘阳阳,黄道强,王重荣,李 宏,周德贵,王志东,陈宜波,赵 雷,龚 蓉,周少川
(广东省农业科学院水稻研究所/广东省水稻育种新技术重点实验室,广东 广州 510640)
香米在蒸煮过程中会散发出令人愉悦的独特香味,被誉为水稻中的珍品,具有较高的经济价值。而稻米香味是国际市场上决定香米价格的重要因素之一[1],优质香米价格是非香稻的3 倍之多[2]。传统香稻品种具有很强的地域特性,泰国的茉莉香米、印度与巴基斯坦的巴斯马蒂香米长期占据国际香米市场主要份额,近年来越南、柬埔寨等国的优质香米出口量也在不断加大。我国作为全球最大的稻米生产国和消费国[3],市场供需不平衡,每年需进口一定数量的优质香米以满足国内高端市场需求。因此,加强香稻品种选育,打造我国香米品牌以取代泰国香米,对于提升我国香米国际影响力具有重要的经济和社会效益。
我国具有丰富的香稻资源,香稻种植历史可以上溯到3 000 多年前。由于香稻多为地方性品种,兼受产量、抗性和改良技术等条件限制,导致我国香稻生产长期不被重视[4-5]。当前,我国稻米消费市场面临转型升级,优质且具有香味的大米深受市场青睐,水稻育种由注重产量向产量品质并重的方向转型。2018—2020 年,全国农业技术推广服务中心连续3 年举办全国优质稻品种食味品质鉴评会,有力推动我国水稻育种向优质稻育种方向转型。
香味物质的挖掘和香味基因的遗传调控是香稻育种的两个根本问题,长期以来备受关注。前人在稻米中发现了上百种挥发性香味化合物[6-9],随着检测技术水平的提高和研究的不断深入,1983 年2-乙酰-1-吡咯啉(2-acetyl-1-pyrroline,2AP)被确认为稻米特征香气成分[10]。水稻香味基因的遗传研究起始于20 世纪70 年代,早期研究推断香味基因可能由单个[11]、2 个[12]或3 个[13]隐性基因控制。随着分子标记技术和水稻功能基因组学的发展,Ahn 等[14]利用分子标记技术将香味基因定位在第8 号染色体上,Bradbury 等[15]图位克隆到香味主效基因Badh2(LOC_Os08g32870),该基因第7 外显子8 bp 的缺失和3 个碱基的突变致使基因功能失活,最终引起2AP 积累。
随着基因组测序技术快速发展,研究者们在香稻资源中挖掘出了更多功能性突变的Badh2单倍型[16-17],这为香稻育种提供了丰富的香稻基因资源;同时,对香味基因的生化基础和2AP 生物合成途径开展了细致研究[18-19],初步揭示了2AP 的生物合成通路[20]。但香味遗传性状十分复杂,不同品种稻米的2AP 含量存在明显差异、相同品种在不同种植条件下的2AP 含量也不相同[2,21-23],而如何特异提高精米2AP 含量也是香米产业发展面临的关键问题[1],这些问题的解决需要进一步解析2AP 生物合成机制,尤其要深入认识籽粒发育过程中2AP 积累机制。本文对已发现的香稻Badh2基因单倍型及不同单倍型香稻中2AP 含量等的研究进行综述,重点阐述2AP 生物合成通路的研究进展,提出从籽粒发育动态角度研究2AP 积累机制,为香稻资源相关基因利用、籽粒特异富集2AP 的香稻品种研发提供依据。
1 香稻Badh2 基因的单倍型
1.1 香味基因Badh2 的功能研究及利用
水稻Badh2基因编码甜菜碱醛脱氢酶,该蛋白能够催化甜菜碱醛合成甜菜碱,从而在抗盐害、冷害和热害等逆境胁迫中发挥重要作用[15,24]。研究发现,水稻中存在1 个BADH2 同源蛋白BADH1,且两个蛋白功能可能发生了明显分化:Fitzgerald 等[25]发现非香稻中这两个蛋白均发挥功能,香稻和非香稻品种中BADH1 的转录本并不存在差异,BADH1 在响应盐胁迫中发挥主要作用;He 等[26]发现BADH1 与苗期水稻耐盐性显著相关,而BADH2 只与稻米香味性状密切关联;BADH2 在pH 10 的碱性条件下具有最大催化活性,催化底物主要为4-氨基丁醛,而BADH1 在pH 9.5 条件下具有催化活性,且对4-氨基丁醛催化活性极低[18]。
表达模式分析表明,Badh2在水稻的地上部分均能表达,亚细胞定位发现BADH2 蛋白主要定位于细胞质[19]。Baicharoen 等[27]深入解析了BADH2 蛋白的3D 结构,发现该蛋白包含NAD+结合域、底物结合域和寡聚化结合域,揭示了关键氨基酸残基如N162、C294、E260 在催化反应中的关键作用,为优质香稻改良提供了理论基础。
随着分子标记辅助育种、基因编辑育种等技术的发展,通过RNAi、TALEN 等生物技术对Badh2进行敲减或敲除,已成功获得转基因香稻材料[28-30],随着CRISPR/Cas9 基因编辑系统在作物育种中的广泛应用,该技术已成为快速获得香稻材料的重要方法[31-32]。
1.2 香稻资源中Badh2 单倍型的挖掘
基于转基因存在的潜在风险挑战,挖掘更多自然变异的香味基因单倍型对于丰富优质香稻育种多样性具有重要意义。通过对香稻资源的序列分析,目前在Badh2基因编码区共发现15 种功能性突变的单倍型(表1),这些变化的核苷酸位点分布在第1、2、4、7、8、10、12、13、14外显子,以小片段序列插入或删除(Indel)和单核苷酸变异(SNP)为主[16-17,33-35]。
从Badh2基因型分布和基因频率分析,badh2-E7(第7 外显子8 bp 的缺失和3 个碱基的突变)是最常见的突变类型,广泛分布在世界主要稻米生产国,如著名香籼稻KDML105 和Basmati370、美国香粳稻Della、我国优质稻花香2 号、广东丝苗米代表品种美香占2 号和象牙香占等均为这种单倍型,表明在较早的香稻育种过程中,这种单倍型被育种家无意识选择并广泛应用。其余14 种单倍型具有较强的地域分布特性,相应的香稻品种数量较少,且多在粳稻中被发现,如江苏的南粳系列均为badh2-E2.1单倍型,浙江和广西的香糯稻为badh2-E4-5单倍型,这些香味基因型可能受特殊地理限制,形成了特色香稻品种。此外,多项研究发现Badh2基因启动子区域变异也能够导致2AP 积累(表1),目前共发现4种单倍型[36-39],主要表现为Badh2基因5′ UTR相应序列缺失。分子功能研究表明,这些材料中2AP 的积累是由于BADH2 蛋白含量显著下降引起的。
充分利用香稻资源中丰富多样的Badh2单倍型进行常规杂交育种,一方面能规避转基因风险,另一方面可能创制出不同香味类型的香稻品种。当前,对大多数Badh2单倍型已开发出特定的分子标记,不仅可以用于筛选香稻基因型,而且在香稻的分子辅助育种进程中发挥重要作用[34-35]。
1.3 不同Badh2 单倍型香稻的2AP 含量分布特征
不同Badh2突变类型材料中的2AP 含量不同(表1),BADH2 蛋白活性是影响2AP 积累的重要因素之一。通过对Badh2基因启动子变异的香稻材料、RNAi 转基因香稻材料的分析,发现BADH2 蛋白水平显著降低[28,40];Kovach等[16]研究发现,发生1 个或2 个碱基插入或缺失的单倍型材料(如badh2-E1、badh2-E10.1、badh2-E14.1)的2AP 含量整体高于单碱基替换或3 bp 插入的单倍型材料(如badh2-E10.3、badh2-E13.1、badh2-E14.2),表明2AP 含量与BADH2 蛋白活性密切相关,但对于BADH2 蛋白活性下降到何种程度才能导致2AP 积累这个问题尚未研究。另外,相同Badh2 单倍型材料之间,2AP 含量仍然存在较大差异,这与遗传背景不同直接相关。
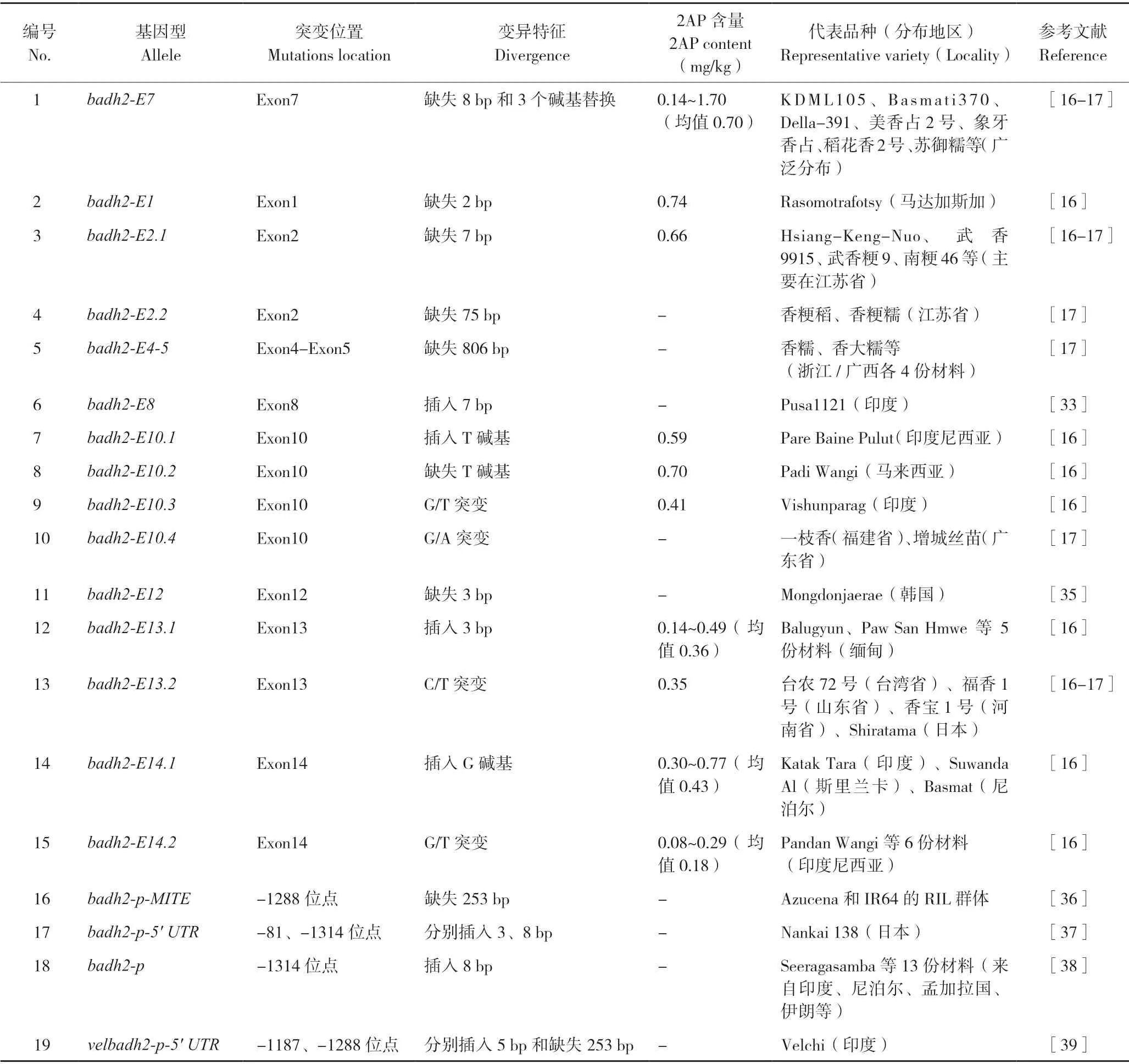
表1 香稻Badh2 基因突变类型Table 1 Type of alleles of Badh2 in aromcotic rice
目前,研究Badh2单倍型与2AP 含量关系最大的障碍在于缺乏遗传背景相同、Badh2变异类型不同的遗传材料,而利用CRISPR/Cas9 对非香稻Badh2的不同外显子、启动子区段设计靶点,获得不同变异类型的香稻材料,能有效解释Badh2单倍型与2AP 含量之间的关系,同时也是对高积累2AP 香稻创制的有益探索。
2 2AP 生物合成通路及其在香稻籽粒中的分布特征
2.1 2AP 代谢通路相关研究
不同香稻品种、不同栽培条件均导致2AP 含量差异,这与2AP 生物合成通路相关代谢物的变化密切相关。BADH2 能够催化4-氨基丁醛氧化,从而生成4-氨基丁酸,而香味基因失活时,4-氨基丁醛能够直接环化生成1-吡咯啉,1-吡咯啉再与乙酰基团结合,无需酶促反应即可合成2AP[41-42],研究发现,1-吡咯啉是2AP 合成中重要的限制性底物[43]。根据1-吡咯啉的来源不同,2AP 生物合成途径主要分为谷氨酸-脯氨酸代谢和多胺代谢两种途径[18,20,44],如图1 所示,谷氨酸和脯氨酸分别在脯氨酸脱氢酶(ProDH)和1-吡咯啉-5-羧酸合成酶(P5CS)的作用下,生成中间产物1-吡咯啉-5-羧酸(P5C),P5C 继而转化为1-吡咯啉;多胺代谢能够生成中间产物4-氨基丁醛,4-氨基丁醛在BADH2 酶失活的情况下环化为1-吡咯啉。

图1 2AP 生物合成通路示意图Fig.1 Schematic diagram of the 2AP biosynthetic pathway
当前,对谷氨酸-脯氨酸代谢途径的研究主要集中在脯氨酸含量以及P5CS、ProDH两个关键基因表达变化对2AP 积累的影响方面。Huang等[45]研究发现,香稻中P5CS 酶活显著高于非香稻;在Badh2启动子区段变异的香稻“Velchi”中脯氨酸含量、P5CS 转录水平和酶活均高于非香稻[39];在香稻中过表达P5CS,籽粒中2AP 含量可提高2 倍[46]。同时,增香栽培研究大多关注谷氨酸-脯氨酸通路对2AP 含量的影响:Mo 等[47]对香稻遮光处理后,脯氨酸含量显著提高,2AP含量提高59%;Bao 等[48]对抽穗期香稻进行水分干湿灌溉处理,相关籽粒中ProDH、P5CS 和二胺氧化酶(DAO)活性显著提高,2AP 含量提高了45%;适当的水氮调控及锌、硒等处理[49-51]均可提高谷氨酸-脯氨酸代谢途径活性从而提高2AP 含量。但目前的关键问题在于尚无直接证据证实植物体内P5C 能够转化为1-吡咯啉,从而制约了对籽粒2AP 含量的精准调控。
多胺在植物生长和发育过程中起重要的调控作用[52-53],同时在多种逆境胁迫中发挥重要作用,如冷害、热害、盐胁迫和病虫害等[54-57];已有研究表明,多胺参与水稻籽粒灌浆的调控,对籽粒充实度、稻米品质、种子活力等均有影响[58-60],腐胺、精胺和亚精胺等多胺物质经酶催化最终能够生成4-氨基丁醛[61-62],但多胺物质与籽粒2AP 积累的相关性研究未见报道。因此,加强香稻籽粒发育过程中多胺代谢对2AP 含量的影响机制研究,挖掘多胺途径中调控2AP 含量的关键代谢物和关键基因,将有助于实现籽粒2AP 含量的定向提高。
2.2 香稻籽粒中2AP 分布特征及积累机制研究
精米中2AP 含量一般明显低于糙米,但不同品种籽粒精米中2AP 分布特征差异较大。Buttery等[63]发现巴斯马蒂香米、KDML105 籽粒中约35%的2AP 分布于精米;应兴华等[64]研究发现桂香丝糯、清香米、泰国香稻1 号R207 等5个香稻品种精米中2AP 含量占比在90%以上。Yoshihashi 等[65]推断精米中2AP 与淀粉颗粒复合体紧密结合,但籽粒中2AP 积累机制尚不明确,目前提出两种机制加以解释:一种认为叶片和鞘中合成的2AP 被转运至籽粒,另一种认为2AP 在籽粒中被从头合成。研究胚乳发育过程中2AP 代谢通路变化特性,对于认识籽粒2AP 积累机制、培育2AP 高积累水稻至关重要。Hinge 等[20]研究发现,随着籽粒成熟2AP 含量不断提高,但籽粒中脯氨酸含量较低,P5CS基因表达水平下降,从而认为籽粒中2AP 是由叶片转运而来,但该研究未考虑多胺代谢对2AP 的影响;潘阳阳等[66]利用代谢组和转录组联合分析技术,对美香占2号乳熟期、蜡熟期和完熟期的籽粒2AP 代谢通路进行分析,发现3 个时期籽粒中均含有2AP 生物合成所需的前体物质,同时,P5CS、ProDH以及多胺代谢途径中二胺氧化酶4(DAO4)、多胺氧化酶4(PAO4)、精胺合成酶和亚精胺合成酶等相关基因具有持续较高的表达水平,推测谷氨酸-脯氨酸转化通路和多胺代谢途径均有助于2AP 的积累。因此,从籽粒动态发育角度研究影响2AP积累的相关代谢物和相关基因具有较高可行性,将成为揭示2AP 积累机制的重要手段。进一步加强胚乳中特异高表达的DAO4、PAO4等基因对2AP 含量影响的研究,对于特异提高精米中2AP含量具有重要意义。
3 展望
随着水稻功能基因组学的发展,香稻资源中Badh2基因单倍型得以深入挖掘,不同单倍型香稻材料之间2AP 含量存在较大差异;2AP 生物合成通路也被初步解析,但其中多数基因的生物学功能尚未研究,对2AP 如何在籽粒中积累这一关键问题仍然缺乏系统认识。利用当前新兴的代谢组、转录组、蛋白质组等多层组学技术,综合研究香稻籽粒发育过程中2AP 相关代谢物和基因的变化特性,尤其是重点关注多胺代谢途径的相关变化,挖掘关键调控基因,将有助于认识籽粒2AP 积累机制。因此,未来香稻的研究可以聚焦以下两个方面:(1)解析不同香稻类型2AP 代谢通路的差异,重点解析多胺代谢对2AP 含量的影响机制;(2)加强不同类型Badh2单倍型香稻资源中DAO4、PAO4等基因的利用,培育2AP高积累的香稻品种。
