巴克夏猪、淮猪及其杂交F1代HAL基因多态性检测
2021-08-24凌清露靳二辉顾有方
凌清露, 李 慧, 陈 苗, 靳二辉, 顾有方
(安徽科技学院 动物科学学院 动物营养调控与健康安徽省重点实验室,安徽 凤阳 233100)
氟烷基因(Halothane,HAL
)又称氟烷敏感基因或兰尼定受体基因,它是猪产生应激综合症和PSE(Pale Soft Exudative)肉的主效基因。Davies等在猪第6号染色体pq上定位氟烷基因,由等位基因HAL
(正常显性)和HAL
(变异隐性)进行控制。Fujii等研究了皮特兰猪和大白猪的兰尼定受体cDNA序列表明该基因的cDNA中发生C→T突变,使Arg置换为Cys时,猪易产生应激综合征。Fisher等研究表明HAL
HAL
型猪日增重显著高于HAL
HAL
型和HAL
HAL
型。帅素容等研究表明HAL
HAL
型肉质最好,HAL
HAL
型肉质最差,HAL
HAL
型肉质在两者之间。刘显军等研究表明HAL
HAL
猪胴体重、眼肌面积、瘦肉率显著高于HAL
HAL
猪。张建军研究表明携带HAL
HAL
型和HAL
HAL
型基因的猪对应激敏感,其肌肉收缩强烈,肌肉的生长和脂肪的消耗速度比正常猪快,从而提高了瘦肉率。在对繁殖性状的影响上,携带有HAL
HAL
基因的母猪对应激敏感度高,母猪受到应激后难以配种,怀孕母猪极易流产。总之,隐性纯合子HAL
HAL
型相对于HAL
HAL
型和HAL
HAL
型表现出了一定的劣势。有关于基因分型技术,目前仍广泛利用聚合酶链式反应限制性片段长度多态性技术(PCRRFLP)检测氟烷基因,而PCR-RFLP在该领域尚未见报道。外种猪与中国地方猪杂交的后代普遍具有能遗传两个亲本优势的能力,具有较好的杂交优势。巴克夏猪源于英国,盛产雪花瘦肉,是一种肉质优良的瘦肉型猪种。淮猪具有繁殖力强、耐粗饲、抗病性能好等特点。将引进的巴克夏猪选育成优质瘦肉型专门化父系,利用淮猪(霍寿黑猪)优良肉质基因资源,导入巴克夏猪血统,杂交而得F代巴淮猪,具有较好的杂交优势,更适合国人对肉质的需求。
本研究采用PCR-RFLP技术检测了巴克夏猪、淮猪(霍寿黑猪)、巴淮猪HAL基因型分布,为巴克夏猪、淮猪(霍寿黑猪)的开发利用以及加速育成肉质优良的巴淮猪提供参考依据。
1 材料与方法
1.1 材料
1.1.1 实验动物 来自安徽浩翔农牧有限公司养猪场的巴淮猪105头、巴克夏猪32头,来自凤台县顺民养殖有限公司的淮猪(霍寿黑猪)38头,采集上述猪耳组织样本于75%乙醇溶液中,-20 ℃保存。
1.1.2 主要试剂与仪器 蛋白酶K、2×Taq PCR Master Mix(含染料)、DNA Marker DL2000,购于天根生化科技(北京)有限公司;限制性内切酶(Hha I)购自纽英伦生物技术(北京)有限公司;Tanon凝胶成像系统、稳压电泳仪、平板电泳槽、PCR仪。
1.2 方法
1.2.1 DNA的提取及检测 参考《分子克隆实验指南》,用苯酚氯仿抽提法提取基因组DNA,取耳组织0.5 g装在1.5 mL离心管中,加入600 μL组织 DNA提取液。在1.5 mL离心管中加入10 μL Proteinase K(20 mg/mL),摇晃充分均匀后置于56 ℃的恒温水浴锅中消化过夜。水浴结束后,向各个1.5 mL离心管中加入约600 μL Tris饱和酚,上下翻转混匀约8 min,12 000 r/min离心8 min。取上步骤的清液,各加入300 mL Tris饱和酚、288 μL氯仿、12 μL异戊醇,上下翻转摇晃混匀8 min,12 000 r/min离心8 min。将离心后的上清液吸入干净的1.5 mL离心管中,加入576 μL氯仿和24 μL异戊醇,上下翻转混匀8 min,12 000 r/min离心8 min。取离心后上清液,加入1 mL冰乙醇,此时水平摇动,可见絮状、白色凝集物团状DNA出现。12 000 rpm离心5 min,DNA沉于管底,小心倒去冰乙醇。加入600 mL 75%乙醇,摇动洗涤DNA,12 000 r/min离心3 min后弃废液,重复步骤摇动洗涤3次。弃去75%的乙醇,斜放自然干燥,等待酒精挥发完全后,加入150 μL TE溶解DNA。将样本分装3份,50 μL/份。留取一份稀释,另外两份于-20 ℃保存备用。根据核酸蛋白仪检测的DNA浓度,按照比例将DNA样品稀释到100~200 ng/μL,于4 ℃保存待用。
1.2.2 引物设计与合成 参照Fujii等的引物,猪HAL
基因上游引物序列5′-TCCAGTTTGCCACAGGTCCTACCA-3′,下游引物序列5′-ATTCACCGGAGTGGAGTCTCTGAG-3′,由生工生物工程(上海)股份有限公司合成。1.2.3HAL
基因的PCR扩增 PCR反应体系(25 μL)为:2×Taq PCR Master Mix 12.5 μL,上下游引物各1 μL,DNA模板1 μL,双蒸水10.5 μL。PCR扩增条件为:95 ℃预变性,5 min;94 ℃变性35 s,61 ℃退火35 s,72 ℃延伸35 s,共35个循环;72 ℃终延伸10 min,4 ℃保存。扩增产物于2%的琼脂糖凝胶电泳进行检测。1.2.4 PCR产物酶切 采用限制性内切酶Hha I对扩增的猪HAL
基因目的片段进行酶切,反应体系(10 μL)为:PCR产物5 μL,内切酶0.5 μL,内切酶缓冲液1.5 μL,双蒸水3 μL。酶切反应温度为37 ℃,酶切产物于3%的琼脂糖凝胶电泳检测,判断基因型。1.2.5 数据分析 利用χ检验判断群体中猪HAL
基因型分布是否处于Hardy-Weinberg平衡状态。利用Popgene软件计算等位基因频率、基因型频率和遗传多态性指标。2 结果与分析
2.1 猪组织基因组DNA提取结果
猪基因组DNA电泳检测见图1,提取的基因组DNA为一条较致密、清晰的条带,并且拖尾不严重,表明基因组DNA的完整性较好,含量较高。
图1 组织基因组DNA
2.2 PCR扩增结果
猪HAL
基因的扩增PCR产物大小为659 bp,见图2,与预计的片段长度一致,无非特异条带,可用作下一步酶切试验。
图2 猪HAL基因扩增结果
2.3 HAL基因PCR-RFLP酶切结果
猪HAL基因扩增产物酶切电泳检测分型结果见图3。HAL
基因存在3种基因型(HAL
HAL
,HAL
HAL
,HAL
HAL
型),该突变位点cDNA第1 834位点C>T,引起Hha I限制性酶切位点的存在与丢失。HAL
HAL
型为493、166 bp二条带型,HAL
HAL
型为659、493、166 bp三条带型,HAL
HAL
型为659 bp单条带型。本研究的3个猪群中均没有检测到HAL
HAL
基因型。
图3 猪HAL基因HhaI酶切结果
2.4 HAL基因的基因型频率和等位基因频率

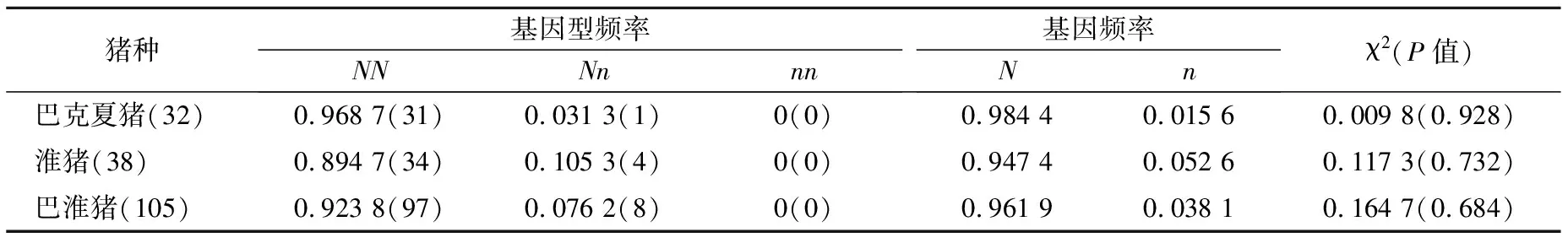
表1 猪HAL基因Hha I酶切位点的PCR-RFLP基因型频率和基因频率
2.5 HAL基因的遗传多态性分析
猪HAL
基因Hha I酶切位点的遗传多态性分析见表2。所检测猪种的HAL
基因,纯合度Ho:巴淮猪(0.926 7)介于巴克夏猪(0.969 2)和淮猪(0.900 3)之间;杂合度He:巴淮猪(0.073 3)介于淮猪(0.099 7)和巴克夏猪(0.030 8)之间。巴克夏猪、淮猪和巴淮猪的HAL
基因Hha I酶切位点均处于低度多态(PIC
<0.25),且巴淮猪多态信息含量 (polymorphism information content, PIC)(0.070 6)处于巴克夏猪(0.030 3)、淮猪(0.094 8)之间。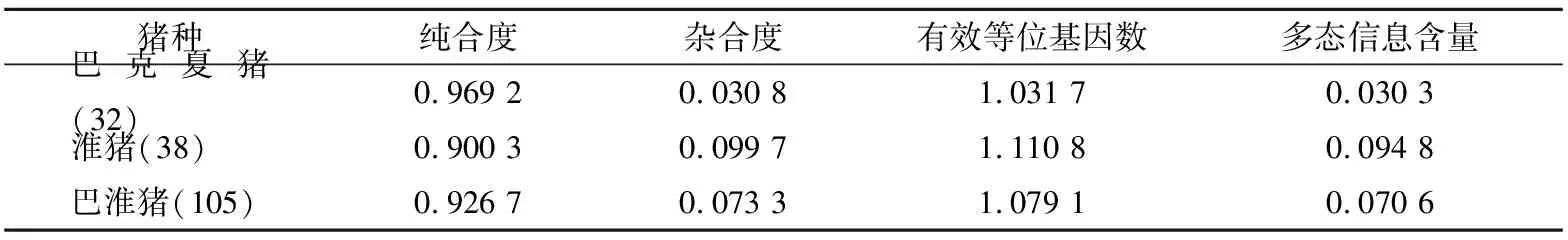
表2 猪HAL基因的遗传多态性指标
3 结论与讨论
HAL
基因是与猪PSE肉相关的主效基因,尽管HAL
等位基因可以增加猪的瘦肉水平,但隐性纯合基因型造成的猪应激综合征会带来严重经济损失,因此不能忽视猪群中等位基因HAL
的检测。研究表明,猪氟烷敏感基因(HAL
)具有多效性,一方面导致肌肉肥大并可增加胴体瘦肉率,另一方面易可诱导发生猪应激综合征。肖倩等研究表明,携带了HAL
隐性等位基因的个体在生长速度、饲料转化效率等方面表现出优异性,比不携带HAL
隐性等位基因的个体更好。石磊对我国引进猪种(大白猪、杜洛克猪和长白猪)的检测中,发现HAL
HAL
猪的背膘厚度比HAL
HAL
和HAL
HAL
型猪薄。本研究对32头巴克夏猪、38头淮猪和105头巴淮猪的HAL
基因进行检测,结果发现3个猪群均处于Hardy-Weinberg平衡状态,没有检测到HAL
HAL
基因型。其中巴克夏猪HAL
基因Hha I酶切位点HAL
HAL
基因型频率较高、HAL
HAL
基因型频率较低;等位基因HAL
频率(98.44%)高于HAL
频率(1.56%)。与皮特兰猪相比,巴克夏猪等位基因HAL
频率结果较低,与我国蒙山黑土猪种相比结果类似。淮猪HAL
基因Hha I酶切位点HAL
HAL
基因型频率较高,HAL
HAL
基因型频率较低;等位基因HAL
频率(94.74%)高于HAL
频率(5.26%)。与报道的黔北黑猪、宁乡猪相比,淮猪等位基因HAL
频率结果偏高。由于氟烷基因HAL
会对猪的肉质、生产性能造成一定的影响,应加快筛除HAL
等位基因,及时淘汰HAL
基因携带者以及可能出现在猪群体中的隐性个体,从而保证猪场的经济效益。本研究中,3个猪群HAL
基因Hha I酶切位点均检测到了HAL
HAL
基因型,但为了防止由HAL
HAL
基因型个体交配出现隐性纯合子而导致猪肉品质下降,在猪群体中剔除HAL
等位基因是必要的。所检测猪种的HAL
基因纯合度Ho
和杂合度He
,巴淮猪(0.
073 3)均介于淮猪(0.
099 7)和巴克夏猪(0.
030 8)之间。巴克夏猪、淮猪和巴淮猪的HAL
基因Hha I酶切位点均处于低度多态(PIC
<0.25),且多态信息含量(PIC)巴淮猪(0.070 6)处于巴克夏猪(0.030 3)、淮猪(0.094 8)之间,结果提示育种过程应加快淘汰HAL
基因的淮猪群体。综上所述,本试验在巴克夏猪、淮猪及它们的F代巴淮猪中均检测到杂合子HAL
HAL
,应及时淘汰HAL
基因携带者,为巴克夏猪、淮猪(霍寿黑猪)的开发利用以及加速育成肉质优良的巴淮猪新品系提供一定的参考依据。