Fas对镉激活PC12细胞线粒体凋亡通路的影响
2021-08-23闻双全顾建红刘学忠卞建春刘宗平
闻双全,陈 洁,王 莉,邹 辉,顾建红,刘学忠,卞建春,刘宗平,袁 燕*
(1.扬州大学兽医学院,扬州 225009; 2.江苏高校动物重要疫病与人兽共患病防控协同创新中心, 扬州 225009; 3.江苏省南通市动物疫病预防控制中心,南通 226011)
镉是环境中常见的重金属污染物,可经皮肤、消化道和呼吸道进入人、畜体内。进入机体内的镉排泄率极低,其生物半衰期长达10~30年。脑是镉毒性作用的重要靶器官之一,镉可增加血脑屏障通透性并进入大脑[1]。本实验室前期研究发现,镉可在大鼠大脑皮质内蓄积[2]。大量研究表明,镉可造成人和动物神经系统功能障碍[3-5]。镉致神经系统毒性机制主要包括诱导氧化应激、凋亡和自噬[6-7]。其中,细胞凋亡是镉致神经毒性的重要机制之一。
细胞凋亡是由基因调控的一种细胞程序性死亡,在调节细胞稳态和多细胞生物发育过程中起着重要作用。Fas是肿瘤坏死因子受体超家族的一员,通过与其配体FasL结合在多种细胞凋亡中发挥关键作用[8-10]。当细胞受到凋亡刺激时, FasL与Fas结合,并通过进一步与Fas相关死亡结构域蛋白(FADD)结合,激活半胱氨酸蛋白酶-8(caspase-8),引起细胞凋亡[11];另外,活化的caspase-8可剪切BH3相互作用域死亡激动剂(BID),诱导线粒体释放细胞色素C(Cyt C),介导线粒体凋亡通路[12]。
本实验室前期研究发现镉可通过激活Fas/FasL通路和线粒体通路诱导神经细胞凋亡[13-14]。但是,这两条通路之间的关系以及Fas在镉致神经细胞凋亡线粒体通路中的作用和调控机制尚未完全阐明。本试验以Fas基因沉默的大鼠肾上腺嗜铬细胞瘤细胞(PC12)为模型,用镉进行处理,通过Western blot、免疫荧光染色等方法探究Fas在镉致PC12细胞凋亡中的作用及其对线粒体通路的调控机制,为揭示镉的神经毒性机制提供理论依据。
1 材料与方法
1.1 细胞
PC12细胞购自中国科学院上海生命科学研究院细胞库。Fas基因沉默PC12细胞株(Fas shRNA组,靶序列:5′-GATCCGGTGCGTGTCAAGCT TTAATCTTCAAGAGAGATTAAAGCTTGACA CGCACCTTTTTTA-3′)和插入非特异性序列的对照细胞株(NC组)由实验室构建筛选获得。
1.2 主要试剂与仪器
RPMI 1640、马血清和胎牛血清购自Life Technologies公司;L-谷氨酰胺和青链霉素购自BBI公司;醋酸镉和L-多聚赖氨酸购自Sigma公司;细胞线粒体分离试剂盒、Cy 3标记山羊抗兔荧光二抗和DAPI染色液购自碧云天生物技术研究所;凋亡诱导因子(AIF)和核酸内切酶G(Endo G)兔多克隆抗体购自Abcam公司;Bcl-2相关X蛋白(Bax)、B细胞淋巴瘤/白血病-2基因(Bcl-2)、Cyt C、细胞色素C氧化酶亚基Ⅳ(COX-Ⅳ)、活化半胱氨酸蛋白酶-9(cleaved caspase-9)、活化半胱氨酸蛋白酶-3(cleaved caspase-3)、活化多聚二磷酸腺苷核糖聚合酶(cleaved PARP)、β-actin兔单克隆抗体和辣根过氧化物酶标记山羊抗兔二抗购自CST公司;BID兔多克隆抗体购自Novus公司;其余试剂均为国产分析纯。
二氧化碳培养箱购自美国Thermo公司;5810R型低温高速冷冻离心机购自德国Eppendorf公司;电泳仪、转膜仪购自美国BIORAD公司;激光共聚焦显微镜购自德国Leica公司。
1.3 细胞的培养与处理
Fas基因沉默的PC12细胞株和对照细胞株接种于RPMI 1640培养基(含10%马血清、5%胎牛血清、1%L-谷氨酰胺、1%青链霉素),置于37 ℃和5% CO2环境的培养箱中培养,当细胞生长状态良好且处于对数生长期时,用10 μmol·L-1Cd处理细胞12 h,分别命名为对照组(NC组)、镉组(Cd组)、Fas shRNA干扰组(Fas shRNA组)、Fas shRNA与镉共处理组(Fas shRNA+Cd组)。
1.4 细胞总蛋白、胞浆蛋白和线粒体蛋白的提取
收集各组细胞,加入适量含有蛋白酶抑制剂的细胞裂解液,冰上裂解30 min,超声裂解20 s;4 ℃,12 000 g·min-1离心10 min,收集上清即为总蛋白。
收集各组细胞,加入适量含有蛋白酶抑制剂的线粒体分离试剂重悬细胞团块,将细胞悬液转移至预冷的玻璃匀浆器中,冰上研磨,随后将匀浆转移至预冷的离心管,4 ℃,600 g·min-1离心10 min,吸取上清至新的离心管,4 ℃,11 000 g·min-1离心10 min,收集上清即为细胞浆蛋白,沉淀为线粒体;向沉淀中加入适量含有蛋白酶抑制剂的线粒体裂解液,冰上裂解30 min,12 000 g·min-1离心10 min,收集上清即为线粒体蛋白。
使用BCA法测定蛋白浓度并将蛋白浓度调整一致,按照1∶4加入5 × SDS-PAGE Loading Buffer,沸水煮10 min,蛋白保存于-80 ℃备用。
1.5 Western blot检测相关蛋白的表达
取等量样品进行SDS-PAGE电泳,转印至PVDF膜上,5%脱脂乳室温封闭1 h,4 ℃孵育相应一抗(BID、Bax、Bcl-2、Cyt C、cleaved caspase-9、cleaved caspase-3、cleaved PARP、AIF、Endo G、β-actin、 COX-Ⅳ抗体按1∶1 000稀释)过夜,室温孵育二抗(按1∶10 000稀释)1 h,ECL显影检测蛋白表达情况。使用Image J软件分析条带灰度值,目的蛋白与β-actin或COX-Ⅳ灰度值的比值即为目的蛋白的相对表达水平。
1.6 免疫荧光染色检测AIF核转位
用0.01% L-多聚赖氨酸包被爬片并将其置于24孔培养板中,将Fas shRNA组和NC组细胞接种于爬片,待其长至对数生长期时,用10 μmol·L-1Cd处理细胞12 h后,弃去培养基,磷酸盐缓冲液(PBS)洗涤2次;4%多聚甲醛固定细胞30 min,弃去固定液,PBS洗涤3次;加入0.5% 曲拉通X-100,室温透膜20 min,PBS洗涤3次;加入5% 牛血清白蛋白,室温封闭1.5 h,弃去封闭液;4 ℃孵育AIF抗体(1∶100)过夜,PBS洗涤3次;室温避光孵育Cy 3标记山羊抗兔荧光二抗(1∶200)1 h,PBS洗涤3次; 滴加DAPI染色液,室温避光孵育5 min,PBS洗涤2次;封片,激光共聚焦显微镜观察并拍照。
1.7 数据分析

2 结 果
2.1 Fas基因沉默对镉致PC12细胞BID活化的影响
10 μmol·L-1Cd分别处理NC组和Fas shRNA组细胞12 h,Western blot检测BID和tBID蛋白表达水平。结果如图1所示,与NC组相比,Cd组tBID/BID比值极显著升高(P<0.01);与Cd组相比,Fas shRNA+Cd组tBID/BID比值极显著降低(P<0.01)。
2.2 Fas基因沉默对镉致PC12细胞Bax/Bcl-2比值变化的影响
10 μmol·L-1Cd分别处理NC组和Fas shRNA组细胞12 h,Western blot检测Bax和Bcl-2蛋白表达水平。结果如图2所示,与NC组相比,Cd组Bax/Bcl-2比值极显著升高(P<0.01);与Cd组相比,Fas shRNA+Cd组Bax/Bcl-2比值极显著降低(P<0.01)。
2.3 Fas 基因沉默对镉致PC12细胞Cyt C释放的影响
10 μmol·L-1Cd分别处理NC组和Fas shRNA组细胞12 h,Western blot检测Cyt C在细胞内分布情况。结果如图3所示,与NC组相比,Cd组线粒体中Cyt C表达量极显著降低(P<0.01),细胞质中Cyt C表达量极显著升高(P<0.01);与Cd组相比,Fas shRNA+Cd组线粒体中Cyt C表达量极显著升高(P<0.01),细胞质中Cyt C表达量极显著降低(P<0.01)。
2.4 Fas 基因沉默对镉致PC12细胞caspase-9、caspase-3和PARP蛋白活化的影响
10 μmol·L-1Cd分别处理NC组和Fas shRNA组细胞12 h,Western blot检测caspase-9、caspase-3和PARP蛋白活化情况。结果如图4所示,与NC组相比,Cd组Cleaved caspase-9、Cleaved caspase-3和Cleaved PARP蛋白水平均极显著升高(P<0.01);与Cd组相比,Fas shRNA+Cd组Cleaved caspase-9蛋白水平显著降低(P<0.05),Cleaved caspase-3和Cleaved PARP蛋白水平极显著降低(P<0.01)。
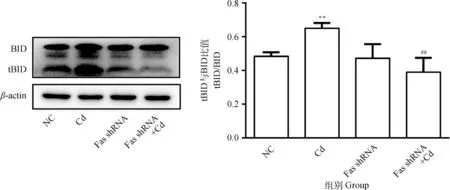
与NC组相比,**表示P<0.01;与Cd组相比,#表示P<0.05,##表示P<0.01。下同Compared with NC group, ** indicates P<0.01; Compared with Cd group, # indicates P<0.05, ## indicates P<0.01.The same as follows图1 Fas基因沉默对镉致PC12细胞BID活化的影响Fig.1 Effects of Fas silencing on the activation of BID in PC12 cells caused by cadmium

图2 Fas基因沉默对镉致PC12细胞Bax/Bcl-2比值变化的影响Fig.2 Effects of Fas silencing on the change of Bax/Bcl-2 ratio in PC12 cells caused by cadmium
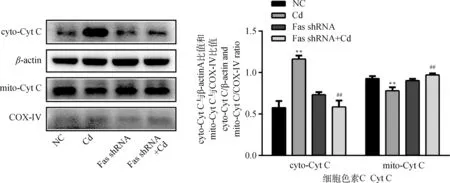
图3 Fas基因沉默对镉致PC12细胞Cyt C释放的影响Fig.3 Effects of Fas silencing on the release of Cyt C in PC12 cells caused by cadmium

图4 Fas基因沉默对镉致PC12细胞caspase-9、caspase-3和PARP蛋白活化的影响Fig.4 Effects of Fas silencing on the activation of caspase-9, caspase-3 and PARP in PC12 cells caused by cadmium
2.5 Fas 基因沉默对镉致PC12细胞AIF和Endo G蛋白表达的影响
10 μmol·L-1Cd分别处理NC组和Fas shRNA组细胞12 h,Western blot检测AIF和Endo G蛋白表达水平。结果如图5所示,与NC组相比,Cd组AIF和Endo G蛋白水平极显著升高(P<0.01);与Cd组相比,Fas shRNA+Cd组AIF和Endo G蛋白水平极显著降低(P<0.01)。
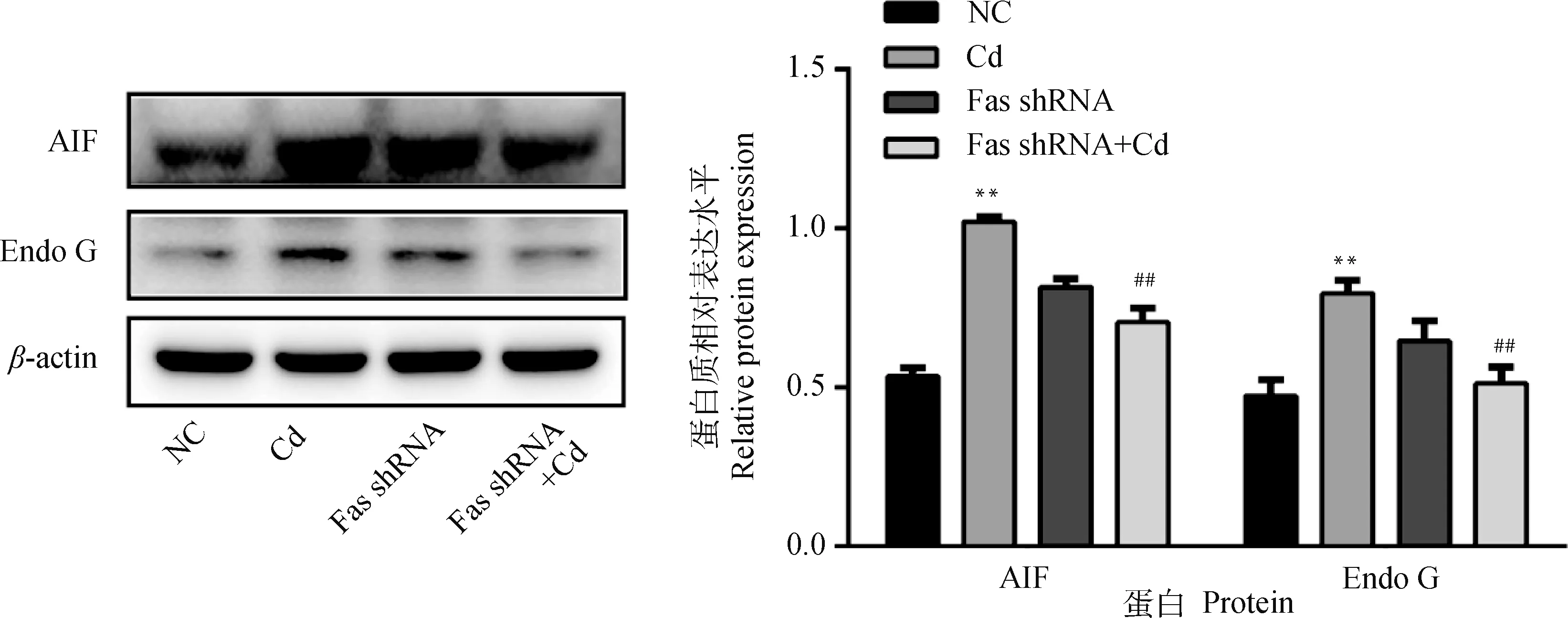
图5 Fas基因沉默对镉致PC12细胞AIF和Endo G蛋白表达的影响Fig.5 Effects of Fas silencing on the expression of AIF and Endo G in PC12 cells caused by cadmium
2.6 Fas 基因沉默对镉致PC12细胞AIF核转位的影响
10 μmol·L-1Cd分别处理NC组和Fas shRNA组细胞12 h,免疫荧光染色法检测AIF核转位情况。结果如图6所示,与NC组相比, Cd组荧光标记蛋白AIF主要集中于细胞核;与Cd组相比,Fas shRNA+Cd组荧光标记蛋白AIF主要分散于细胞质。
3 讨 论
细胞凋亡是由基因调控的程序性死亡,重金属镉可诱导包括神经细胞在内的多种细胞发生凋亡[6,15-16]。死亡受体通路是经典凋亡通路之一,其中,Fas/FasL信号通路备受关注。Fas/FasL信号通路主要通过Fas/FasL/FADD/caspase-8途径介导细胞发生凋亡。体内外研究表明,镉可激活Fas/FasL信号通路诱发神经细胞凋亡[13,17]。抑制Fas/FasL信号通路中的关键蛋白,能够对细胞凋亡起到一定抑制作用。Ullah等[18]研究发现,抑制Fas能够降低脑缺血导致的细胞凋亡。Zhang等[19]研究发现,干扰Fas显著抑制氧糖剥夺再灌注诱导的人神经母细胞瘤细胞凋亡。本实验室前期研究发现,利用Anti-FasL抑制FasL可减轻镉导致的PC12细胞凋亡[20]。

红色荧光代表AIF,蓝色荧光代表细胞核The red puncta represent AIF and the blue puncta represent nuclei图6 Fas基因沉默对镉致PC12细胞AIF核转位的影响(标尺=20 μm)Fig.6 Effects of Fas silencing on the nuclear transposition of AIF in PC12 cells caused by cadmium (scale bar=20 μm)
线粒体在哺乳动物细胞凋亡过程中发挥重要作用。线粒体功能障碍会导致活性氧积累,引起氧化应激和凋亡[21]。线粒体凋亡通路主要由Bcl-2家族蛋白介导。当各种凋亡刺激作用于线粒体时,会激活Bcl-2家族促凋亡成员,导致促凋亡和抗凋亡蛋白的失衡,从而影响线粒体膜通透性[22-23],导致线粒体释放Cyt C等各种促凋亡因子[24-25]。释放的Cyt C与凋亡蛋白酶活化因子-1(Apaf-1)结合形成复合物,激活caspase-9,进而激活caspase-3[26]。激活的caspase-3剪切PARP,抑制受损DNA的修复,促进凋亡细胞死亡[27-28]。体内外研究表明,镉可激活线粒体通路诱发神经细胞凋亡[17,29]。另外,线粒体通路和Fas/FasL通路之间存在紧密联系,Fas/FasL通路激活的caspase-8将BID剪切为tBID,tBID转位到线粒体,诱导Cyt C释放,从而激活线粒体通路[12]。Yu等[30]研究发现,在小鼠脊髓损伤模型中,Fas缺陷抑制BID、caspase-9和caspase-3的激活,增加抗凋亡蛋白Bcl-2和Bcl-xL的表达。Zhang等[19]研究发现,干扰Fas能够抑制氧糖剥夺再灌注导致的人神经母细胞瘤细胞Bax和Caspase-3上调及Bcl-2下调。本研究结果显示,Fas基因沉默能够显著或极显著抑制镉引起的PC12细胞tBID/BID比值和Bax/Bcl-2比值升高,Cyt C释放,caspase-9、caspase-3和 PARP蛋白活化。说明Fas通过调控caspase依赖的线粒体通路参与镉致PC12细胞凋亡。
AIF和Endo G是位于线粒体膜间隙的核酸酶,凋亡发生时被释放到细胞质中并转位到细胞核,通过caspase非依赖途径介导细胞凋亡[31-33]。大量研究表明,在细胞凋亡过程中,Fas可诱导AIF和Endo G从线粒体释放到细胞质或细胞核中[32,34-35]。Yu等[30]研究发现,在小鼠脊髓损伤模型中,Fas缺陷抑制AIF从线粒体转位到细胞核。本研究结果显示,Fas 基因沉默能够极显著抑制镉引起的AIF和Endo G蛋白表达升高,并抑制AIF核转位。说明Fas通过调控caspase非依赖的线粒体通路参与镉致PC12细胞凋亡。
4 结 论
镉通过激活线粒体通路诱导PC12细胞凋亡,干扰Fas能够抑制镉激活的线粒体凋亡通路,说明Fas通过调控线粒体通路参与镉致PC12细胞凋亡;这为揭示镉的神经毒性机制提供了新的理论基础。
