大黄酸对伪中间葡萄球菌的抗菌活性及作用机制
2021-08-23周志新付琳清王晗晟孙东波郑家三
宋 军,周志新,付琳清,王晗晟,刘 梦,孙东波,郑家三
(黑龙江八一农垦大学动物科技学院,大庆 163319)
伪中间葡萄球菌(Staphylococcuspseudintermedius)是一种凝固酶阳性条件致病菌[1],经常引起犬的脓皮病。这种常见的条件致病菌还能够引起犬、猫等动物的外耳道炎、伤口感染、尿道感染及其他一些感染[2]。耐甲氧西林伪中间葡萄球菌(methicillin-resistantStaphylococcuspseudintermedius,MRSP)及耐其他抗菌药物菌株的出现,已经超出兽医临床抗生素用药范围[3],引起了越来越多的关注。特别是近年来,在犬的主人和兽医身上也发现了伪中间葡萄球菌的定植,并相继出现伪中间葡萄球菌感染人的报道[4-5]。因此,伪中间葡萄球菌作为一种潜在的重要动物传染性病原菌,将会对人类公共健康造成严重威胁。
遏制动物源细菌耐药是人类控制细菌耐药的重要组成部分。针对目前细菌耐药性问题,包括抗菌肽、噬菌体、益生菌、中草药等抗菌剂或抗生素替代产品的研发逐步开展,其中,中草药来源广泛、安全性好,是绿色、有效的抗生素替代品,具有广泛的应用前景。大黄酸是一种活性蒽醌衍生物单体,主要从蓼科药用植物大黄中提取。大黄酸广泛存在于大黄、何首乌、虎杖等中药中,具有多种药理作用,如抗炎、抗肿瘤、抗氧化、抗菌、抗病毒和降血糖作用[6-11]。
前期国内外学者研究表明,大黄酸具有潜在的抗菌活性[12-14],与抗生素联用有增效或部分增效作用[15],并且能够抑制生物被膜形成,但对其作用机制研究较浅[16]。为此,本文以伪中间葡萄球菌为研究对象,探讨大黄酸的抗菌活性及抗菌机制,为犬脓皮病及伪中间葡萄球菌感染的防治提供基础。
1 材料与方法
1.1 菌株及药品来源
伪中间葡萄球菌分离株来源于2018—2020年大庆市农大动物医院脓皮症患犬,伪中间葡萄球菌ATCC 49444购自上海北诺生物科技有限公司;大黄酸(纯度>98%)购自中国食品药品检定研究院。
1.2 培养基与试剂盒
LB液体培养基、胰蛋白胨大豆肉汤(TSB)、Baird-Parker琼脂培养基、亚碲酸钠卵黄增菌液购自海博(青岛)生物有限公司;β-半乳糖苷酶活性检测试剂盒购自北京索莱宝科技有限公司;活性氧检测试剂盒购自碧云天生物技术有限公司。
1.3 主要仪器
电热恒温培养箱(上海森信实验仪器有限公司);超声波振荡仪(宁波新芝生物科技有限公司);低温冷离心机(德国艾本德股份公司);酶标仪Elx800(美国BioTek公司);Infinite 200 pro多功能酶标仪(瑞士TECAN);Hitachi S-4800 扫描电子显微镜(日本日立公司);JEM-2100 Plus透射电子显微镜(日本电子株式会社)。
1.4 细菌分离鉴定
1.4.1 细菌分离 无菌采集脓皮病患犬皮屑或脓汁共计38份,分别接种于LB液体培养基,37 ℃增菌培养8 h后,接种于Baird-Parker琼脂培养基中,培养24 h挑取平皿上黑色有透明带或带晕的单菌落接种于LB液体培养基,置于37 ℃,160 r·min-1纯培养12 h,进行革兰染色镜检。利用细菌DNA基因提取试剂盒提取分离株DNA,-20 ℃保存备用。
1.4.2 细菌分子生物学鉴定 参考Bannoehr等[17]伪中间葡萄球菌特异性引物16S rRNA、mecA(甲氧西林耐药基因)和hsp60(热休克蛋白)对分离菌株进行PCR扩增、鉴定。PCR扩增体系(总体积为25 μL):RNase-free 2×Taq PCR Mix12.5 μL,上、下游引物(表1)各1 μL,基因组DNA模板1 μL,H2O 9.5 μL。PCR反应程序参考Bannoehr等[17]的方法。PCR扩增产物经1%琼脂糖凝胶电泳鉴定后,送吉林库美生物科技有限公司进行测序、比对。
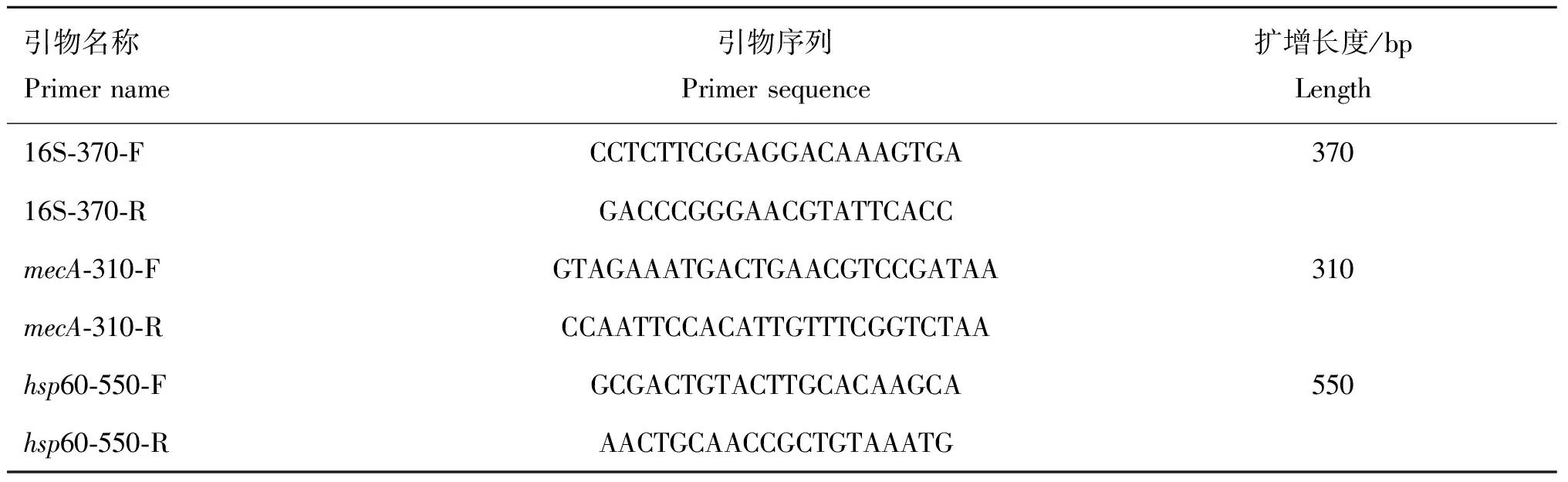
表1 16S rRNA, mecA和hsp60基因扩增引物[17]
1.5 大黄酸对伪中间葡萄球菌抗菌活性
1.5.1 最小抑菌浓度和最低杀菌浓度 最小抑菌浓度(minimum inhibitory concentration,MIC)的测定参考Song等[18]的方法略作修改。大黄酸以DMSO(二甲基亚砜)为溶剂,配制成5 mg·mL-1备用。用TSB培养基对大黄酸进行倍比稀释,每孔100 μL加至96孔微量培养板中,同时每孔加入100 μL 浓度约为105CFU·mL-1对数生长期菌液,37 ℃培养24 h,每组3个重复。TSB培养基不加药为阴性对照组。以没有肉眼可见的细菌生长的最低浓度为大黄酸的MIC。最低杀菌浓度(minimum bactericidal concentration,MBC)是指将MIC以上各孔培养物,分别吸取100 μL 接种在Baird-Parker琼脂基础培养基上,37 ℃ 恒温培养24 h,进行菌落计数,其菌落数少于5个的大黄酸浓度即为MBC[19]。
1.5.2 大黄酸对伪中间葡萄球菌生长的影响 将伪中间葡萄球菌培养至对数生长期,按1%接种不同浓度大黄酸(1/4×MIC、1/2×MIC、1×MIC与2×MIC)至无菌TSB培养基中,置于37 ℃,180 r·min-1摇床培养8 h,每间隔30 min取样测600 nm处吸光度,绘制时间-抑菌曲线。以未添加大黄酸的菌液作为对照组,每组3次重复。
1.6 溶血活性
参考Song等[18]的方法,采集健康犬血液在4 ℃ 下1 000×g离心5 min,得到犬红细胞(cRBCs)用阿氏液洗涤3次,用PBS (pH=7.4)稀释10倍。然后,将50 μL cRBCs与等量不同浓度的大黄酸(1×MIC、2×MIC、4×MIC和8×MIC)加入到96孔细胞培养板中,37 ℃孵育60 min 后测定上清液在450 nm处的吸光度。未经处理cRBCs的分别用0.1% Triton X-100和阿氏液处理,分别作为阳性和阴性对照。每组3个重复,测试两次。溶血活性按下式计算:溶血率(%)= [(样品OD570-阴性OD570)/(阳性OD570-阴性OD570)]×100。
1.7 黏附试验
参照李燕杰等[19]方法并进行适当修改,将对数生长期的ATCC 49444稀释到1×106CFU·mL-1。取含有不同浓度(1/4×MIC、1/2×MIC、1×MIC与2×MIC) 大黄酸的细菌悬液1 000 μL加入到24孔细胞培养板中(每孔含有无菌载玻片),37 ℃培养120 min。用PBS洗去未黏附的细菌细胞,将载玻片转入装有10 mL PBS溶液试管中,于20 kHz、25 ℃ 条件下超声处理60 s,平板计数评估细菌细胞的黏附率[19]。
1.8 大黄酸对伪中间葡萄球菌作用机制研究
1.8.1 细胞膜通透性测定 内膜通透性通过测定细菌在ONPG(邻硝基苯β-D-半乳吡喃糖苷)为底物的培养基中,细胞质β-半乳糖苷酶活性的释放。将ATCC 49444培养至对数生长期,并接种至含不同终浓度(1×MIC和2×MIC)的大黄酸液体培养基中,使含菌浓度为108CFU·mL-1,同时用PBS代替大黄酸药液作为对照组,37 ℃ 160 r·min-1摇床培养。每60 min各取1.0 mL,用0.22 μm无菌滤器过滤,取滤液,参照试剂盒说明书测定β-半乳糖苷酶活性。
1.8.2 活性氧测定 使用活性氧(ROS)检测试剂盒测定伪中间葡萄球菌内ROS的水平。将对数生长期的ATCC 49444稀释到OD600nm= 0.4和10 μmol·L-1DCFH-DA孵育20 min,PBS(pH 7.4)清洗和重悬菌体。100 μL细菌悬液与等体积不同浓度的大黄酸(1×MIC和2×MIC)混合后加入96孔荧光酶标板中,37 ℃条件下,每隔10 min,利用Infinite 200 pro多功能酶标仪(帝肯,中国)进行测定,其中激发波长为488 nm,发射波长为525 nm。 每个样品3个重复,测试两次。
1.8.3 扫描电子显微镜观察大黄酸对伪中间葡萄球菌细胞形态的影响 参照Wang等[20]方法处理扫描电镜样品,将培养至对数生长期浓度为108CFU·mL-1菌悬液与大黄酸(终浓度1×MIC)混合,37 ℃作用6 h后,4 000 r·min-1离心10 min,用PBS清洗3遍,菌体用2.5% (w/v)戊二醛4 ℃条件下固定过夜。随后,用乙醇(30%、50%、70%、85%、90%和100%)进行梯度脱水,用叔丁醇代替乙醇脱水2次,干燥样品,喷金制样,采用日立S-4800扫描电子显微镜观察。
1.8.4 透射电子显微镜观察大黄酸对伪中间葡萄球菌细胞超微结构的影响 前期菌体固定与扫描电镜样品制备方法一致,戊二醛固定后的菌体用1%锇酸溶液固定1 h,用PBS清洗菌体3遍,丙酮梯度脱水。环氧树脂包埋过夜,50 ℃烘干48 h。1%乙酸双氧铀染色,切片制样后采用JEM-2100 Plus透射电子显微镜观察。
1.9 数据处理
试验结果以“平均数±标准偏差”表示,采用GraphPad Prism 8统计学软件分析,组间的差异比较采用t检验,差异性检验水准设为P<0.05(*)差异显著,P<0.01(**)差异极显著。
2 结 果
2.1 革兰染色和生物学特性检测
分离菌株在Baird-Parker琼脂培养基中,培养24 h后,菌落呈黑色、湿润、隆起、光亮、表面光滑,外层有明显浑浊带(图1A);革兰染色镜检为阳性球菌(图1B),菌呈球形,大小均匀,葡萄串状,革兰染色呈蓝紫色。

A. Baird-Parker琼脂生长特征; B.革兰染色结果(100×)A. Growth characteristics of Baird Parker Agar; B. Results of Gram staining(100×)图1 分离菌株生物特征的检测结果Fig.1 Detection results of biological characteristics of isolated bacteria
2.2 分子生物学鉴定
分离菌株16S rRNA,mecA和hsp60基因均扩增出特异性条带,与图2所示基因片段大小相一致。利用Blast程序对测序结果进行分析比对,共分离到20株伪中间葡萄球菌,分离率为52.63%(各菌株同源性均达98%以上)。
2.3 大黄酸对伪中间葡萄球菌的抗菌活性
通过测定大黄酸对伪中间葡萄球菌标准株和分离株(20株)的抗菌活性,表明大黄酸对伪中间葡萄球菌最小抑菌浓度(MIC)为12.5 μg·mL-1,最小杀菌浓度(MBC)为25 μg·mL-1或50 μg·mL-1。
2.4 抑菌动力学曲线
向对数生长期菌液中加入不同浓度大黄酸后,伪中间葡萄球菌呈不同程度的抑制(图3)。加入大黄酸的终浓度为2×MIC时,8 h内伪中间葡萄球菌被完全抑制,菌液澄清;当大黄酸在亚抑菌浓度(1/2×MIC)时,细菌生长缓慢,3 h内能够抑制细菌生长,但随着作用时间延长,细菌逐渐增长。上述结果说明,大黄酸能明显抑制伪中间葡萄球菌的生长。

A. 16S rRNA扩增结果; B. mecA扩增结果; C. hsp60扩增结果。 M.DNA分子量标准(100~2 000 bp);1.阴性对照;2.目的条带A. Results of 16S rRNA amplification; B. Results of mecA amplification; C. Results of hsp60 amplification. M. DNA molecular weight standard (100-2 000 bp); 1. Negative control; 2. Target band图2 标准株16S rRNA, mecA和hsp60基因PCR扩增结果Fig.2 PCR amplification results of 16S rRNA, mecA and hsp60 of standard strain
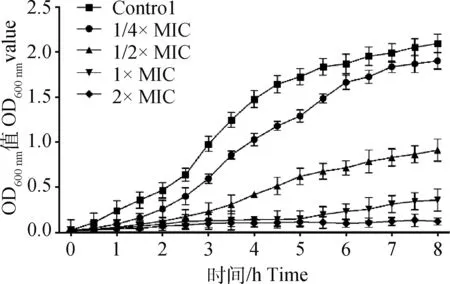
图3 不同浓度大黄酸对伪中间葡萄球菌的抑菌活性曲线Fig.3 Antibacterial activity curve of different concentrations rhein against Staphylococcus pseudintermedius
2.5 溶血活性
为了进一步了解大黄酸对哺乳动物细胞膜的毒性,检测了不同浓度大黄酸对犬血红细胞的溶血活性。当大黄酸浓度为1×MIC、2×MIC、4×MIC和8×MIC时,溶血活性分别为1.21%、1.86%、3.14%和5.14%。
2.6 黏附试验
为观察大黄酸对伪中间葡萄球菌早期生物膜形成的影响,评价生物膜形成的初始阶段——黏附。结果如图4,1/4×MIC的大黄酸能够抑制伪中间葡萄球菌的黏附,和对照组相比,抑制率为18.91%。随着大黄酸浓度的增加,大黄酸浓度为2× MIC时,抑制率为87.13%。说明大黄酸对黏附的抑制作用具有剂量依赖性。

图4 大黄酸处理可减少伪中间葡萄球菌的黏附Fig.4 Rhein treatment reduces adhesion of Staphylococcus pseudintermedius
2.7 细胞膜通透性测定
在细胞遭受损伤通透性改变的情况下,β-半乳糖苷酶会泄漏至胞外,通过检测胞外β-半乳糖苷酶活力的变化,也可反映细菌细胞膜的损伤情况[21]。在培养期间,对照组胞外β-半乳糖苷酶活性变化不显著,大黄酸(1×MIC和2×MIC)处理组胞外β-半乳糖苷酶活性随着时间延长而增加(图5)。
2.8 大黄酸对伪中间葡萄球菌ROS的影响
如图6所示,大黄酸对伪中间葡萄球菌产生的ROS具有浓度依赖性作用。在浓度为1×MIC时产生的ROS是空白对照组的3.9倍。在浓度为2×MIC时,荧光强度比空白对照组增加了6.4倍。总之,和对照组(大黄酸未处理组)相比,大黄酸处理后使伪中间葡萄球菌细胞产生大量ROS。
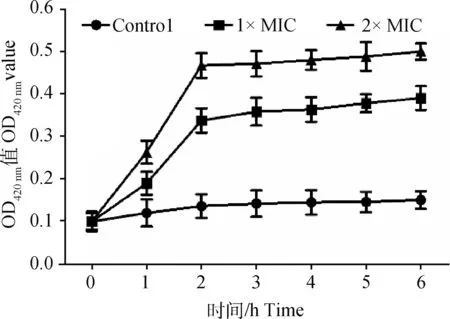
图5 大黄酸作用前后伪中间葡萄球菌悬液中β-半乳糖苷酶活力变化Fig.5 Effect of rhein on β-galactosidase activity of Staphylococcus pseudintermedius
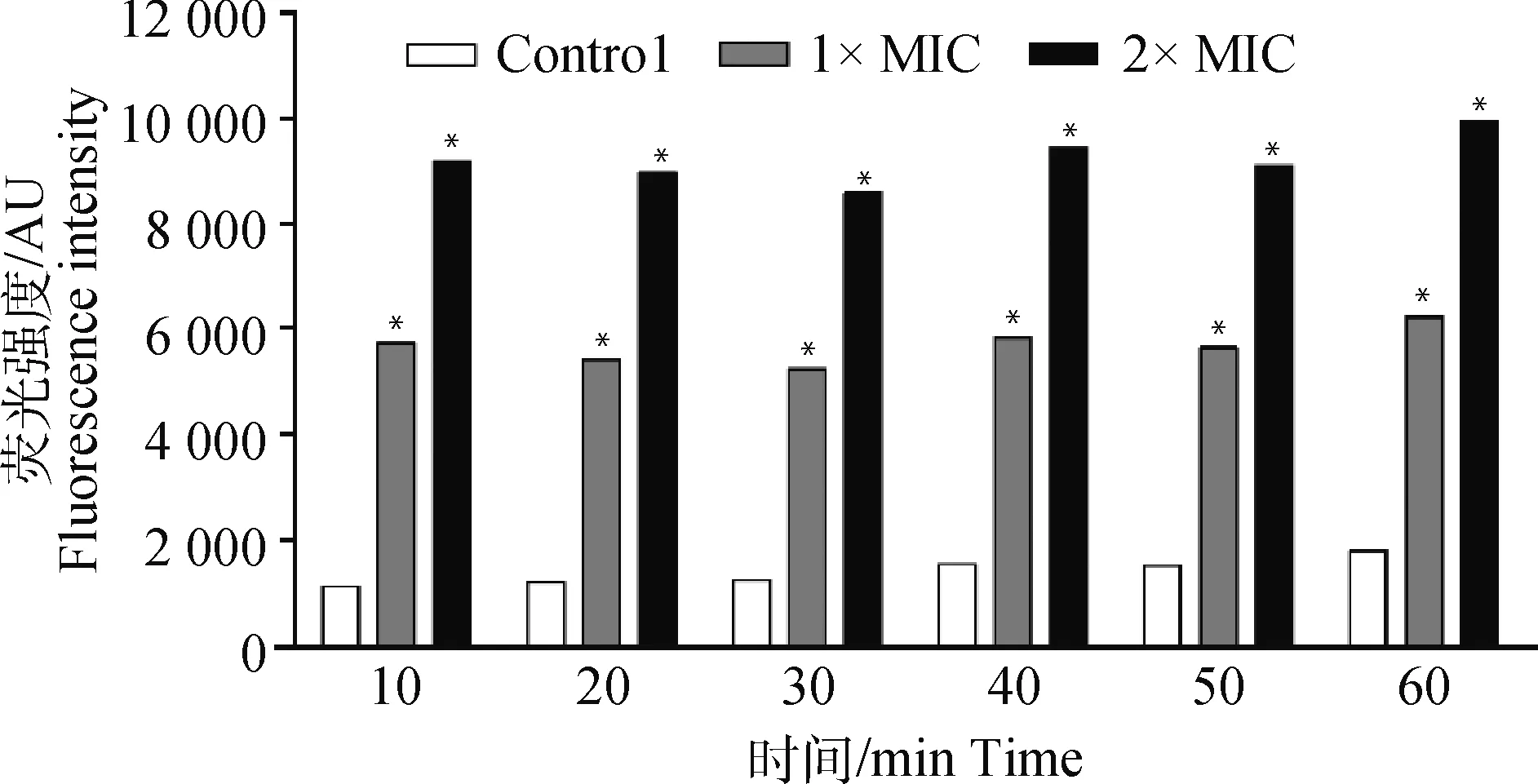
图6 大黄酸对伪中间葡萄球菌ROS的影响Fig.6 Effect of rhein on ROS of Staphylococcus pseudintermedius
2.9 大黄酸对伪中间葡萄球菌细胞形态和超微结构的影响
通过扫描电子显微镜和透射电子显微镜直接的观察伪中间葡萄球菌的外部形态和内部结构的变化[18,21]。SEM观察结果显示,未加大黄酸空白对照组(图7A)伪中间葡萄球菌形态饱满圆润、表面光滑。大黄酸(1×MIC)处理组(图7B)大部分细胞呈黏连状态,细胞表面存在大量分泌物、细胞皱缩。TEM观察结果(图7C)显示,未经大黄酸处理的伪中间葡萄球菌细菌细胞壁和细胞膜完整,内容物致密,分布均匀。大黄酸(1×MIC)处理的细胞,电子密度降低、细胞壁破裂、内容物泄漏(图7D)。说明大黄酸对伪中间葡萄球菌的内部结构及外部形态均造成了破坏性影响。
3 讨 论
宠物源耐药细菌的出现被认为是潜在的公共卫生问题,严重威胁着宠物主人和兽医的健康[22]。伪中间葡萄球菌就是其中之一。伪中间葡萄球菌是一种凝固酶阳性条件致病菌,具有多重耐药性,是引起犬脓皮病最主要的致病菌[23]。因此,迫切需要天然、安全、效果好的抗生素替代物,控制宠物源耐药细菌的感染。
本研究从38份临床样本中分离得到20株耐甲氧西林伪中间葡萄球菌,说明伪中间葡萄球菌是犬脓皮症最主要的致病菌之一。测得大黄酸对耐甲氧西林伪中间葡萄球菌最小抑菌浓度(MIC)为12.5 μg·mL-1。不同抗菌药物、抗菌肽、天然产物等抑制细菌生长的机制较为复杂,包括针对细菌细胞壁、细胞膜的合成,核酸和蛋白质的合成和抑制,干扰细菌代谢等[24]。本研究中,亚抑菌浓度的大黄酸能够在3 h内抑制伪中间葡萄球菌的生长,之后随培养时间的延长缓慢增长(图3)。说明大黄酸可能弱化了细胞壁对维持细胞形态的作用[25],导致细菌被抑制。为评估药物的安全性,本研究测定了大黄酸对犬红细胞的毒性,当大黄酸浓度高达8×MIC 时,对犬红细胞几乎没有溶血活性,说明大黄酸具有治疗犬脓皮病的潜力。黏附是细菌定植入侵及形成生物被膜的关键步骤[26]。本研究通过测定不同浓度的大黄酸对伪中间葡萄球菌黏附的影响,评价其抗生物膜作用,结果表明,在体外亚抑菌浓度下,大黄酸可以阻止伪中间葡萄球菌的黏附,减少细菌定植和影响生物被膜的形成。

A、C 空白对照组;B、D. 1×MIC大黄酸处理组A,C Blank control group; B,D. 1×MIC rhein treated group图7 电镜观察大黄酸对伪中间葡萄球菌细胞形态和超微结构的影响Fig.7 The effect of rhein on the cell morphology and ultrastructure of Staphylococus pseudintermedius by electron microscope
细菌的保护屏障细胞膜,在防御系统中发挥着重要作用[27]。同时,细胞膜也是许多药物、抗菌肽等作用的靶点之一[28]。前期,关于大黄酸抗菌机制的研究较少,国内学者证明了大黄酸通过损伤细菌的细胞膜,从而抑制细菌的生长和繁殖[16]。本研究通过测定大黄酸作用后伪中间葡萄球菌细胞膜通透性和完整性,评价大黄酸的抑菌作用机制。结果显示,大黄酸处理后,胞外β-半乳糖苷酶活性随着时间延长而增加,表明大黄酸能改变伪中间葡萄球菌细胞膜通透性,而且能破坏细胞膜,使存在于细胞浆内的核酸和酶等生物大分子外流,细胞营养物质丢失,从而发挥抑菌抗菌作用,与覃静等[16]报道的大黄酸对金黄色葡萄球菌的作用结果相似。此外,有研究表明,抗菌药物能够通过刺激细菌细胞产生高水ROS,发挥抗菌作用[29-30]。因此,本研究探讨了不同浓度的大黄酸对伪中间葡萄球菌的ROS水平。当大黄酸终浓度为1×MIC和2×MIC时,细菌细胞内ROS水平显著高于对照组。过量ROS的产生会导致细菌氧化损伤,影响细菌活性,甚至导致细菌死亡[31]。最后,本研究结合扫描电镜和透射电镜观察,发现大黄酸处理后,细菌细胞表面存在大量分泌物呈黏连状态,细胞表面褶皱、内部电子密度降低、细胞壁破裂、内容物外流,说明大黄酸对伪中间葡萄球菌的外部形态及内部结构均造成破坏性影响,与上述结果相吻合。
综上所述,大黄酸通过多种途径对伪中间葡萄球菌发挥抑菌和杀菌作用,为犬脓皮病及伪中间葡萄球菌感染的防治提供基础。
4 结 论
大黄酸通过弱化伪中间葡萄球菌细胞壁对细胞形态的维持,破坏细胞膜的通透性、完整性,并诱导细菌细胞产生大量ROS,从而发挥抑菌和杀菌作用。这为研究中药单体作为抗菌药物治疗犬脓皮病提供了理论基础和科学依据。
