外来入侵植物肿柄菊在云南的扩散风险研究
2021-08-16陈剑王四海朱枫袁春明
陈剑 王四海 朱枫 袁春明

摘 要: 預测外来入侵植物的潜在入侵范围以及对不同环境的适应性,能够为外来植物入侵提供预警和预防。该文通过实地调查肿柄菊(Tithonia diversifolia)在云南的入侵范围,以及收集世界其他地区分布点,结合12个关键环境变量,利用MaxEnt模型对肿柄菊适生区范围和环境限制因子进行分析。结果表明:在世界范围适生区分布格局中,肿柄菊在云南无高适生区,次适生区总面积占全省总面积的3.58%,中适生区占32.90%,低适生区占42.80%,非适生区占20.72%;有扩散风险的区域主要为中适生区,普遍分布于云南省26° N以南地区;实地调查肿柄菊分布最高海拔2 200 m,在海拔1 000~1 500 m分布最为密集,而模型预测显示海拔2 000 m以下都适宜肿柄菊生长,且500~2 000 m为最适宜海拔范围;在12个环境变量中年降雨量、人类足迹、年均温、海拔等环境变量对肿柄菊的分布起到了最重要的作用。综上结果表明,目前在云南肿柄菊零星的分布点已到达其潜在分布区的边缘,但已入侵区域还远小于潜在可入侵区域。为了预防肿柄菊在我国南方地区大面积扩散带来危害,有必要对其入侵和扩散态势进行监测。
关键词: 外来入侵植物, 肿柄菊, MaxEnt模型, 环境变量, 扩散
中图分类号: Q948
文献标识码: A
文章编号: 1000-3142(2021)05-0789-10
Risk evaluation of Tithonia diversifolia dispersal in Yunnan Province,China
CHEN Jian1,2, WANG Sihai1,2*, ZHU Feng3, YUAN Chunming 1,2
( 1. Yunnan Academy of Forestry and Grassland, Kunming 650201, China; 2. Yunnan Key Laboratory of Forest Plant Cultivation and Utilization /State Forestry Administration Key Laboratory of Yunnan Rare and Endangered Species Conservation and Propagation, Kunming 650201, China; 3. College of Geography and Ecotourism, Southwest Forestry University, Kunming 650224, China )
Abstract: Predicting the potential range of invasive alien plants and their adaptabilities to different environments could give early warning and prevention measures. The diverse bioclimatic types in Yunnan province would provide rich potential habitats for alien invasive plants. Tithonia diversifolia, an alien plant invading in tropical and sub-tropical regions of Yunnan, has quickly dispersed in recent years. In order to evaluate the risk of its invasion in different geographical environments, we investigated the invasion situation of T. diversifolia in Yunnan Province, and collected the distribution samples in the world. Using this species distribution coordinates and 12 main environmental variables, we analyzed the suitability of T. diversifolia to different environments and the dominating environmental factors by MaxEnt model. The results were as follows: Yunnan had not high-suitability area in the worldwide pattern of T. diversifolias distribution, meanwhile sub-, medium-, low-, un-suitability area accounted for 3.58%, 32.90%, 42.80% and 20.72% of the provincial area, respectively; Medium-suitability area was the main risk region, which widely interspersed in the south of 26° N in Yunnan; Field investigation showed T. diversifolia mainly distributed at the altitude of 1 000-1 500 m and its upper altitudinal limit was 2 200 m, while the model predicted that the altitude region below 2 000 m was suitable, especially between 500-2 000 m; The most important environmental factors affecting distribution of T. diversifolia were annual precipitation, human footprint, annual average temperature and altitude. In conclusion, although T. diversifolia has reached sporadically its suitable geographical limiting boundary at present, the invaded area is far less than the potential suitable area. It is necessary to keep monitoring T. diversifolias dispersal dynamic in the future, so that hazard caused by T. diversifolia could be controlled in time.
Key words: alien invasive plant, Tithonia diversifolia, MaxEnt model, environmental variables, dispersal
肿柄菊(Tithonia diversifolia)为菊科肿柄菊属一年生草本植物,原产墨西哥及中美洲地区,曾作为观赏植物、绿肥和防治土壤侵蚀植物被广泛引种到亚洲、非洲和澳洲的许多国家和地区。目前在东南亚、非洲、太平洋的一些热带和亚热带地区成为入侵杂草(Yang et al., 2012;Jama et al., 2000)。肿柄菊在我国的云南、广西、广东、福建、海南、台湾等地已普遍分布(王四海等,2004; 罗瑛等,2009)。在云南,近年来随着高速公路网的建设,肿柄菊的擴散速度明显加快,有向更高纬度和海拔扩散的趋势(朱枫等,2018)。肿柄菊在热带和亚热带地区呈灌木或亚灌木状生长,种子千粒重较大(4.5~6.5 g),种子的远距离传播要依附于交通工具、人畜或其他媒介,这种具有特定路线和方式的传播特点增加了其侵占新领地的时滞(王四海等,2008),从而可能导致肿柄菊的当前实际分布和潜在适生区具有较大的空间差异。另外,云南由于受复杂地形和大气环流的影响,各种气候类型呈现出相互镶嵌的特点,影响植物分布的两个主要因素(温度和降雨)组合类型多样(陈宗瑜,2001),在这种环境条件下肿柄菊的适应性会随着环境改变而不断发生着变化(王四海等,2019)。当前云南从北热带到北亚热带气候类型均有肿柄菊分布,但是在不同的气候区域群落面积分布极不均匀,在北亚热带仅为点状的零星分布(朱枫等,2018)。扩散时滞和环境适应性都可能引起肿柄菊群落地理空间分布的不均匀,对肿柄菊在云南多变环境下的适应性研究能确定其入侵潜能。
肿柄菊在引种扩散过程中已适应多种生态气候,在不同的地区限制其分布的环境因子会有差异。肿柄菊在100年前引入非洲的乌干达,现在已扩散到非洲的20多个国家,主要分布在撒哈拉以南降雨较多的区域,向北的继续扩散受到干旱环境的限制(Obiakara & Fourcade, 2018);在原产地墨西哥,肿柄菊在北部的分布会受到冬季霜冻影响(Lentz et al., 2008)。云南多样化的地理环境可能会导致肿柄菊的适应性在空间分布更为复杂。利用生态位模型或生境适应性模型探讨引种植物对新环境的适应能力是近些年常使用的方法,通过模拟物种在环境中的生态位,然后把物种分布区投射到地理空间上,进而实现预测物种的潜在分布范围(Elith et al., 2010)。在众多植物分布预测模型中,MaxEnt模型运用最为普遍,常用来预测外来入侵植物潜在分布区(许仲林等,2015;刘晓彤等,2019)。本研究通过MaxEnt模型,结合气候、土壤、海拔、人类影响指数(人类足迹)等多种环境因子对肿柄菊在云南的适应性进行分析,拟确定肿柄菊在不同区域的潜在适应能力和将来的入侵范围,以及影响肿柄菊分布的主要环境因子。
1 材料与方法
1.1 肿柄菊分布信息采集和分析
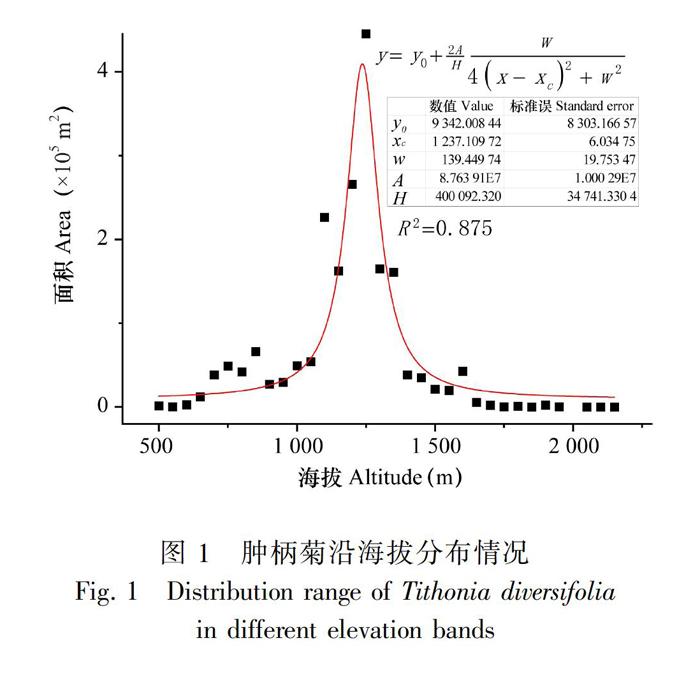
根据肿柄菊易于沿公路扩散和分布的特点(王四海等,2004),在2016年末和2017年初肿柄菊开花结实期,群落外貌特征具有较远距离能明显识别的季节,沿云南主要国道线和省道线沿路调查肿柄菊分布情况。依据群落为斑块分布的特点(王四海等,2004),记录斑块(分布点)的经纬度、海拔信息,并采用目测法快速评估斑块面积,调查范围覆盖肿柄菊在云南的分布区域。根据调查数据,从分布最低点起把每50 m高差内的肿柄菊群落面积进行累加,所获得的各海拔段群落面积用Oringin 9.1软件拟合其分布曲线,获得肿柄菊沿海拔分布规律。
1.2 肿柄菊适生区分析
(1)软件及使用
肿柄菊适生区分析利用MaxEnt V3.4.1和arcgis 10.2。MaxEnt用于物种适生区建模运算,输入数据为物种地理分布数据、海拔、气候变量、土壤变量和人类足迹。模型重复运行类别为交叉验证,样本分布数据共分为10组,每一组数据均用于建模和交叉验证(Urbani et al., 2017),重复次数为5,运算后输出ASCII格式的物种适生区预测结果,用arcgis 10.2将其转换成栅格图层并按生境适宜度(0~1.00)进行分级,获得肿柄菊分布的5个适宜性等级,即高适生区(0.80~1.00)、次适生区(0.60~0.80)、中适生区(0.40~0.60)、低适生区(0.20~0.40)、非适生区(0~0.20),并按该分级标准对其进行重分类后计算各级适生区面积。模型的预测准确性以模型计算的ROC曲线的AUC值(曲线与横坐标所围成的面积)进行评价,AUC 评价指标(Swets, 1988):很差(≤0.060);较差(0.600~0.700);一般(0.700~0.800);良好(0.800~0.900);极好(0.900~1.000)。
(2)环境数据来源与处理
肿柄菊参与建模数据来自两部分,第一部分为从全球生物多样性信息网站(https://www.gbif.org/)、中国国家标本资源平台(www.nsii.org.cn)、中国数字植物标本馆(http://www.cvh.ac.cn/)等平台上查询世界范围内肿柄菊分布点,并除去重复、错误的分布点;第二部分来自本研究实地采集的群落样点。为防止模型过度拟合,以上两部分分布点数据每5 km范围内只保留一个分布点(Veloz, 2009),最终共采用1 850个世界分布点(含云南省范围内的3个分布点),460个云南省实地采集分布点,统一转换为十进制小数格式的.CSV文件用于建模。
世界和中国云南行政边界矢量图来自国家地理信息系统网站(http://ngcc.sbsm.gov.cn/);19个典型气候变量和海拔数据来自世界气候数据库(www.worldclim.org),气候数据为1960—1990年共30 a的平均值,空间分辨率为30 arc sec。因19个气候变量为非独立变量,为提高模型模拟准确度,用arcgis 10.0 多元分析工具各变量进行相关性分析,对Pearson相关系数大于0.8的某一组数据集仅保留其中一个(Xu et al., 2019),最终使用的气候变量为BIO1、BIO2、BIO10、BIO12、BIO15、BIO17、BIO18、BIO19等8个(表1);人类足迹数据来自哥伦比亚大学社会经济数据及应用中心(https://sedac.ciesin.columbia.edu),空间分辨率为30 arc sec;土壤变量来自世界土壤数据库(http://www.fao.org/soils-portal/soil-survey/soil-maps-and-databases/harmonized-world-soil-database-v12/en/),空间分辨率为30 arc sec,共34个土壤变量,参与MaxEnt建模后,仅列出参与建模重要值大于1的两个变量S_GRAVEL和S_OC(表1)。以上所有环境数据均以云南省边界进行裁剪,并统一转换为ASCII格式。最终参与建模的环境变量共12个,各变量具体含义如表1。
为了分析肿柄菊随海拔升高的适应性变化,把全省面积根据海拔段<500 m、500~1 000 m、1 000~2 000 m、2 000~2 500 m、2 500~3 000 m、>3 000 m进行分类,根据MaxEnt适生区分析结果,计算在不同海拔段每类适生区面积占各自海拔段总面积的百分比。
2 结果与分析
2.1 肿柄菊在云南沿海拔分布现状
野外调查结果显示, 当前肿柄菊在云南分布最低海拔约在100 m,最高海拔为2 200 m,在1 000~1 500 m海拔段分布最为普遍(图1)。共调查到的肿柄菊群落面积累计为1 965 548 m2,1 000~1 500 m范圍内群落面积共计1 574 556 m2,占调查群落总面积的80.11%。可见在1 000 m以下和1 500 m以上海拔段肿柄菊有着更多的潜在分布区。
2.2 肿柄菊在云南适生区预测
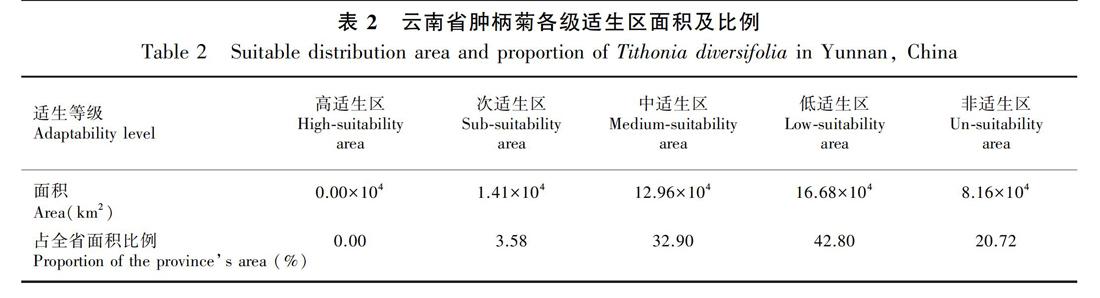
2.2.1 肿柄菊适生区分布格局 根据肿柄菊在全球适生状况预测,在云南没有高适生区,次适生区主要零星分布在云南中部,至西南部和南部的区域,以及西北部的河谷地带,面积约占全省总面积的3.58%;中适生区除滇西北和滇东北较高海拔区域外,大面积连续分布在云南的其余区域,占全省总面积的32.90%;低适生区间断分布于全省各地,在云南中部和南部地区主要位于海拔相对较高区域,在滇西北和滇东北主要位于海拔相对较低区域,低适生区占全省总面积的42.80%;非适生区主要分布在滇西北和滇东北,占全省总面积的20.72%(表2,图2)。肿柄菊在云南适生区域总体上随海拔高差而变化,不同的适生区呈镶嵌分布的特点。
在500 m以下海拔段,低适生区占该海拔段的面积最大,为67.59%;其次为中适生区,占29.43%;非适生区占2.94%。在500~1 000 m海拔段,次适生区、中适生区、低适生区和非适生区的面积分别占该海拔段总面积的4.28%、53.51%、38.09%和4.11%。在1 000~1 500 m海拔段,次适生区、中适生区、低适生区和非适生区的面积分别占该海拔段总面积的11.34%、49.71%、34.38%和4.56%(图3)。在1 500~2 000 m海拔段,中适生区和低适生区在该海拔段所占面积变大,分别为49.71%和44.20%;次适生区和非适生区占比较小,分别为1.98%和6.04%。到了2 000~2 500 m海拔段,低适生区面积占比显著变大,为73.50%。2 500~3000 m海拔段仅非适生区面积就高达74.83%,3 000 m以上海拔段非适生区面积已达100%(图3)。在500~2 000 m海拔段次适生区和中适生区所占面积都在50%以上,该海拔段最适宜肿柄菊生长;2 000 m海拔段以上,从低适生区面积最大过渡到非适生区面积最大,大于2 500 m海拔高度肿柄菊就难以生长。500 m以下海拔段低适生区面积占比较大,这可能是因为在云南500 m以下地区主要是河谷深切区域,特殊地形限制了肿柄菊的生长。
2.2.2 影响肿柄菊分布的主要环境因素 用于肿柄菊分布预测的12个环境变量中(表1),在肿柄菊适生区建模过程中,贡献率大于1%的有6个环境因子,这6个环境因子的贡献值占12个环境变量贡献值总和的98.5%,贡献值较多的环境因素为年降雨量、人类足迹、年均温、海拔、最暖季度平均温度、降雨季节性(表3)。从直接贡献值看主要影响肿柄菊分布的环境因素主要是年降雨量、人类足迹、年均温和海拔。置换重要值是将该特征变量值随机置换为数模型运算后获得的值,可在建模贡献率的基础上对各变量贡献进行再次评估,该值越大,表明该变量影响越大。影响肿柄菊分布的置换重要值较高的为年均温、人类足迹、最暖季度平均温度、年降水量(表3)。
综合以上情况,年均温、人类足迹、降水季节性、海拔、最暖季降水、最干季降水等6个环境变量对肿柄菊的分布起到了最重要的作用。图4为这6个环境变量在单独起作用时与肿柄菊出现概率的响应曲线,当各变量的值满足年降水量>852 mm、人类足迹影响指数>10、年均温17.2~23.7 ℃、海拔880~2 257 m、最暖季平均温18.7~25.2 ℃、降雨季节性33.7~122.3时,对肿柄菊的分布有最佳响应(肿柄菊出现概率为0.5以上),也即为肿柄菊最适环境变量范围。
2.2.3 模型精度评价 图5是检测模型精度的ROC曲线(受试者工作特征曲线),AUC值为曲线与横坐标所围的面积。AUC值越接近0.5(随机模拟值),说明模型精度越低; AUC值越接近1, 则模型精度越高。本研究中重复测试的AUC平均值为0.883,平均标准差为0.002。说明建模预测效果良好。
3 讨论与结论
3.1 肿柄菊的现有分布与潜在分布区
选择的12个环境变量中,年降雨量、人类足迹、年均温、海拔、最暖季度平均温度、降雨季节性等6个环境变量对肿柄菊的分布起到了最重要的作用。这种环境因素对肿柄菊分布的影响是基于对全球范围的总体分析,在不同的区域影响肿柄菊分布的主导因素可能会有所不同。在云南北热带、南亚热带和中亚热带这些适合肿柄菊生长的气候类型区,传播媒介(如公路网)是影响肿柄菊分布的主导因素(朱枫等,2018);然而在北亚热带地区,冬季的低温显著影响肿柄菊的结籽量,这一区域低的结籽量影响肿柄菊的进一步扩散(王四海等,2019)。在小尺度范围的生境类型也影响着肿柄菊的分布(朱枫等,2018)。在未来肿柄菊扩散过程中,由于局部影响分布主导因素的差异,即使适应性相同的区域也可能存在扩散蔓延速度不同。
通过当前肿柄菊在全球的分布点预测其潜在分布范围,除了原产地中美洲和南美洲外,肿柄菊还可以大面积分布于非洲、亚洲,总体上高适生区分布范围较少,预测分布区与已入侵区相对比,肿柄菊主要分布在次适生区和中适生区。云南当前分布格局与世界的基本类似,已带来严重生态危害的分布区域(王四海等,2004;朱枫等,2018;陈剑等,2020)也仅限于分布在次适生区和中适生区。由于肿柄菊在入侵的过程中气候生态位较为保守(Obiakara & Fourcade, 2018), 在未来肿柄菊造成严重危害的区域应主要是次适生区和中适生区。
尽管当前肿柄菊在云南分布的范围很广(跨越多于5个纬度,海拔变幅超过2 000 m),但只是在有些地区群落才高密度分布(朱枫等,2018),模型预测在500~2 000 m海拔段都较适宜肿柄菊生存,而当前1 000~1 500 m海拔段肿柄菊分布最为密集。这表明现有的分布远没有填满潜在的分布区域,未来扩散会在两个层面进行,一方面已经有肿柄菊分布的地方,将会继续扩散和蔓延使群落分布更为密集;另一方面会向环境适宜区尚未有分布的区域扩散,如沿怒江、澜沧江、金沙江更为上游的河谷区域,以及云南东部的一些区域。
3.2 肿柄菊在我国的分布潜力
云南海拔高度从76.4 m到6 740 m,垂直高差达6 000 m以上。全省拥有从热带、亚热带、温带到寒带等多种气候类型,几乎囊括我国所有的生态系统类型(杨宇明等,2008)。由于云南特殊的地势地貌和多样化的生物气候类型,使得外来植物的各种气候适应性能力都能得到体现。本研究虽然以肿柄菊在云南的分布情况进行分析,但其气候的适应性也可以作为在全国分布潜力的推断依据。气温和降雨是影响植物分布的两个主要因素,无论是模型预测还是当前的实际分布情况,肿柄菊能分布最冷的区域为北亚热带地区,很显然肿柄菊再向北更冷的区域扩散时将会受到低温的限制。肿柄菊在云南已有的分布区年降雨量最少区域仅为578 mm(王四海等,2019),我国600 mm等降雨量线大致沿青藏高原、黄土高原至华北地区,这个范围远超过肿柄菊所能向北分布的界限,从区域尺度看降雨不会是肿柄菊向北分布的限制因子。肿柄菊在云南能分布地区的最冷月份(一月)平均气温约8 ℃,如果以最冷月(一月)平均气温8 ℃推断,肿柄菊在我国大致分布在广西、广东和福建及其以南地区,这一分布范围大致与世界较适宜分布区域沿我国南部地区分布范围基本相当。
外来植物经过对入侵地环境的长期适应有时会发生生态位偏移,所以在某种程度上外来植物入侵范围的精确预测具有困难(Guisan et al., 2014;李委涛等,2014)。原产于中美洲的紫茎泽兰(Ageratina adenophora)、飞机草(Eupatorium odoratum)2种外来植物像肿柄菊一样都是于20世纪30年代在云南建立了逃逸种群(薛纪如等,1979;王四海等,2008),当前紫茎泽兰已扩散到西藏、四川、重庆和湖北等地(王翀等,2014),而飞机草仅限于在云南中南部、广西西部和南部、广东和福建南部等地区分布(杨波等,2009),两种植物原产地和引入时间虽相似,但入侵潜力具有很大的不同。在21世纪初,飞机草在云南的入侵范围与肿柄菊大致相同(王四海等,2004),但当前肿柄菊入侵的海拔高度和向北的扩散范围都大于飞机草(朱枫等,2018),表现出较强的入侵态势。紫莖泽兰能耐受-5 ℃的低温,飞机草在0 ℃时叶片和枝条可冻死(吴仁润等,1984),肿柄菊根据目前分布情况推测在冬季可以忍耐短期0 ℃以下低温,这三种植物在耐冻性方面的差异决定了它们潜在分布区的不同。另外,即使外来入侵植物没有发生生态适应性进化,它们也可以占据与原产地不同的、非一致性的气候生态位(Datta et al., 2019),这也可以解释一些外来植物原产地相同,但入侵范围可能不同的原因。综合考虑到肿柄菊入侵气候生态位保守性(Obiakara & Fourcade, 2018)、入侵时滞较长(王四海等,2008),以及全球气温变化因素,经过较长时间的扩散和适应,肿柄菊也有可能向北零星扩散到一月平均温度小于8 ℃的低适生区。虽已有标本记载在广西、广东、福建、海南等地有肿柄菊分布,但对于其大面积密集分布并带来危害性的报道并不多。为了预防肿柄菊在我国南方地区大面积扩散带来危害,有必要对其入侵和扩散态势进行监测(李委涛等,2014)。
参考文献:
CHEN J, WANG SH, YANG W, et al., 2020. Community dynamics of an alien invasive plant species Tithonia diversifolia [J]. Chin J Ecol, 39(2):469-477. [陈剑, 王四海, 杨卫, 等, 2020. 外来入侵植物肿柄菊群落动态变化特征 [J]. 生态学杂志, 39(2): 469-477.]
CHEN ZY, 2001. Climate of Yunnan [M]. Beijing: China Meteorological Press. [陈宗瑜, 2001. 云南气候总论 [M]. 北京: 气象出版社.]
DATTA A, SCHWEIGER O, KHN I, 2019. Niche expansion of the invasive plant species Ageratina adenophora despite evolutionary constraints [J]. J Biogeogr, 46: 1306-1315.
ELITH J, KEARNEY M, PHILLIPS S, et al., 2010. The art of modeling range-shifting species [J]. Mothods Ecol Evol [J], 1: 330-342.
GUISAN A, PETITPIERRE B, BROENNIMANN O, et al., 2014. Unifying niche shift studies: insights from biological invasions [J]. Trend Ecol Evol, 29(5): 260-269.
JAMA B, PALM CA, BURESH RJ, et al., 2000. Tithonia diversifolia as a green manure for soil fertility improvement in western Kenya: A review [J]. Agrofor Syst, 49(2):201-221.
LENTZ DL, BYE R, SNCHEZ-CORDERO V, 2008. Ecological niche modeling and distribution of wild sunflower in Mexico [J]. Int J Plant Sci, 169(4): 541-549.
LI WT, ZHENG YL, FENG YL, 2014. Differences in growth traits between Chromolaena odorata plants from invasive and native populations [J]. Acta Ecol Sin, 34(23): 6890-6897. [李委涛, 郑玉龙, 冯玉龙, 2014.飞机草入侵种群与原产地种群生长性状的差异 [J]. 生态学报, 34(23): 6890-6897.]
LIU XT, YUAN Q, NI J, 2019. Research advances in modeling plant species distribution in China [J]. Chin J Plant Ecol, 43(4): 273-283. [刘晓彤, 袁泉, 倪健, 2019. 中国植物分布模拟研究现状 [J]. 植物生态學报, 43(4): 273-283.]
LUO Y, LIU Z, ZHAO JH, et al., 2009. Analysis of mineral elements and quality evaluation of Tithonia diversifolia(Hemsl.) A. Gray. [J]. Trop Agric Eng, 33(3): 36-39. [罗瑛, 刘壮, 赵君华, 等, 2009. 肿柄菊的矿质营养元素分析及评价 [J]. 热带农业工程, 33(3):36-39.]
OBIAKARA MC, FOURCADE Y, 2018. Climatic niche and potential distribution of Tithonia diversifolia (Hemsl.) A. Gray in Africa [J]. PLoS ONE, 13(9): e0202421.
SWETS JA, 1988. Measuring the accuracy of diagnostic systems [J]. Science, 240(4857): 1285-1293.
URBANI F, D′ALESSANDRO P, BIONDI M, 2017. Using Maximum Entropy Modeling (MaxEnt) to predict future trends in the distribution of high altitude endemic insects in response to climate change [J]. B Insectol, 70(2): 189-200.
VELOZ SD, 2009. Spatially autocorrelated sampling falsely inflates measures of accuracy for presence-only niche models [J]. J Biogeogr, 36(12): 2290-2299.
WANG C, LIN HL, HE L, et al., 2014. Research on responses of Eupatorium adenophorums potential distribution to climate change [J]. Acta Pratacult Sin, 23(4): 20-30. [王翀, 林慧龙, 何兰, 等, 2014. 紫茎泽兰潜在分布对气候变化响应的研究 [J]. 草业科学, 23(4): 20-30. ]
WANG SH, CHEN J, LI NY, et al., 2019. Factors affecting seed production of Tithonia diversifolia, an invasive alien plant [J]. Ecol Environ Sci, 28(7): 1369-1378. [王四海, 陈剑, 李宁云, 等, 2019. 外来植物肿柄菊结籽量影响因素分析 [J]. 生态环境学报, 28(7): 1369-1378.]
WANG SH, SUN WB, CHENG X, 2004. Attributes of plant proliferation, geographic spread and the natural communities invaded by the naturalized alien plant species Tithonia diversifolia in Yunnan,China [J]. Acta Ecol Sin, 24(3): 444-449. [王四海, 孙卫邦, 成晓, 2004. 逃逸外来植物肿柄菊在云南的生长繁殖特性、地理分布现状及群落特征 [J]. 生态学报, 24(3): 444-449.]
WANG SH, SUN WB, CHENG X, et al., 2008. Reproductive characteristics of Tithonia diversifolia and its geographical spread in Yunnan Province of South-West China [J]. Acta Ecol Sin, 28(3): 426-432. [王四海, 孙卫邦, 成晓, 等, 2008. 外来植物肿柄菊(Tithonia diversifolia)的繁殖特性及其地理扩散 [J]. 生态学报, 28(3): 426-432.]
WU RR, ZHANG DY, LU XS, 1984. On the distribution, perniciousness and control of the two weeds,Chromolaena coelestina and C. odorata in Yunnan Province of China [J]. Grassland Chin, 2: 17-22. [吴仁润, 张德银, 卢欣石, 1984. 紫茎泽兰和飞机草在云南的分布、危害与防治 [J]. 中国草原, 2: 17-22.]
XU DP, ZHUO ZH, WANG RL, et al., 2019. Modeling the distribution of Zanthoxylum armatum in China with MaxEnt modeling [J]. Global Ecol and Conserv, 19: e00691.
XUE JR, DONG SC, YIN JH, et al., 1979. Adaptability and damage situation of Eupatorium adenophorum and control measures [J]. Acta Bot Yunnan, 1(1): 106-114. [薛纪如, 董世仁, 印嘉祜, 等, 1979. 紫茎泽兰的适应性能、危害情况及防除措施 [J]. 云南植物研究, 1(1): 106-114.]
XU ZL, PENG HH, PENG SZ, 2015. The development and evaluation of species distribution models [J]. Acta Ecol Sin, 35(2): 557-567. [许仲林, 彭焕华, 彭守璋, 2015. 物种分布模型的发展及评价方法 [J]. 生态学报, 35(2): 557-567.]
YANG B, XUE YG, TANG XF, et al., 2009. Predicting potential geographic distribution of Eupatorium odoratum in China [J]. Plant Protect, 35(4): 70-73. [杨波, 薛跃规, 唐小飞, 等, 2009. 外来入侵植物飞机草在中国的适生区预测 [J]. 植物保护, 35(4): 70-73.]
YANG J, TANG L, GUAN YL, et al., 2012. Genetic diversity of an alien invasive plant mexican sunflower (Tithonia diversifolia) in China [J]. Weed Sci, 60(4): 552-557.
YANG YM, WANG J, WANG JH, et al., 2008. Studies on the biodiversity and its conservation in Yunnan, China [M]. Beijing: Science Press. [杨宇明, 王娟, 王建皓, 等, 2008. 云南生物多樣性及其保护研究 [M]. 北京: 科学出版社.]
ZHU F, WANG SH, CHEN J, et al., 2018. Distribution characteristics and influencing factors of Tithonia diversifolia, an alien invasive plant species, in Yunnan Province, China [J]. Chin J Ecol, 37(9): 27-34. [朱枫, 王四海, 陈剑, 等, 2018. 外来入侵植物肿柄菊在云南的分布特征及其影响因素 [J]. 生态学杂志, 37(9): 27-34.]
(责任编辑 何永艳)
收稿日期: 2020-03-06
基金项目: 国家自然科学基金(31560092);云南省森林植物培育与开发利用重点实验室开放基金项目( ZZCX2016-10);云南省技术创新人才培养资助项目(2019HB066) [Supported by the National Natural Science Foundation of China (31560092); Open Fund of Yunnan Key Laboratory of Forest Plant Cultivation and Utilization (ZZCX2016-10); Talent Training Program on Technical Innovation of Yunnan Province (2019HB066)]。
作者简介: 陈剑(1978-),硕士,助理研究员,主要从事林业可持续发展研究,(E-mail)chenjianyaf@126.com。
通信作者: 王四海,博士,副研究员,主要从事植物保护与利用研究,(E-mail)wangsh6688@163.com。
