桂林岩溶石山落叶阔叶林种类组成及其环境解释
2021-08-16林红玲梁士楚姚义鹏姜勇包含农彩汪宋静黄佳甘鑫梅
林红玲 梁士楚 姚义鹏 姜勇 包含 农彩汪 宋静 黄佳 甘鑫梅


摘 要: 為了解落叶阔叶林森林植被的群落类型和组成结构,该文在桂林阳朔县和灵川县岩溶生境典型的落叶阔叶林内设置49个20 m × 20 m的随机样方,在野外调查基础上进行群落数量分类和排序,对其群落类型进行划分,并对不同类型的群落种类组成、结构及其分布与环境因子的耦合关系进行分析。结果表明:(1)桂林岩溶石山落叶阔叶林可划分为3个群丛组类型,即群丛组A:南酸枣-喜树+巴豆群丛组(ASS. Choerospondias axillaris-Camptotheca acuminata+Croton tiglium),群丛组B:光皮梾木-喜树群丛组(ASS. Swida wilsoniana-Camptotheca acuminata),群丛组C:麻栎-檵木群丛组(ASS. Quercus acutissima-Loropetalum chinense)。(2)不同群落类型中常绿和落叶物种的丰富度、多度、胸高断面积及重要值的变化:在乔木层中,3个群丛组均为落叶物种占优势;在灌木层中,群丛组A、B以落叶物种占优势,群丛组C以常绿物种占优势。(3)在1~5 cm和5~10 cm径级内,群丛组A、B的落叶物种在4个度量指标上均大于常绿物种,群丛组C除物种丰富度外,均为常绿物种大于落叶物种;在 ≥10 cm径级中,除群丛组B的物种丰富度外,3个群丛组中的各项指标均为落叶物种占主导地位。(4)岩石裸露率、海拔、坡向、土壤含水量、土壤pH、土壤有机质、全氮、速效氮、速效钾、林冠开阔度和人为干扰是影响研究区不同类型群落分布的主要环境因子。
关键词: 群落类型, 落叶阔叶林, 环境因子, 数量分类, 岩溶石山
中图分类号: Q948
文献标识码: A
文章编号: 1000-3142(2021)05-0758-11
Species composition and environmental analysis of
deciduous broad-leaved forests in karst hills of Guilin
LIN Hongling1,2, LIANG Shichu1,2, YAO Yipeng3, JIANG Yong1,2*, BAO Han1,2,
NONG Caiwang1,2, SONG Jing1,2, HUANG Jia1,2, GAN Xinmei1,2
( 1. Key Laboratory of Wild Animal and Plant Ecology Guangxi Colleges and Universities, Guangxi Normal University, Guilin 541006,
Guangxi, China; 2. College of Life Sciences, Guangxi Normal University, Guilin 541006, Guangxi, China; 3. Guilin
University of Aerospace Technology, Guilin 541004, Guangxi, China )
Abstract: Deciduous broad-leaved forests are one of the typical forest vegetation types in the subtropical karst hills. In order to understand the community type and composition structure of this type of forest vegetation, we established forty-nine 20 m × 20 m random plots in deciduous broad-leaved forests with typical karst habitats in Yangshuo County and Lingchuan County of Guilin. We conducted an in-depth analysis of the coupling relationship from different community types, structures, and environmental factors. The results were as follows: (1) The deciduous broad-leaved forests in the karst hills could be classified into three community types based on quantitative methods, Ass. Choerospondias axillaris-Camptotheca acuminata+ Croton tiglium (Association Group A), Ass. Swida wilsoniana-Camptotheca acuminata (Association Group B), Ass. Quercus acutissima-Loropetalum chinense (Association Group C). (2) The species richness, species abundance, basal area, and the importance value for evergreen and deciduous species were different for each community type. Deciduous species were dominant in the tree layer for all three association groups. Shrub layer for Association Group A and Group B were also dominated by deciduous species, while Association Group C was dominated by evergreen species. (3) The species richness, species abundance, basal area and importance value of deciduous species in Association Group A and Group B were greater than evergreen species within the 1-5 cm and 5-10 cm diameter classes. In Association Group C, all indicators were greater for evergreen species than deciduous species for the 1-5 cm and 5-10 cm diameter classes. For the ≥10 cm diameter class, deciduous species were dominant for all indicators in all three association groups except for the species richness in Association Group B. (4) The rock bare rate, soil water content, elevation, aspect, pH, soil organic matter, total nitrogen, available nitrogen, available potassium, canopy openness and human disturbance gradient were the dominant factors affecting the distribution of different types of communities in the study.
Key words: community type, deciduous broad-leaved forests, environmental factors, quantitative classification, karst hills
植物群落的数量分析作为植被生态学研究的重要手段之一,能够合理和准确地揭露植物组成种类、植物群落和植被与环境之间的复杂生态关系(苏日古嘎等,2010)。关于群落性质,植物生态学家一直存在着两种不同的观点。“有机体派”学者认为,植物群落是自然单位,因具备明确的边界而与其他群落间断、可分,可以像物种那样分类;而“个体派”学者认为,群落和生境是连续的,他们采取生境梯度分析的方法,运用排序的手段来研究连续群落的变化。而实际上,植物群落的间断性和连续性是并存的。群落的间断性可用分类来揭示,而连续性用排序来揭示(梁士楚,2015)。目前,国际上常采用双向指示种分析法(TWINSPAN)与除趋势对应分析法(DCA)相结合来进行数量分类和排序,对于分类结果而言,这两种方法的结合使用不仅起到了交叉验证的作用,而且还更深刻地反映了植物群落及环境之间的生态关系(张金屯,2018)。
落叶阔叶林(deciduous broad-leaved forests)是温带地带性植被类型,主要分布在中纬度地区(梁士楚,2015);落叶阔叶林是组成亚热带海拔较高的山地垂直植被带谱的重要类型(刘亚兰等,2010)。它具有明显的季相变化,其最明显的特征表现为树木的生长仅在温暖的季节,树叶在入冬前枯死并凋落。树木可以通过落叶来躲避极端生境对自身生长造成的伤害和不利影响,可以说是物种适应环境胁迫的一种重要手段(Poorter & Markesteijn, 2008)。在大范围的地理学尺度上,虽然决定树木落叶的最主要因素是气候条件(丁易和臧润国,2008),但是在小范围的局部尺度下,环境异质性和森林演替状况是影响群落中落叶树种数量的重要因素之一。例如,在中亚热带石灰岩地区,植物常常由于土壤水分的缺乏和低温等胁迫而导致落叶(Liu et al., 2020)。落叶树种对于整个群落结构、物种组成及更新、养分和物质的循环等方面具有重要意义。例如,落叶树种在落叶季节能增加林下光照,使原有的物种种类组成和更新情况得以改变,不仅增加了植物群落对光照、空间、营养物质的利用能力,而且还提高了群落中的生物多样性(林庆凯,2017)。
桂林位于广西壮族自治区东北部,具有典型的岩溶地貌特征,石质荒漠化程度高且生态环境极其脆弱(袁道先和章程, 2008),具有景观异质性高、水文地质结构明显、岩石裸露率高、碳酸盐溶蚀性强和土壤贫瘠等特征(胡芳等,2018)。而落叶阔叶林是亚热带岩溶石山的主要群落组类型之一(苏宗明和李先琨,2003),它是在岩溶地区特殊生境下所形成的一种典型的森林植被类型,在岩溶森林生态系统的恢复和保护中具有难以替代的作用和地位,对防止岩溶石山石质荒漠化、调节表层岩溶带的蓄水功能、调节森林碳库等多项生态功能发挥重要作用(李梦德,2016)。因此,本文以桂林岩溶石山落叶阔叶林为研究对象。首先,运用TWINSPAN分类和DCA排序对其群落类型进行科学的划分。其次,分析不同群落类型的群落种类组成和结构,运用冗余分析的方法对不同群落类型与环境因子的耦合关系进行分析,以解决以下科学问题:(1)不同群落类型的物种组成和群落结构是怎样的?(2)影响不同群落类型变化和分布的环境因子是什么?合理解答这两个科学问题,可以揭示桂林岩溶石山落叶阔叶林植物群落与环境之间的关系,从而为探索岩溶石山落叶阔叶林的物种共存及其维持机制,以及为岩溶植被资源的管理与保护和退化植被生态修复提供科学合理的依据。
1 材料与方法
1.1 研究区概况
研究区位于广西壮族自治区东北部的桂林市阳朔县葡萄镇禄迪村(110°22′59.59″E、24°57′14.28″ N)和灵川县辇田尾村(110°26′5.88″ E、25°18′34.92″ N),其村落后山均为典型的裸露型岩溶地貌,具有土层薄、蓄水能力弱、碳酸钙含量高及环境容纳量低等生境特点(李月娟等,2019)。气候类型属于中亚热带湿润气候,全年雨水充裕,主要集中在夏季,年降雨量为1 814~1 941 mm,全年光照充足,年蒸发量为1 337~1 857 mm,年平均湿度为73%~79%。年平均气温为17.4~21 ℃,年极端最低和最高气温分布范围为-3.9~1.3 ℃和37.6~39 ℃(梁士楚等,2018)。
1.2 样地的建立与调查方法
在充分踏查的基础上,于2019年7—9月在桂林市区阳朔县禄迪村和灵川县辇田尾村的岩溶石山落叶阔叶林内,选择49个长和宽各为20 m的样方进行群落学调查,其中禄迪村设置了24个随机样方,辇田尾村设置了25个随机样方。对样方内所有胸径(DBH)≥1 cm 的乔木、灌木和木质藤本等木本植物个体开展每木检尺,记录内容包括物种名称、个体在样方内的相对坐标,其中乔木记录物种的胸径和基径,灌木记录物种的基径。利用精度为0.01的可触式智能双星导航仪(Touch35),获取海拔、坡向、坡度、经纬度的数据,同时记录岩石裸露率环境特征。本次调查共记录到的木本植物82种,隶属于37科70属,其中常绿植物37种、落叶植物45种。
1.3 土壤样品的采集与测定
利用田字法将每個样方分割成16个长、宽各为5 m的小样方,清除土壤表层的枯枝落叶等杂质后,运用环刀法采集土样,并测定土壤含水量(soil water content, SWC, %);采用五点法采集表层的土壤样品,待实验室风干处理后进行土壤养分指标的测定。测定土壤pH值(soil pH)、土壤有机质(soil organic matter, SOM, g·kg-1)、全氮(total nitrogen, TN, g·kg-1)、速效氮(available nitrogen, AN, mg·kg-1)、速效钾(available potassium, AK, mg·kg-1)5个指标。土壤养分指标的测定方法均详见鲍士旦(2000)。对每份土壤样品平行测定3次,取平均值作为本研究的土壤环境因子分析的基础数据。
1.4 数据处理
首先,以样方为单位分别计算乔木和灌木的物种重要值。计算公式(梁士楚,2015;刘润红等,2019):乔木重要值=(相对多度+相对显著度+相对频度)/3;灌木重要值=(相对多度+相对频度)/2。其次,基于物种在样方中的重要值,利用R软件中twinspanR程序包中的twinspan函数与vegan程序包中的decorana函数进行双向指示种分析和DCA排序。为了更好地比较不同群落类型的物种组成和群落结构特征,我们选择物种的丰富度、多度、胸高断面积、重要值指标作为度量标准,将植物个体胸径(DBH)划分为小径级(1 cm≤DBH<5 cm)、中径级(5 cm≤DBH<10 cm)和大径级 ( ≥10 cm) 3个等级(龙文兴,2011;黄永涛,2015)。为了探究海拔、坡向、岩石裸露率、林冠开阔度、人为干扰、土壤pH、土壤有机质、全氮、速效氮和速效钾等环境因子对物种分布是否具有显著的影响,我们采用RDA排序(Canonical Correlation Analysis)结合随机化排列分析两者之间的关系,通过vegan包中的envfit函数,进行蒙特卡洛置换检验(随机模拟999次)检验环境因子的显著性。将不同环境因子变量对物种分布的影响进行变差分解,得到不同环境因子以及环境因子之间的解释能力。其中,海拔为样方4个顶点及样方中心位置的海拔平均值;岩石裸露率为样方内裸露于地表的基岩面积与样方面积的比值(Nee, 2005),人为干扰梯度以数字等级来划分,分为1~3个等级,数字越大,表示干扰程度越严重;在每个20 m × 20 m的样方中心位置,使用外带180°鱼眼镜头变换器的数码相机(Canon EOS 5D),在离地面1.3 m处向上对林分冠层进行拍摄取像,运用Gap Light Analyzer (GLA)分析处理图像计算林冠开阔度(CO, %)(朱教君等,2005;李根柱等,2009)。统计分析与绘图均在R 3.6.3软件中进行。
2 结果与分析
2.1 群落类型及其物种组成
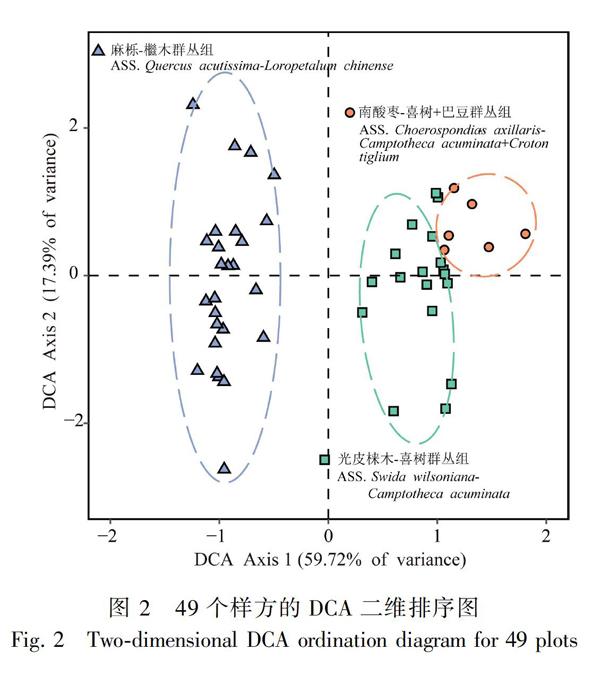
应用TWINSPAN分类结合DCA排序的结果(图1,图2)表明,在落叶阔叶林内共调查49个样方可以被划分为3个群丛组类型。
群丛组A:南酸枣-喜树+巴豆群丛组(ASS. Choerospondias axillaris-Camptotheca acuminata+Croton tiglium),包括样方1~5和样方7,共6个。该群丛组位于阳朔县禄迪村,乔木层优势种为南酸枣(Choerospondias axillaris),伴生种有光皮梾木(Swida wilsoniana)、巴豆(Croton tiglium)、朴树(Celtis sinensis)等,灌木层优势种为喜树(Camptotheca acuminata)、巴豆,其他伴生种有朴树、牡荆(Vitex negundo var. Cannabifolia)、薄叶鼠李(Rhamnus leptophylla)和柞木(Xylosma congesta)等。群丛组B:光皮梾木-喜树群丛组(ASS. Swida wilsoniana-Camptotheca acuminata),包括样方6和样方8~24,共18个。该群丛组位于阳朔县禄迪村,乔木层优势种为光皮梾木,伴生种有南酸枣、枫香(Liquidambar formosana)、朴树等,灌木层优势种为喜树,其他伴生种有光皮梾木、红背山麻杆(Alchornea trewioides)、牡荆、柞木和干花豆(Celtis sinensis)等。群丛组C:麻栎-檵木群丛组(ASS. Quercus acutissima-Loropetalum chinense),包括样方25~49,共25个。该群丛组位于灵川县辇田尾村,乔木层优势种为麻栎(Quercus acutissima),伴生种有光蜡树(Fraxinus griffithii)、枫香、花榈木(Ormosia henryi)等,灌木层优势种为檵木(Loropetalum chinense),其他伴生种有粗糠柴(Mallotus philippensis)、柃木(Eurya japonica)、光蜡树和短序荚蒾(Viburnum brachybotryum)等。
2.2 群落物种组成结构及其变化
如图(3:A-D)所示,在乔木层,群丛组A、B、C的物种丰富度、多度、胸高断面积和重要值在4个度量指标上均表现为落叶物种大于常绿物种,乔木层个体高度分布区间3~24 m,以落叶物种占主导地位,主要优势物种包括光皮梾木、南酸枣、麻栎、枫香、光蜡树和朴树等。在灌木层,群丛组A、B的落叶物种在这4个指标上均大于常绿物种,灌木个体高度分布区间0.75~3 m,主要由落叶物种喜树、光皮梾木、干花豆和常绿物种粗糠柴、巴豆和柞木组成。群丛组C则表现出相反的变化趋势,即在4个度量指标上常绿物种所占比例大于落叶物种,常绿物种檵木占优势地位,其次是粗糠柴、柃木、短序荚蒾和落叶物种光蜡树。
从物种丰富度方面来看,除了群丛组B≥10 cm的径级外,其余均为落叶物种占优势(图4:A)。从多度、胸高断面积和重要值3个指标(图4:B-D)来看,除了群丛组C的1~5 cm径级和5~10 cm径级内常绿物种比例大于落叶物种外,其余均显示为落叶物种大于常綠物种。
2.3 群落物种组成与环境因子的关系
采用物种多度和环境因子两个数据矩阵,对桂林岩溶石山落叶阔叶林进行RDA排序,前两轴的累计解释率达到53.7%,包含了11个环境因子的大部分信息解读;采用前两轴绘制RDA二维排序图,结果如图5所示。49个样方在排序图中分布结果与前文数量分类的研究结果相符,反映了3种不同群丛组类型所处的环境梯度。第一排序轴主要反映了植物群落分布格局在土壤养分含量、岩石裸露率、土壤含水量等环境因子梯度上的变化,第二排序轴主要反映的是干扰等环境因子在梯度上的变化。综合表1的结果看,岩石裸露率、海拔、坡向、土壤含水量、土壤pH、土壤有机质、全氮、速效氮、速效钾、林冠开阔度和人为干扰是影响研究区不同类型群落分布的主要环境因子。
环境因子的变差分解结果(图6)显示,11个环境因子对物种分布的解释能力为64.3%。其中,地形因子占10.4%,土壤因子占7.5%,干扰因子占0.8%,地形因子和土壤因子耦合作用的解释率占41.3%,土壤因子和干扰因子耦合作用的解释率占0.6%,干扰因子和地形因子耦合作用的解释率占1.4%,三者共同作用的解释率占2.3%,未能解释的部分比例占35.7%。
3 讨论与结论
3.1 岩溶石山落叶阔叶林数量分类与排序
数量分类和排序是研究植被生态学最为常用的数量分析手段。迄今为止,许多学者通过对各种数量分类方法的对比和研究应用,均认为理论上较为完善和应用效果较好的方法是双向指示种分析法(TWINSPAN)和除趋势对应分析法(DCA)(孙小伟,2018)。前者作为目前群落分类史上较为理想的分类方法之一,采用数字量化样方的方法直接给出具体群落的分类个数,同时以此为依据得出详细的优势群落类型,具有简单直接,通俗易懂的分类特点。后者利用样方间在物种组成上的相似性,将物种进行排列使其反映出一定的生态梯度,产生直观可视的二维分类排
序图(Braak & Smilauer, 2012;张金屯,2018)。因此,二者的结合使用可以更准确地相互印证、相互补充、相互完善,能够让我们得到更加符合野外实际情况的解释,客观准确地揭示出植被、植物群落与植物群落和环境因子的生态关系(黄永涛,2015;张金屯,2018)。本研究通过二者结合的方法将桂林岩溶石山落叶阔叶林内的49个样方分为3个群丛组,分类结果在DCA可视化二维图上清晰的表达出来。本研究分析结果系统地反映了岩溶石山落叶林群落类型的多样性,同时进一步了解了群落间的物种分布随环境因子梯度变化的替换过程。桂林坐落于典型岩溶石山地貌区, 是广西区岩溶石山分布的主要地区之一,生境异质性高,使得该地区的植物群落结构多样、物种组成丰富(宋同清等,2010;胡芳等,2018)。本研究依据排序与分类的结果显示,与实际调查结论基本吻合,证明数量化分析方法的结果是可信的。
3.2 物种组成和群落结构与环境之间的关系
本研究中,变差分解结果表明环境是影响研究区物种分布的主要原因,而RDA 排序的结果也较好地揭示了桂林岩溶石山落叶阔叶林植物群落中组成的物种差异以及分布与环境因子之间的相互关系,具有一定的生态学意义。生态学家们普遍认为,常绿与落叶物种由于在资源利用方式上有着不同的偏好,所以在生境的选择上可能有所差异。群丛组A分布在土壤水分含量低、干扰程度较低以及岩石裸露率、土壤pH、土壤养分含量、林冠开度较高的阳坡;群丛组B分布在土壤水分量低以及土壤pH、岩石裸露率、土壤养分含量、干扰程度较高的阳坡;群丛组C则是分布在土壤pH、岩石裸露率、林冠开度、土壤养分含量相对偏低以及土壤水分含量和干扰程度较高的区域。
群落的垂直结构指的是群落在垂直方向上的配置状态,对于了解群落更新和动态特征有着重要的作用(杨晓东,2014; 梁士楚,2015)。本研究发现,在垂直结构上,A、B、C 3个群丛组乔木层的落叶物种在我们选择的4个度量指标中均大于常绿物种,其中优势物种均为高大的阳性乔木;而常绿物种在乔木层中处于较低的亚层处,这些物种在乔木层中个体数较少,多以小乔木和大型灌木的形式存在,导致常绿物种不能在乔木层占据优势地位。在光照条件较好的区域,乔木层落叶物种种类更多,而落叶树物种在乔木层占优势,往往影响着林下植物的光照条件(Chabot & Hicks, 1982; Quigley & Platt, 2003)。与常绿物种相比,落叶物种更适合生长在光照较强和土壤养分较高的环境,因为其采取高生长的生态策略,对资源的利用效率高,所以在生长季节能快速获取更多的光照、水分和营养物质等资源以满足自身的快速生长(白坤栋等,2015)。此外,落叶物种还能以落叶的方式克服极端生境对树木生长造成的危害和不良影响(Pooter & Markesteijn, 2008; 黄永涛, 2015),因此其在与常绿物种竞争的过程中占优势地位。在我们的调查中发现,首先,群丛组A和群丛组B位于海拔较高的阳坡生境,随着海拔梯度的升高,光照强度逐渐增加,适合的水热条件促进了落叶物种的生长。其次,从物种组成以及多度来看,占据乔木层优势地位的落叶物种南酸枣、光皮梾木和光蜡树等均是岩溶石山区典型的先锋物种(司彬,2007),并且样地内树木的胸径偏低,矮小植物的个体数量多,高大植物个体的数量相对偏少。这些现象都表明群丛组A、B处于干扰后自然恢复初期阶段,群丛组C处于扰后自然恢复中期阶段。在灌木层中,群丛组A、B在丰富度等4个度量指标均表现为落叶物种大于常绿物种,而群丛组C则相反。群丛组A的优势种以落叶物种喜树为主,其次是常绿物种巴豆和落叶物种光皮梾木;而群丛组B的优势种以落叶物种喜树和光皮梾木为主,其次是常绿物种粗糠柴。这两个群丛组的灌木层都是落叶物种占优势,其中光皮梾木和喜树均以更新苗的形式存在,群落的天然更新状态良好。群丛组C的优势种以常绿物种檵木为主,在灌木层中占据绝对的优势地位,其次是落叶物种光蜡树。由于群丛A、B位于林冠开度(CO)較大的阳坡生境,乔木层主要以落叶物种占主导地位。因此,与群丛组C位于海拔偏低的阴坡生境相比,较强的林下光照对于灌木层中阳生性落叶物种的萌发和生长起了促进作用,而中生耐阴常绿植物的萌发和生长则受到了抑制,导致了喜光落叶物种的增加。尽管群丛组C林内光照条件相对较差,但适宜的低光生境促进了常绿物种在灌木层中迅速生长的格局。
径级结构通常作为衡量植物群落生长发育和演替状况的重要指标(陈列,2014;伏捷等,2018)。群丛组A、B的3个径级区间的落叶物种在4个量度指标上均大于常绿物种,群丛组C内中小径级区间的常绿物种在3个度量指标上大于落叶物种(物种丰富度除外),但是在径级≥10 cm个体上4个度量指标都呈现相反的变化趋势,即落叶物种大于常绿物种。导致这样的分布格局不仅与常绿和落叶物种的生活型属性有关,而且还与群落类型所处的干扰后自然恢复所处阶段以及相对应的环境梯度的改变,如土壤养分含量、地形因子、干扰等组合的异质性生境有关(丁易和臧润国,2008;康敏明,2010)。本研究区3个群丛组内,小中径级个体主要是由小乔木和灌木构成,由于构成群丛组A、B的小乔木和灌木物种主要是落叶物种,因此出现了中小径级内落叶物种在4个度量指标上占比高的现象。而群丛组C内小乔木和灌木物种是以常绿树种为主,在中小径级内除物种丰富度外,在其余3个指标上均展示出常绿物种大于落叶物种的变化趋势。丁易和臧润国(2008)对海南岛热带低地雨林的研究表明,程度较高干扰可能是导致群落中落叶树种数量增加的原因之一;黄永涛(2015)对湖北星斗山和木林子两个国家级自然保护区的常绿落叶阔叶混交林的研究表明,干扰能够显著地改变群落结构和物种组成,从而影响群落发展的方向。群丛组A、B的中小径级在各个指标上都是落叶树种占优势,这说明人为干扰在一定程度上影响着群落的更新和演替,使得长期以内落叶物种都会在这两个群丛组中占据主导地位。群丛组C的常绿物种在中小径级区间占据了优势地位,暗示在未来的资源竞争中,常绿物种将在群落中占主导,具有逐渐发展到乔木层替换部分落叶物种的演替趋势,进而完成岩溶石山落叶林向常绿落叶阔叶混交林的演替发展过程(欧祖兰等,2005)。
3.3 结论
(1)桂林岩溶石山落叶阔叶林49个样方可划分3个群丛组类型,即(A)南酸枣-喜树+巴豆群丛组、(B)光皮梾木-喜树群丛组、(C)麻栎-檵木群丛组。(2)垂直结构上,在乔木层中,3个群丛组均是落叶物种占优势;在灌木层中,群丛组A和群丛组B以落叶物种占优势,群丛组C则以常绿物种占优势。径级结构上,在1~5 cm和5~10 cm径级内,群丛组A和群丛组B均是落叶物种占优势,群丛组C除了物种丰富度以外,均是常绿物种占据优势地位;在≥10 cm径级中,除了群丛组B的物种丰富度以外,3个群丛组中的各项指标均为落叶物种占主导地位。(3)冗余分析的结果表明,岩石裸露率、土壤含水量、海拔、坡向、土壤pH、土壤有机质、全氮、速效氮、速效钾、林冠开度和人为干扰都是影响研究区不同类型群落分布的主要环境因子。
本研究结果既为进一步探讨岩溶石山落叶阔叶林内不同落叶树种以及落叶树种与常绿树种共存机制以及落叶阔叶林向常绿落叶阔叶混交林进展演替过程奠定了基础,也进一步丰富了桂林岩溶石山森林植被生态学研究的资料,可为岩溶石山森林生物多样性保护管理提供参考依据。
参考文献:
BAI KD, MO L, LIU M, et al., 2015. Nutrient resorption patterns of evergreen and deciduous tree species at different altitudes on Maoer Mountain, Guangxi [J]. Acta Ecol Sin, 35(17): 5776-5787. [白坤栋, 莫凌, 刘铭, 等, 2015. 广西猫儿山不同海拔常绿和落叶树种的营养再吸收模式 [J]. 生态学报, 35(17): 5776-5787.]
BAO SD, 2000. Soil agro-chemistrical analysis [M]. 3rd ed. Beijing: China Agriculture Press. [鲍士旦, 2000. 土壤农化分析 [M]. 3版. 北京: 中国农业出版社.]
Biodiversity Committee, Chinese Academy of Scienses, 1994. Principles and methodologies of biodiversity studies [M]. Beijing: Science and Technology of China Press. [中国科学院生物多样性委员会, 1994. 生物多样性研究的原理与方法 [M]. 北京: 中国科学技术出版社.]
BRAAK CJFT, SMILAUER P, 2012. Canoco reference manual and users guide: Software for ordination, version 5 [M]. Ithaca NY: Microcomputer Power.
CHABOT BF, HICKS DJ, 1982. The ecology of leaf life spans [J]. Ann Rev Ecol Syst, 13(1): 229-259.
CHEN L, 2014. Population structure of main tree species and the response of tree radial growth toclimte in broadleaved Korean pineforests on Changbai Mountain [D]. Beijing: Beijing Forestry University. [陈列, 2014. 长白山阔叶红松林主要树种种群结构及其林木径向生长对气候响应 [D]. 北京: 北京林业大学.]
DING Y, ZANG RG, 2008. Changes in deciduous trees during recovery of tropical lowland rain forests on abandoned shifting cultivation lands in Hainan Island, South China [J]. Biodivers Sci, 16(2): 103-109. [丁易, 臧潤国, 2008. 海南岛热带低地雨林刀耕火种弃耕地恢复过程中落叶树种的变化 [J]. 生物多样性, 16(2): 103-109.]
FU J, ZHANG H, HE H, et al., 2019. Analyses on structure and succession trend of forest communities in paleo-stone stream of mountainous area of eastern Liaoning [J]. J Plant Resour Environ, 28(2): 1-9. [伏捷, 张华, 何红, 等, 2019. 辽东山区古石河森林群落结构及演替趋势分析 [J]. 植物资源与环境学报, 28(2): 1-9.]
HU F, ZENG FP, DU H, et al., 2018. Scale-dependent spatial patterns for species diversity in a karst evergreen and deciduous broad-leaved mixed forest of Northwest Guangxi [J]. Acta Ecol Sin, 38(17): 6074-6083. [胡芳, 曾馥平, 杜虎, 等, 2018. 桂西北喀斯特常绿落叶阔叶混交林物种多样性分布格局的尺度效应 [J]. 生态学报, 38(17): 6074-6083.]
HUANG YT, YAO L, AI XR, et al., 2015. Quantitative classification of the subtropical evergreen-deciduous broadleaved mixed forest and the deciduous and evergreen species composition structure across two national nature reserves in the southwest of Hubei, China [J]. Chin J Plant Ecol, 39(10): 990-1002. [黃永涛, 姚兰, 艾训儒, 等, 2015. 鄂西南两个自然保护区亚热带常绿落叶阔叶混交林类型及其常绿和落叶物种组成结构分析[J]. 植物生态学报, 39(10): 990-1002. ]
KANG MM, 2010. Dynamicsof restoration of disturbed evergreen broad-leaved forests and ecological strategies of main woody species in Tiantong national forests park, Zhejiang [D]. Shanghai: East China Normal University. [康敏明, 2010. 浙江天童受损常绿阔叶林恢复动态及主要树种适应策略 [D]. 上海: 华东师范大学. ]
LI MD, 2017. Usingthe Forest-DNDC model to simulate karst forest carbon cycle characteristics [D]. Beijing: University of Chinese Academy of Sciences. [李梦德, 2017. 利用Forest-DNDC模型模拟喀斯特森林碳循环特征 [D]. 北京: 中国科学院大学.]
LI YJ, LI JF, CHANG B, et al., 2019. Scale variation and correlation of plant functional traits in the dominant Cyclobalanopsis glauca community in the karst area of Guilin City, Southwest China [J]. Acta Ecol Sin, 39(15): 5555-5563. [李月娟, 李娇凤, 常斌, 等, 2019. 桂林岩溶石山青冈群落植物功能性状的尺度变化与关联 [J]. 生态学报, 39(15): 5555-5563.]
LI YZ, WANG HX, ZHU JJ, 2009. Monthly changes of leaf area index and canopy openness of Larix olgensis in mountainous regions in east Liaoning Province [J]. J NE For Univ, 37(7):20-22. [李根柱, 王贺新, 朱教君, 2009. 辽东山区长白落叶松叶面积指数和林冠开阔度的月动态 [J]. 东北林业大学学报, 37(7): 20-22.]
LIANG SC, 2015. Ecology [M]. Wuhan: Huazhong University of Science and Technology Press. [梁士楚, 2015. 生态学 [M]. 武汉: 华中科技大学出版社.]
LIANG SC, HU G, ZHANG ZH, et al., 2018. Ecological study of common woody plants in Guilin karst rocky mountains [M]. Beijing: Science Press. [梁士楚, 胡刚, 张忠华, 等, 2008. 桂林岩溶石山常见木本植物生态学研究 [M]. 北京: 科学出版社.]
LIN QK, 2017. Effects of gaps on the plants regeneration and species diversity in Tiantong evergreen broad-leaved forest [D]. Shanghai: East China Normal University. [林庆凯, 2017. 天童常绿阔叶林林窗对植物群落更新和物种多样性的影响 [D].上海: 华东师范大学.]
LIU CC, QIAO XG, WANG Z, et al., 2020. Distinct taxonomic and phylogenetic patterns of plant communities on acid and limestone soils in subtropical and tropical China [J]. J Veg Sci, 31(1): 194-207.
LIU RH, TU HR, LI JF, et al., 2019. Numerical classification and ordination of Cyclobalanopsis glauca communities in karst hills of Guilin, Southwest China [J]. Acta Ecol Sin, 39(22): 8595-8605. [刘润红, 涂洪润, 李娇凤, 等, 2019 . 桂林岩溶石山青冈群落数量分类与排序 [J]. 生态学报, 39(22): 8595-8605]
LIU YL, GUO RQ, SUN SC, 2010. Variations in the vertical vegetation zonation of subtropical Chinese mountains: The importance of climatic seasonality [J]. Acta Ecol Sin, 30(14): 3912-3922. [劉亚兰, 郭汝清, 孙书存, 2010. 中国亚热带山地植被垂直带分布对气候季节性的响应 [J]. 生态学报, 30(14): 3912-3922.]
LONG WX, 2011. The community structure and assembly rules of tropical cloud forests in Hainan Island, South China [D]. Beijing: Chinese Academy of Forestry. [龙文兴, 2011. 海南岛热带云雾林群落结构及组配机制研究 [D]. 北京: 中国林业科学研究院.]
NEE S, 2005. The neutral theory of biodiversity: Do the numbers add up? [J]. Funct Ecol, 19(1): 173-176.
OU ZL, LI XK, SU ZM, 2005. Population variation and species diversity in succession process of karst vegetation in Yangshuo, Guangxi [J]. Ecol Sci, 24(4): 295-297. [欧祖兰, 李先琨, 苏宗明, 2005. 广西阳朔岩溶植被演替过程种群变化及物种多样性 [J]. 生态科学, 24(4): 295-297.]
POORTER L, MARKESTEIJN L, 2008. Seedling traits determine drought tolerance of tropical tree species [J].Biotropica, 40(3): 321-331.
QUIGLEY MF, PLATT WJ, 2003. Composition and structure of seasonally deciduous forests in the Americas [J].Ecol Monogr, 73(1), 87-106.
SI B,2007. Study on restoration models of vegetation and their characteristics in typical karst rocky desertification area [D]. Chongqing: Southwest University. [司彬, 2007. 典型喀斯特石漠化地区植被恢复模式及其特征研究 [D]. 重庆: 西南大学.]
SONG TQ, PENG WX, ZENG FP, et al., 2010. Community composition and biodiversity characteristics of forests in karst cluster-peak-depression region [J]. Biodivers Sci, 18(4): 355-364. [宋同清, 彭晚霞, 曾馥平, 等, 2010. 喀斯特峰丛洼地不同类型森林群落的组成与生物多样性特征 [J]. 生物多样性, 18(4): 355-364.]
SU RGG, ZHANG JT, ZHANG B, et al., 2010. Numerical classification and ordination of forest communities in the Songshan National Nature Reserve [J]. Acta Ecol Sin, 30(10): 2621-2629. [苏日古嘎, 张金屯, 张斌, 等, 2010. 松山自然保护区森林群落的数量分类和排序 [J]. 生态学报, 30(10): 2621-2629.]
SU ZM, LI XK, 2003. The types of natural vegetation in karst region of Guangxi and its classified system [J]. Guihaia, 23(4): 289-293. [苏宗明, 李先琨, 2003. 广西岩溶植被类型及其分类系统 [J]. 广西植物, 23(4): 289-293]
SUN XW, 2018. Vegetation classification based on mature community and gap-disturbance community in Tiantong Mountain, Zhejiang Province [D]. Shanghai: East China Normal University. [孙小伟, 2018. 浙江天童山成熟群落与林窗干扰群落的植被分类研究 [D]. 上海: 华东师范大学.]
YANG XD, 2014. Effects of plants light utilization and water transport capacity on stratification of tree architecture in evergreen broad-leaved forests [D]. Shanghai: East China Normal University. [杨晓东, 2014. 常绿阔叶林植物光线利用和水分运输能力对树木构型垂直层次性的影响 [D]. 上海: 华东师范大学.]
YUAN DX, ZHANG C, 2008. Karst dynamics theory in China and its practice [J]. Acta Geosci Sin, 29(3): 355-365. [袁道先, 章程, 2008. 岩溶动力学的理论探索与实践 [J]. 地球学报, 29(3): 355-365.]
ZHANG JT, 2018. Quantitative ecology [M]. 3rd ed. Beijing: Science Press. [张金屯, 2018. 数量生态学 [M]. 3版. 北京: 科学出版社.]
ZHU JJ, KANG HZ, HU LL, 2005. Estimation on optical porosity or canopy closure for a forest stand with hemispherical images [J]. Chin J Ecol, 24(10): 1234-1240. [朱教君, 康宏樟, 胡理樂, 2005. 应用全天空照片估计林分透光孔隙度(郁闭度) [J]. 生态学杂志, 24(10): 1234-1240.]
(责任编辑 蒋巧媛)
收稿日期: 2020-12-13
基金项目: 国家自然科学基金(31860124); 2020年广西高校大学生创新创业计划项目(202010602054,202010602253) [Supported by the National Natural Science Foundation of China (31860124); College Students Innovation and Entrepreneurship Training Program (202010602054, 202010602253)]。
作者简介: 林红玲(1996-),硕士研究生,主要研究方向为植物生态学,(E-mail)1043282896@qq.com。
通信作者: 姜勇,博士,副教授,研究方向为植物生态学,(E-mail)yongjiang226@126.com。
