广西大石围天坑群天坑森林主要木本植物种间关联性研究
2021-08-16黄林娟于燕妹安小菲余林兰薛跃规
黄林娟 于燕妹 安小菲 余林兰 薛跃规

摘 要: 為了解大石围天坑群天坑森林植物群落的种间联结特征,该文以大石围天坑群中7个典型天坑森林为研究区建立18个样地,取其中重要值大于1的木本植物为研究对象,采用方差比例、 χ2检验、Pearson相关系数检验、Spearman秩相关系数检验及PCA排序方法,对天坑森林内的主要木本植物种间关联性和生态种组进行定量分析。结果表明:(1)乔木层总体联结性呈现显著正联结,灌木层总体联结性呈现不显著正联结,群落总体关联性呈现显著正联结,表明天坑森林植物群落处于稳定的阶段。(2) χ2检验、Pearson相关系数检验和Spearman秩相关系数检验发现,乔木层正联结种对多于负联结种对且显著联结种对占多数,灌木层负联结种对多于正联结种对,且显著联结种对占多数,表明乔木层群落稳定性高于灌木层,乔木层种间竞争合作强,灌木层种间竞争排斥性强,群落主要木本植物之间联结性强,种间保持紧密联结的联结特征。(3)PCA排序将天坑森林植物分为四大生态种组,物种分布由天坑底部至顶部呈现圈层分布方式,且物种分布方式总体受生境差异和物种自身特性影响。可见,植物紧密的联结关系是天坑森林多样性维持机制之一,而群落内部种间关联性的自我调控及物种对生境异质性的趋同适应是群落维持稳定的基础。
关键词: 天坑森林, 种间关联性, 主要木本植物, 生态种组, 圈层分布, 大石围天坑群
中图分类号: Q948.1
文献标识码: A
文章编号: 1000-3142(2021)05-0695-12
Interspecific association of main woody plants in tiankeng
forests of Dashiwei Tiankeng Group, Guangxi
HUANG Linjuan1,2, YU Yanmei1,2, AN Xiaofei1,2, YU Linlan1,2, XUE Yuegui1,2*
( 1. College of Life Sciences, Guangxi Normal University, Guilin 541006, Guangxi, China; 2. Key Laboratory of Ecology of Rare and Endangered
Species and Environmental Protection (Guangxi Normal University), Ministry of Education, Guilin 541006, Guangxi, China )
Abstract: For understanding the interspecific associations of main woody plant in tiankeng forests, we chose seven typical tiankeng forests as researched area and established a total of 18 sample plots in Dashiwei Tiankeng Group. We selected woody plants with importance values >1 as the study object, and quantitatively analyzed interspecific associations and correlations by a set of methods, namely, variance ratio, χ2 test, Pearson correlation coefficient test, Spearman rank correlation coefficient test and principal component analysis ( PCA ). The results were as follows: (1)Overall interspecific association were significant positive correlations in tree layer, non-significant positive associations in shrub layer and significant positive associations in community, indicating that the plant community in tiankeng forests was at a stable stage. (2)The χ2 test, Pearson correlation coefficient test and Spearman rank correlation coefficient test found that there was a difference between positive and negative correlation in tree layer and shrub layer. Many positive correlation species pairs existed in tree layer and many negative correlation species pairs existed in shrub layer, but both of them were more significant correlation species pairs than non-significantanes, which indicates that the stability of community composition in tree layer was higher than shrub layer, and there was a strong competition and cooperation between species pairs in tree layer, a strong competition and repulsion between species pairs in shrub layer, and a close interspecific association in community. (3)According to the results of PCA, the 24 main woody species were divided into four ecological species groups. The tiankeng forests community distribution was in a circle distribution pattern from the bottom to the top of the tiankeng topography, which was affected by environmental differences and species characteristics. In conclusion, these results suggest that the close interspecific association may be one of the maintenance mechanisms of tiankeng forest species diversity, while the self-regulation of interspecific association within the community and convergence adaptation of species to habitat heterogeneity are the basis for maintaining the stability of the community.
Key words: tiankeng forest, interspecific association, main woody plants, ecological species group, circle distribution, Dashiwei Tiankeng Group
种间联结是以物种与物种之间的相互作用为基础,研究生境异质下物种在空间分布上的相互联结性(张金屯等,2003;Muhamed et al., 2014; Yuan et al., 2018)。由于不同的物种对生境因子的需求不同,造成了物种间复杂的关系(王乃江等,2010)。对植物群落种间联结性的研究,有利于了解种群在生境中的结构、功能和地位,预测种群数量的动态变化和群落的演替,对退化喀斯特地区植被的恢复、生物多样性的保护和森林资源的可持续利用具有重要的意义(张金屯,2011;殷声锋等,2016)。因此,很多学者采用定性或定量的方法,结合物种的多度、重要值等数量特征(徐满厚等,2014),对常态地貌上的植物群落进行了大量种间关联性研究,但对一些特殊生境中的植物群落种间关系的研究却较少。例如,高寒草原植物群落研究发现群落总体关联表现为不显著联结,伴生种或杂草在群落中表现出很强的依赖性(房飞等,2012),桂西南喀斯特山地蚬木(Excentrodendron tonkimense)次生林群落总体种间关系表现为显著正联结,枫香(Liquidambar formosana)和麻栎(Quercus acutissima)次生林群落总体种间关系表现为不显著正关联植物群落,群落中生态位重叠指数与种对间的显著正联结种对在群落中所占比例有关(马丰丰等,2017)。
天坑森林,形成于天坑发育中晚期,为天坑负地形中所形成的森林,主要分布在我国西南地区,为特殊生境下的喀斯特森林生态系统(朱学稳等,2003;黄保健, 2004)。天坑特殊地形中土壤、水分和光照等生态因子分布差异显著,物种经过长期的相互作用与适应进化,形成了自己独特的天坑森林植物群落(Su et al., 2017)。作为负地形生境下的原生性森林,其群落结构、群落外貌、区系组成、垂直结构及群落的演替等方面均与一般的喀斯特森林不同,为石灰岩植物区系的现代避难所(冯慧喆,2015;苏宇乔等,2016)。因此,探讨天坑森林植物群落种间关系,在对深入了解喀斯特负地形生境下物种的组合、种群进化和群落物种多样性维持机制方面具有重要的作用(林宇,2005;范蓓蓓,2014)。本文首次对大石围天坑群天坑森林主要木本植物种间联结性研究,主要探究以下几个问题:(1)喀斯特负地形生境下植物群落的分布方式;(2)负地形生境下植物群落种对间的联结特征;(3)以种间联结特征为基础,进一步探讨天坑森林物种多样性维持机制。
1 研究区域概况
广西大石围天坑群(106°10′—106°51′ E,24°30′—25°03′ N)位于中国广西壮族自治区乐业县,属桂西山原北部,由贵州高原向广西盆地过渡的高峰深洼带。该区域地理位置处于暖湿气流交汇处,气候温和、降水丰富、干湿季分明,具有地方特色的中亚热带温湿气候;水文条件优越,处于百朗地下河“S”型褶皱中部,水利资源充沛;地形地貌复杂、多样,主要以岩溶地貌形成的石山为主,包括大大小小、形态各异的天坑溶洞和岩溶峰丛等;海拔差异较大,天坑坑口集中出现在1 200~1 266 m之间;土壤以石灰岩风化的石灰土为主,pH一般呈现中性或偏碱性;植被类型多样、复杂,主要为常绿阔叶林和常绿落叶混叶交林。天坑森林则分布在天坑负地形结构上,以耐阴性和喜阳性常绿阔叶物种居多,落叶物种数较少,且随着天坑坑口的开放而逐渐增多(范蓓蓓,2014;朱学稳等,2018)。
2 研究方法
2.1 样地设置
在充分踏查了众多天坑森林的基础上,本次研究结合天坑森林面积、群落结构和物种丰富度等特点,选择了7个具有代表性的天坑森林为研究对象,建立了18个20 m × 20 m的样地,样方情况如表1所示,并在此基础上将每个样地划分为4个10 m × 10 m的小样方进行灌木调查。对样方内所有胸径≥2 cm的木本植物进行全面调查,测量并记录物种种名、胸径、高度和盖度等,同时记录每个样方的海拔、经纬度、坡度、坡向等生境因子。
2.2 数据处理
2.2.1 群落总体联结性检验 本文分别选取天坑森林内乔木层和灌木层重要值大于1的16种和10种主要物种进行群落总体联结性分析与种间联结性分析。群落的总体联结性首先根据物种在样方中是出现-不出现,将原始数据转换成(0,1)二元数据矩阵;然后使用方差比率法來测量VR值,并通过统计量W是否落入(χ20.95,χ20.05)置信区间来检验总体关联性是否显著。详细方法及计算公式参考Schluter(1984)和王伯荪和彭少麟(1985)。
2.2.2 种间联结性检验 种间联结性采用χ2检验进行种对间的定性分析,基于物种在样方中存在-不存在,将原始数据转换成(0,1)二元数据矩阵进行计算,并进一步使用多度数据进行Pearson相关系数和Spearman秩相关系数检验,详细检验方法及计算过程参考张金屯(2011)。
2.2.3 生态种组 生态种组是一个群落中生态习性相似和生物学形态类似的物种组合(张明等,2017)。本研究综合物种χ2检验、Spearman秩相关系数、Pearson相关系数检验的结果,运用PCA排序,进行生态种组的划分,详细检验方法及计算过程参考Husson et al.(2010)。本文所有数据处理均在WPS Excel 2020和R.4.0.0中处理。
3 结果与分析
3.1 群落总体联结性
大石围天坑群天坑森林植物群落的总体联结(表3)显示,灌木层方差比例值(VR=2.556 5)大于1,且检验值落入效应区间(9.39,28.87)内,表明灌木层总体联结呈现不显著的正联结,乔木层(VR=1.209 2)和群落总体(VR=3.298 9)的方差比例值大于1,且检验值未落入效应区间(9.39,28.87)内,表明乔木层和群落总体种间联结呈现显著的正联结。以上结果说明了天坑森林木本植物群落总体上处于相对稳定的阶段,且乔木层物种稳定性要高于灌木层。
3.2 种间联结性分析
3.2.1 乔木层种间联结性 从χ2检验半矩阵图(图1)可以看出,在乔木层120对主要种对中,正联结的有78对,占总对数的65%;负联结的有42对,占总种对数的 35%;正负联结比为1.86。其中:极显著正联结的有9对,包括短叶黄杉(Pseudotsuga brevifolia)与化香树(Platycarya strobilacea)、香木莲(Manglietia aromatica)与多核鹅掌柴(Schefflera brevipedicellata)、 南酸枣 (Choerospondias axillaris) 与灰岩润楠(Machilus calcicola)、南酸枣与山胡椒(Lindera glauca)、掌葉木(Handeliodendron bodinieri)与簇叶新木姜子(Neolitsea confertifolia)、粉叶润楠(Machilus glaucifolia)与多核鹅掌柴、贵州琼楠(Beilschmiedia kweichowensis )与灰岩润楠、贵州琼楠与近轮叶木姜子(Litsea elongata var. subverticillata)、近轮叶木姜子与山胡椒,占总种对数的7.5%;极显著负联结的有3对,包括化香树与掌叶木、化香树与短叶黄杉、短叶黄杉与灰岩润楠,占总种对数的2.5%;显著联结的有84对,占总种对数的70%;不显著联结的有36对,占总种对数的30%,绝大多数种对呈现显著联结。因此,天坑森林乔木层主要物种之间的种间竞争合作较强,种间联结紧密。
由Pearson相关检验结果(图2:a,表4)得知,在120个种对中,正联结的有73对,占总对数的60.83%;负联结的有47对,占总种对数的 39.17%;正负联结比为1.55。其中,极显著正联结的有10对,包括香木莲与簇叶新木姜子、香木莲与日本杜英(Elaeocarpus japonicus)、南酸枣与川钓樟(Lindera pulcherrima var. hemsleyana)、南酸枣与山胡椒、簇叶新木姜子与日本杜英、 贵州琼楠与灰岩润楠、贵州琼楠与近轮叶木姜子、贵州琼楠与山胡椒、近轮叶木姜子与灰岩润楠、近轮叶木姜子与山胡椒,占总种对数的8.33%;无极显著负联结种对存在;显著联结种对有77对,占总种对数的64.1%;不显著联结种对有43对 ,占总种对数的35.83%。绝大多数种对呈现显著联结,表明乔木层主要物种之间的种间竞争合作较强,种间联结紧密,此结果与乔木层χ2检验的结果一致。
由Spearman秩相关系数检验结果(图2:b,表4)得知,在120个种对中,正联结的有71对,占总对数的59.17%;负联结的有49对,占总种对数的 40.83%;正负联结比为1.45。其中:极显著正联结的有9对,包括香木莲与日本杜英、南酸枣与山胡椒、簇叶新木姜子与日本杜英、簇叶新木姜子与灰岩润楠、贵州琼楠与灰岩润楠、贵州琼楠与近轮叶木姜子、贵州琼楠与山胡椒、近轮叶木姜子与灰岩润楠、近轮叶木姜子与山胡椒,占总种对数的7.5%;无极显著负联结种对存在;显著联结的有85对,占总种对数的71%;不显著联结的有35对,占总种对数的29%。绝大多数种对呈现显著联结,表明乔木层的主要物种之间的种间竞争合作较强,种间联结紧密,此结果与乔木层χ2检验和Pearson相关检验结果一致。
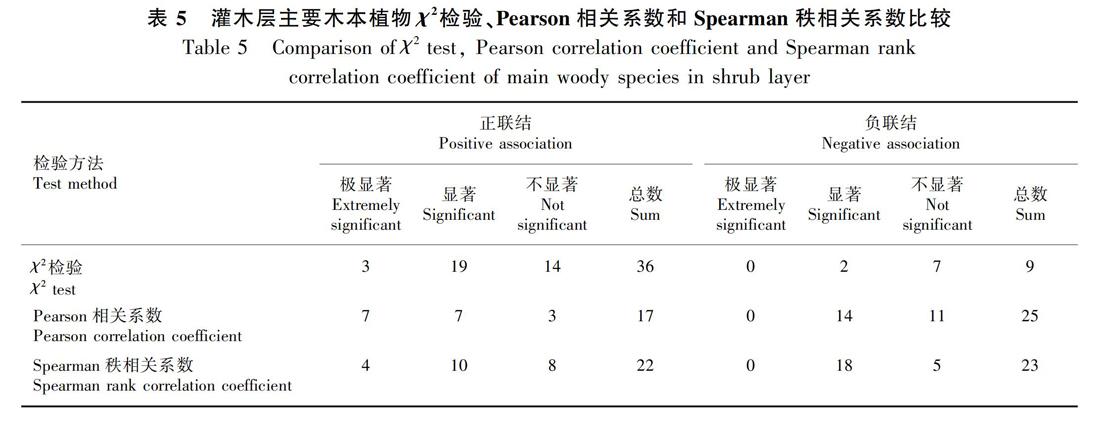
3.2.2 灌木层种间联结性 从χ2检验(图3)可以看出,灌木层45个种对间呈正联结的有36对,占总对数的80%;负联结的有9对,占总对数的20%;正负联结比为4。其中:极显著正联结的有3对,包括通脱木(Tetrapanax papyrifer)与水同木(Ficus fistulosa)、珊瑚树(Viburnum odoratissimum)与红果山胡椒(Lindera erythrocarpa)、珊瑚树与水同木,占总种对数的3%;无极显著负联结种对存在;显著联结的有24对,占总种对数的53.33%;不显著联结的有21对,占总种对数的46.67%。绝大多数种对呈现显著联结,天坑森林灌木层主要物种之间的种间竞争排斥较强,种间联结紧密。
由Pearson相关检验结果(图4:a,表5)得知,在45个种对中,正联结的有17对,占总对数的37.78%;负联结的有25对,占总种对数的 55.56%;正负联结比为0.68。其中:极显著正联结的有7对,包括通脱木与珊瑚树、通脱木与水同木、通脱木与红果山胡椒、乌冈栎(Quercus phillyreoides)与化香树、珊瑚树与水同木、珊瑚树与红果山胡椒、水同木与红果山胡椒,占总种对数的6%;无极显著负联结种对出现;显著联结的有17对,占总种对数的68.89%;不显著联结的有14对,占总种对数的31.11%。绝大多数种对呈现显著关联,表明灌木层的主要物种之间的种间竞争排斥较强,种间联结紧密,此结果与灌木层χ2检验的结果一致。
由Spearman秩相关系数检验结果(图4:b,表5) 得知, 在45个种对中, 正联结的有22对,占总对数的49%;负联结的有23对,占总种对数的51%;正负联结比为0.96。其中:极显著正联结的有5对,包括星毛鸭脚木(Schefflera minutistellata)与通脱木、通脱木与水同木、珊瑚树与水同木、珊瑚树与红果山胡椒、水同木与红果山胡椒,占总种对数的3%;无极显著负联结种对出现;显著联结的有32对,占总种对数的71.11%;不显著联结的有13对,占总种对数的28.89%。绝大多数种对呈现显著关联,表明灌木层的主要物种之间的种间竞争排斥较强,种间联结紧密,此结果与灌木层χ2检验和Pearson相关检验结果一致。
3.3 生态种组划分
本研究结合物种的生态习性、种间联结性及其生境特征,利用PCA排序将天坑森林内主要木本植物分为四大生态种组(图5),结合样地调查结果发现,物种分布由负地形结构底部至顶部边缘区呈现一定的圈层变化,主要木本植物逐渐由第I生态种组,过渡到第Ⅱ和第Ⅲ生态种组,最后为第Ⅳ生态种组。第I生态种组包括香木莲、多核鹅掌柴、星毛鸭脚木、通脱木、珊瑚树、水同木、红果山胡椒、异叶梁王茶(Metapanax davidii),该组植物喜湿喜肥、较耐阴,正联结种对较多,主要分布于土壤聚集效应强、水分含量及土壤有机质含量最高的天坑底部森林,物种生态习性与生境基本一致。第Ⅱ生态种组包括贵州琼楠、日本杜英、针齿铁仔(Myrsine semiserrata)、 灰岩润楠、 簇叶新木姜子,该生态组植物较喜湿耐阴,种对间呈现显著正联结,种间联结性强,主要分布于岩石裸露、坡度较大、土壤层较薄弱的天坑中部倒石坡,接近底部森林,物种生态习性与生境基本一致。第Ⅲ生态种组包括川钓樟、掌叶木、南酸枣、青冈(Cyclobalanopsis glauca)、西桦(Betula alnoides)、粉叶润楠、山胡椒、近轮叶木姜子,该生态组植物略喜阳,种对间既有正联结又有负联结,显著正联结种对较多、种间联结性强,分布于岩石裸露、坡度较大、土壤层薄弱的天坑中部倒石坡森林, 接近边缘森林,物种生态习性与生境基本一致。第Ⅳ生态种组包括短叶黄杉、乌冈栎、化香树,该组植物喜阳耐贫瘠,种间联结呈显著正联结,种间联结性强,主要分布在海拔高、光照充足的天坑顶部边缘森林,生态习性与生境基本一致。同一生态种组的物种,生态习性、资源利用方式及对环境的适应性均具有相似性,通过生态位分离方式,种对间具有较强的正联结关系,不同的生态种组因自身生物学特性、资源的利用及生态习性等方面的差异性,对生境的适应程度不一致而导致生态位重叠,种对间保持一定的负联结。
4 讨论
4.1 天坑森林物种圈层分布特征
生态种组的划分以种间联结性为基础,将群落中生态习性相似的种联合起来,反映种群对环境的适应及物种之间的协同进化(Su et al., 2015; Vahdati et al., 2017)。在大石围天坑群天坑森林分布区, 不同的生态种组物种集群分布于同一生境中。在垂直方向上,天坑环境由负地形底部至顶部,土壤、水分、光照、温度等环境因子呈现一定的梯度分布差异。首先,负地形结构上土壤的堆积效应由天坑底部至顶部逐渐减弱,坑底的土壤有机质含量、土壤肥力均高于其周边地形;其次,天坑底部与地下河流连接,为植物的生长提供充足的水分,且负地形底部有利于水分、热量的留存,而顶部易于流失;最后,受负地形结构影响,坑底受植物的遮挡较多,光照较少,以多核鹅掌柴、水同木等喜湿喜肥较耐阴植物居多,中部为底部和顶部植物的中间过渡带,光照被崖壁遮挡,物种以香木莲、南酸枣等略喜阳耐阴物种为主,而到了顶部,窗口全开,光照最为充足,多分布化香树、短叶黄杉、乌冈栎等喜阳物种(冯慧喆,2015;朱学稳等,2018)。在垂直方向上,负地形结构上群落的圈层分布加大了种间的相遇概率,使物种面对垂直水平生境的差异时,可以通过对生境的适应和生态位的互补,维持多物种共存。在水平方向上,天坑地形地貌特殊,生境多样化、复杂化,小尺度下生境异质性加大了物种之间的组合方式,物种通过种间竞争合作,使不同的物种共存于同一镶嵌体(Tilman, 2000)。这种长期垂直和水平方向上的物种共存状态,形成了天坑森林物种圈层分布的特殊分布方式。
4.2 天坑森林种间联结性分析
在环境条件相似的情况下,物种通过资源的划分、习性互补等生态策略产生生态位的重叠,种间关系表现为一方的偏利或双方互利的正联结(徐晓峰等,2018)。例如,在天坑森林种对中,短叶黄杉与化香树,均处于乔木层第一层,对光照、水分等环境资源的需求相似,两者通过种间合作合理划分资源,种间关系呈现极显著正联结;掌叶木与多核鹅掌柴,掌叶木喜阳而多核鹅掌柴喜湿耐阴,当处于天坑同一微生境时,通过生态习性互补,两者生态位重叠而呈现显著正联结。在环境条件差异较大的情况下,物种因生物学特性的不同、生境差异和资源竞争压力而产生的生态位分离,种间关系则表现为物种一方或双方的负联结(Baron et al., 2015)。例如,在天坑森林种对中,化香树与香木莲,化香树对土壤肥力需求较弱,对光照的需求较强,因此常分布在光照充足、地势较高的天坑森林边缘区,而香木莲,喜阳喜肥,分布在土壤有机质含量高的天坑森林核心区至边缘区中间过渡带,两者因光照、土壤营养元素等生态因子的差异而产生了生态位分离,种间关系呈现显著负联结;种对通脱木与异叶梁王茶,均分布在天坑森林底部林下层,两者因对同一空间剩余资源的分配而产生激烈竞争,种间排斥性强,种间关系呈现显著负联结。种对间由于生态习性的相似性、差异性和互补性,产生生态位重叠和生态位分离,实现种对间联结,此结果与多数群落的种间联结方式一致(刘润红等,2018)。
根据Pearson相关系数和Spearman秩相关系数检验显示,乔木层正联结种对多于负联结种对且显著联结种对多于不显著联结种对,种间的联结性强,这与乔木层的总体种间关联性一致。因为天坑森林是一个以乔木树种为主体的植物群落,在长期的演替中,乔木层物种在群落组成、空间分布等方面均占据优势地位(林宇,2005),生态位重叠程度和分化程度高,种对间竞争合作强,从而乔木层整体稳定性高,群落总体呈现显著的正联结。灌木层负联结种对多于正联结种对且显著联结种对多于不显著联结种对,种间的联结性强,这与灌木层的总体种间关联性不一致。这是由于灌木层物种自身空间分布、生物学特性方面处于劣势,部分物种受乔木层物种调控,种对间存在有利的竞争排斥作用,从而灌木层负联结种对较多,稳定性较低,群落总体呈现不显著正联结。例如,乔木层物种山胡椒与灌木层珊瑚树、水同木与红果山胡椒,种对间均为显著负联结,不仅增大了灌木层的种间排斥性,而且也促进群落间的演替(范蓓蓓,2014),反映了乔木层主要种群对灌木层种间关系的调控。群落内部优势种群对其他种群种间联结性的自我调控,进而维持群落总体紧密联结的联结特征。
4.3 群落演替与种间关系
种群间的总体关联性,反映了群落所处的演替阶段以及群落的稳定性,当群落演替到顶级阶段,众学者对此没有统一定论。黄世能和王伯荪(2000)认为群落演替的总体朝向无关联发展;王永繁和余世孝(2005)认为群落演替在不同的尺度下既可以表现为无关联,也可以表现为正联结;一般而言,随着群落的演替,群落的正联结比例越高,群落结构越稳定(刘秀珍等,2010)。大石围天坑群天坑森林主要种群乔木层总体呈显著联结,灌木层呈不显著联结,总体呈现显著正联结,且显著性联结的种对较多,种间的关联性强,表明大石围天坑群天坑森林主要木本植物植物群落,乔木层物种的组成及稳定性要优于灌木层,群落总体处于相对稳定的群落阶段,物种间相互依存,种间联结紧密。这主要是由于天坑森林处于特殊的负地形结构中,受到人类活动的影响较少,因此植物的生长状况良好,使天坑森林内部群落总体呈现稳定局面,这与Bátori et al.(2014)的研究结果一致。天坑独特的负地形结构为现代植物群落的生长提供良好的水热条件(苏宇乔等,2016)。因此,天坑森林群落种间联结性的研究,对本地植物的保护、保育及退化喀斯特地区的植被恢复具有重要意义。
本文从χ2入手,从一定的置信区间上检验种间的联结性,结果显示种间正联结种对多于负联结种对且显著联结种对多于不显著联结种对,表明在植物群落自然演替过程中,群落的结构和物种的组成已趋近于稳定。种间联结的比例明显高于其他地形下的植物群落(郑振宇和龙翠玲,2014),其中的原因与天坑森林的自身特性及其特殊负地形生境有关。一方面,天坑森林以乔木树种为主体,其自身的特性使其在群落演替过程中占据竞争有利地位,并经过长期的植物之间的相互作用及对生境的适应,在天坑内部形成特殊的圈层分布方式,种间关联较强。另一方面,天坑森林独特的天坑负地形对内部物种起到保护和保育作用,使其内部物种联结紧密,对外与周边的喀斯特森林相连(朱学稳,2018),组成大石围天坑群紧密联结的森林植被分布系统。因此,推断紧密的联结关系可能是大石围天坑群天坑森林物种多样性的维持机制之一,而群落内部种间關联性的自我调控及物种对生境异质性的趋同适应是群落维持稳定的基础。天坑森林与周边的喀斯特森林毗邻,但本文仅从天坑森林取样调查进行种间关系的研究是不够的,未来需要进一步将天坑森林与周边的喀斯特森林结合起来,在生态位理论和植物生理生态方面进行种间联结性的研究(王昱等,2018;杨勇等,2018),进一步揭示特殊生境下植物群落的内在联系,为天坑森林物种多样性的维持机制提供参考意义。
5 结论
(1)天坑森林物种分布由负地形结构底部至顶部边缘区呈现一定的圈层分布特征,且其变化原因与天坑垂直和水平方向生境差异有关。(2)天坑森林乔木层物种种间竞争合作强,灌木层物种竞争排斥性强,但群落内部优势种群对其他种群种间联结性的自我调控,从而使群落总体维持紧密的联结特征。(3)基于天坑森林物种分布方式及其联结特征推断,紧密的联结关系是天坑森林多样性维持机制之一,而群落内部种间关联性的自我调控及物种对生境异质性的趋同适应是群落维持稳定的基础。
参考文献:
BARON E, RICHIRT J, VILLOUTREIX R, et al., 2015. The genetics of intra-interspecific competitive response and effect in a local population of an annual plant species [J]. Funct Ecol, 29(10): 1361-1370.
BTORI, ZN, FARKAS TN, ERDS LS, et al., 2014. A comparison of the vegetation of forested and non-forested solution dolines in Hungary: A preliminary study [J]. Biologia, 69(10): 1339-1348.
BAI YH, BI RC, ZHANG QD, 2012. Interspecific relationship of woody plants in Quercus wutaishanica community in Wulu mountain nature reserve, Shanxi Province of China: A quantitative analysis [J]. Chin J Ecol, 31(8):1942-1948. [白玉宏, 毕润成, 张钦弟, 2012. 山西五鹿山自然保护区辽东栎群落木本植物种间数量关系 [J]. 生态学杂志, 31(8): 1942-1948.]
FANG F, HU YK, ZHANG W, et al., 2012. Numerical analysis of inter-specific relationships in alpine steppe community in Bayanbulak [J]. Acta Ecol Sin, 32(6): 1898-1907. [房飞, 胡玉昆, 张伟, 等, 2012. 高寒草原植物群落种间关系的数量分析 [J]. 生态学报, 32(6): 1898-1907.]
FANG BB, 2014. The study on characteristics and succession of karst tiankeng community in Dashiwei, Guangxi [D]. Guilin: Guangxi Normal University: 3-88. [范蓓蓓, 2014. 广西大石围天坑群天坑植物群落特征及演替研究 [D]. 桂林: 广西师范大学: 3-88.]
FENG HZ, 2015. The study onorigion and evolution of karst tiankeng flora in Dashiwei, Guanxi [D]. Guilin: Guangxi Normal University: 3-100. [冯慧喆, 2015. 广西大石围天坑群植物区系的起源和演化研究 [D]. 桂林: 广西师范大学: 3-100.]
HUANG BJ, CAI WT, XUE YG, et al., 2004. Research on tourism resource characteristics of tiankeng group in Dashiwei, Guangxi [J]. Geogr Geogr Infor Sci, (1): 109-112. [黄保健, 蔡五田, 薛跃规, 等, 2004. 广西大石围天坑群旅游资源研究 [J]. 地理与地理信息科学, (1): 109-112.]
HUANG SN, WANG BS, 2000. Dynamics of association between three species in a secondary tropical montane rain forest at Jianfengling on Hainan island [J]. Chin J Plant Ecol, 24(5):569-574. [黄世能, 王伯荪, 2000. 海南岛尖峰岭次生热带山地雨林树种间的联结动态 [J]. 植物生态学报, 24(5): 569-574.]
HUSSON F, LE S, PAGES J, 2010. Exploratory multivariate analysis by example using R, chapman and hall [M]. 1st ed. Florida USA: The Chemical Rubber Company Press.
JIAN XM, SHUI W, CHEN YP, et al., 2018. Interspecific relationships of dominant species in the grassland community of moderately degraded tiankeng of Yunnan, China [J].Chin J Appl Ecol, 29(2):492-500. [簡小枚, 税伟, 陈毅萍, 等, 2018. 云南中度退化的喀斯特天坑草地植物群落优势种种间关系 [J]. 应用生态学报, 29(2): 492-500.]
LIN Y, 2005. Species diversity of karst tiankeng forest in Dashiwei Tiankengs, Guangxi [D]. Guilin: Guangxi Normal University: 3-45. [林宇, 2005. 广西大石围天坑群天坑森林物种多样性研究 [D]. 桂林: 广西师范大学: 3-45.]
LIU RH, JIANG Y, CHANG B, et al., 2018. Interspecific associations and correlations among the main woody plants in a Pterocarya stenoptera community in a riparian zone of Lijiang river, Guilin, Southwest China [J]. Acta Ecol Sin, 38(19):6881-6893. [刘润红, 姜勇, 常斌, 等, 2018. 漓江河岸带枫杨群落主要木本植物种间联结与相关分析 [J].生态学报, 38(19):6881-6893.]
LIU XF, ZHANG F, ZHANG JT, et al., 2010. Analysis on interspecific relation of dominant species of plant communities during succession on abandoned farmlands in Guancen Mountain, Shanxi [J]. J Shanxi Univ (Nat Sci Ed), 33(1): 142-146. [劉秀珍, 张峰, 张金屯, 等, 2010. 管涔山撂荒地植物群落演替过程中优势种种间关系分析 [J]. 山西大学学报(自然科学版), 33(1): 142-146. ]
MA FF, PAN G, LI XQ, et al., 2017. Interspecific relationship and canonical correspondence analysis within woody plant communities in the karst mountains of southwest Guangxi, southern China [J]. J Beijing For Univ, 39(6):32-44. [马丰丰, 潘高, 李锡泉, 等, 2017. 桂西南喀斯特山地木本植物群落种间关系及CCA排序 [J]. 北京林业大学学报, 39(6): 32-44.]
MUHAMED H, LINGUA E, MAALOUF J, et al., 2014. Shrub-oak seedling spatial associations change in response to the functional composition of neighbouring shrubs in coastal dune forest communities [J]. Ann For Sci, 72(2):231-241.
SCHLUTER D, 1984. A variance test for detecting species associations, with some example applications [J]. Ecology, 65: 998.
SHEN LN, HOU MF, XU WB, et al., 2020. Research on flora of seed plants in Dashiwei Karst Tiankeng Group of Leye, Guangxi [J]. Guihaia, 40(6):751-764. [沈利娜, 侯满福, 许为斌, 等, 2020. 广西乐业大石围天坑群种子植物区系研究 [J]. 广西植物, 40(6): 751-764.]
SU SJ, LIU JF, HE ZS, et al., 2015. Ecological species groups and interspecific association of dominant tree species in Daiyun mountain national nature reserve [J]. J Mt Sci, 12(3): 637-646.
SU YQ, TANG QM, MO FY, et al., 2017. Karst tiankengs as refugia for indigenous tree flora amidst a degraded landscape in southwestern China [J]. Sci Rep, 7: 4249.
SU YQ, XUE YG, FAN BB et al., 2016. Plant community structure and species diversity in Liuxing tiankeng of Guangxi [J]. Acta Bot Boreal-Occident Sin, 36(11):2300-2306. [苏宇乔, 薛跃规, 范蓓蓓, 等, 2016. 广西流星天坑植物群落结构与多样性 [J]. 西北植物学报, 3(11): 2300-2306.]
TILMAN D, 2000. Causes, consequences and ethics of biodiversity [J]. Nature, 405(6783):208- 211.
VAHDATI FB, MEHRVARZ SS, DEY DC, et al., 2017. Environmental factors-ecological species group relationships in the Surash lowland-mountain forests in northern Iran [J]. Nord J Bot, 35(2): 240-250.
WANG BS, PENG SL, 1985. Studies on the measuring techniques of interspecific association of lower subtropical evergreen-broadleaved forest. The exploration and the revision on the measuring formulas of interspecific association [J]. Acta Phytoecol Geobot, 9(4): 274-285. [王伯蓀, 彭少麟, 1985. 南亚热带常绿阔叶林种间联结测定技术研究——Ⅰ. 种间联结测式的探讨与修正 [J]. 植物生态学与地植物学丛刊, 9(4): 274-285.]
WANG BS, YU SX, PENG SL, et al., 1996. Handbook of plant community experiments [M]. Guangzhou: Guangdong Higher Education Press. [王伯荪, 余世孝, 彭少麟, 等, 1996. 植物群落学实验手册 [M]. 广州: 广东高等教育出版社.]
WANG NJ, ZHANG WH, LU YC, et al., 2010. Interspecific assocition among the plants communities in the forest at Ziwulin area in Shaanxi Province [J]. Acta Ecol Sin, 30(1): 67-78. [王乃江, 张文辉, 陆元昌, 等, 2010. 陕西子午岭森林植物群落种间联结性 [J]. 生态学报, 30(1):67-78.]
WANG Y, BI RC, RU WM, 2018. Niche andinterspecific associations for dominant speciesin wetland herb communities in the northern branch of the Zhuozhang river watershed, Shanxi [J]. Acta Agr Sin, (2):276-285. [王昱, 毕润成, 茹文明, 2018. 山西浊漳河北源湿地草本植物群落优势种生态位与种间关系 [J]. 草业科学, (2): 276-285.]
WANG YF, YU SX, 2005. Analysis of overall inter-species correlation in the succession process of forest community in Heishiding Nature Reserve, Guangdong [J]. J Sun Yat-Sen Univ(Nat Sci Ed), (4): 88-91. [王永繁, 余世孝, 2005. 黑石顶森林群落演替过程种间总体关联性分析 [J]. 中山大学学报(自然科学版), (4): 88-91.]
XU MH, LIU M, ZHAI DT, et al., 2016. A review of contents and methods used to analyze various aspects of plant interspecific associations [J]. Acta Ecol Sin, 36(24): 8224-8233. [徐满厚, 刘敏, 翟大彤, 等, 2016. 植物种间联结研究内容与方法评述 [J]. 生态学报, 36(24): 8224-8233.]
XU XF, WANG P, ZHANG SB, et al., 2018. Studies on intra and inter-specific competition of Hamamelis mollis [J]. J Trop Subtrop Bot, 26(3): 272-277. [徐晓峰, 王盼, 张苏炯, 等, 2018. 金缕梅种内及其与常见伴生种之间的竞争关系 [J]. 热带亚热带植物学报, 26(3): 272-277.]
YANG Y, XU X, XU Y, et al., 2020. Adaptation strategies of three dominant plants in the trough-valley karst region of northern Guizhou Province, southwestern China, evidence from associated plant functional traits and ecostoichiometry [J]. Earth Environ, 48(4): 413-423. [杨勇, 许鑫, 徐玥, 等, 2020. 黔北优势植物对槽谷型喀斯特生境的适应策略: 基于功能性状与生态化学计量相关联的证据 [J]. 地球与环境, 48(4): 413-423.]
YIN SW, WANG ZH, ZHANG CH, 2016.Interspecific association among dominant species of bryophytes in karst severe rocky desertification area [J]. Ecol Sci, 35(5): 90-95. [殷声锋, 王智慧, 张朝晖, 2016. 喀斯特重度石漠化地区苔藓植物优势种的种间关联性研究 [J]. 生态科学, 35(5): 90-95.]
YUAN ZY, WEI BW, CHEN Y, et al., 2018. How do similarities in spatial distributions and interspecific associations affect the coexistence of quercus species in the Baotianman national nature neserve, Henan, China [J]. Nat Ecol Evol, 8(5): 2580-2593.
ZHANG JT, 2011. Quantitative ecology [M]. 3rd ed. Beijing: Science Press: 133-167. [張金屯, 2011. 数量生态学 [M]. 3版. 北京: 科学出版社: 133-167.]
ZHANG JT, JIAO R, 2003.Interspecific association between woody plants in Shenweigou of Guandi mountains, Shanxi Province [J]. Bull Bot Res, (4):458-463. [张金屯, 焦蓉, 2003. 关帝山神尾沟森林群落木本植物种间联结性与相关性研究 [J]. 植物研究, (4): 458-463.]
ZHANG M, LIU FD, AN SQ et al., 2017. Photosynthetic capacity and water use efficiency of different ecological species groups in tropical montane rain forest, Hainan Island [J]. Ecol Environ, 26(4): 576-581. [张明, 刘福德, 安树青, 等, 2017. 海南热带山地雨林不同生态种组光合能力与水分利用效率 [J]. 生态环境学报, 26(4): 576-581.]
ZHEN ZY, LONG CL, 2014. Interspecific association of woody plant species at different topography sites in Maolan karst forest [J]. For Res Manag, (4): 78-84. [郑振宇, 龙翠玲, 2014. 茂兰喀斯特森林不同地形部位木本植物种间联结性分析 [J]. 林业资源管理, (4): 78-84.]
ZHU XW, CHEN WH, 2006.Tiankengs in the karst of China [J]. Carsol Sin, (S1): 7-24. [朱学稳, 陈伟海, 2006. 中国的喀斯特天坑 [J]. 中国岩溶, (S1): 7-24.]
ZHU XW, HUANG BJ, ZHU DH, et al., 2003. Dashiwei tiankeng group, Leye, Guangxi discoveries, exploration, definition and research [M]. Nanning: Guangxi Science and Technology Publishing House. [朱学稳, 黄保健, 朱德浩, 等, 2003. 广西乐业大石围天坑群发现 探测 定义与研究 [M]. 南宁: 广西科学技术出版社.]
ZHU XW, ZHANG HY, CHEN WH, et al., 2018. Leye Tiankeng [M]. Nanning: Guangxi Science and Technology Publishing House. [朱学稳, 张远海, 陈伟海, 等, 2018. 乐业天坑 [M]. 南宁: 广西科学技术出版社.]
ZHU XW, ZHU DH, HUANG BJ, et al., 2003. A brief study of karst tiankeng [J]. Carsol Sin, (1): 51-65. [朱学稳, 朱德浩, 黄保健, 等, 2003. 喀斯特天坑略论 [J]. 中国岩溶, (1): 51-65.]
(责任编辑 蒋巧媛)
收稿日期: 2021-01-19
基金项目: 国家自然科学基金(3196020109);珍稀濒危动植物生态与环境保护教育部重点实验室研究基金(ERESEP2019Z04) [Supported by the National Natural Science Foundation of China (3196020109); Key Laboratory Research Foundation of Ecology of Rare and Endangered Species and Environmental Protection (Guangxi Normal University), Ministry of Education (ERESEP2019Z04)]。
作者简介: 黄林娟(1994-),硕士研究生,主要从事植物生态学研究,(E-mail)hglinjuan@163.com。
通信作者: 薛跃规,教授,研究生导师,主要从事植物分类和植物区系研究,(E-mail)xueyuegui@126.com。
