香椿种子萌发初始阶段物质代谢的研究
2021-08-16沈永宝尹中明
宋 明,杨 博,沈永宝,尹中明
(1. 崇明区林业站,上海 崇明 202150; 2. 南京林业大学生物与环境学院, 江苏 南京 210037; 3.上海市林业总站,上海 静安 200072)
作为种子植物所特有的延存器官[1],种子萌发是种子植物生活史的重要阶段。 种子萌发时,呼吸代谢加强,其内部会发生一系列的生理生化变化,为种子发芽和幼苗初期的生长提供必需的营养物质和能量[2]。人们对木本植物种子萌发的生理过程已广泛研究[3],如: 白皮松(Pinusbungdan)[4]种子在萌发过程粗脂肪含量前期变化幅度不大,种子突破种皮后才开始大量利用脂肪,并且脂肪降解生成大量的糖。 而阿月浑子(Pistaciavera)[5]、欧 洲 榛子(Corylusavellana)[6]等种子萌发过程也表现出相似的情况。油松(Pinustabulaeformis)[7]种子在萌发初始阶段胚乳中的蛋白质和粗脂肪最先降解。除油松外,更多的研究是注重种子萌发过程后期,即胚根突破种壳之后的时期,而对种子萌发初始阶段,即干燥种子吸水至胚根突破种壳阶段的详细代谢过程,罕有研究报道。 本文以香椿[Toonasinensis(A.Juss)Roem] 种子为试验材料,通过探索种子萌发初始阶段的物质代谢,进一步了解种子在萌发准备阶段内在复杂的生理生化机制。
1 材料与方法
1.1 种子来源
香椿种子由河南省栾川县林业局提供,其千粒重为13.04 g,含水量为11.59%,发芽率为85%。
1.2 种子培养
用风选法挑选饱满的香椿种子300 g,均匀置于有湿润脱脂棉的发芽板中,用湿润脱脂棉覆盖,置于(25±1)℃培养箱中进行培养,并定期喷洒适量水,使种子充分吸水。以胚根突破种皮1 mm为露白标准,以此为取样的结束时间。每9 h取样品1次,测定种子含水量、呼吸速率以及淀粉、可溶性糖等生理、生化指标;用于生理、生化指标测定的样品装入密封袋,放入超低温冰箱中保存。
1.3 测定方法
1.3.1 含水量的测定 在种子培养过程中,按照种子培养方法,每次随机取出10 g种子(每次2个重复),然后用吸水纸吸干其表面的水分,依据林木种子检验规程[8](GB2772-1999),采用低恒温烘干法立即进行种子含水量的测定。
1.3.2 呼吸速率的测定 在种子培养过程中,按照种子培养方法,每次随机取出5 g种子(每次2个重复),然后用吸水纸吸干其表面的水分,放入棉纱布中,用美国LI-COR 公司生产的Li-6400 光合作用呼吸仪来测定种子的呼吸强度。
1.3.3 内含物的测定 以种胚为材料进行各种生理生化指标的测定。可溶性糖及淀粉含量的测定采用蒽酮比色法[9];可溶性蛋白含量的测定采用考马斯亮蓝G-250 染色法[9];粗脂肪含量的测定采用索氏提取法[9]。各指标测定均设3次重复,可溶性糖含量,计算其平均值。
本文数据的分析与处理通过Microsoft Excel软件完成。
2 结果与分析
2.1 种子含水量和呼吸速率的变化
由图1可以看出,香椿种子在前9 h内,种子吸收的水分较多,处于快速吸水阶段。9—27 h香椿种子的吸水速度较9 h前放慢,27 h以后香椿种子含水量逐步趋于稳定状态。

图1 香椿种子萌发初始阶段含水量及呼吸速率的变化
在快速吸水阶段(前9 h),种子迅速吸水,含水量达到51.94%,为干种子含水量的4.48倍。9—27 h期间,含水量由51.94%上升到59.86%,种子吸水幅度变小。培养最初9 h内含水量的大幅度升高主要是靠胶体物质的吸胀作用。种子细胞在其原生质体和细胞壁中含有蛋白质、戊聚糖、淀粉和纤维素等,有很强的亲水性;在培养9 h后,原生质的水合程度趋向饱和,液泡以及大量新的原生质又未形成,因而种子缺少吸水的动力,吸水速度放慢;在种子培养后期,种子胚部细胞开始分裂,胚迅速伸长,细胞体积加大,种子内贮藏的有机物大规模地水解为渗透性很强的小分子物质,这时的吸水是与代谢作用相关的渗透性吸水。
呼吸是种子生命活动的基本过程,它为种子萌发提供必需的能量和物质。一般情况下,种子萌发过程中呼吸代谢是不断加强的。由图1可知,香椿种子也遵循这一规律,其呼吸速率的变化主要可分为3个阶段:在0—27 h这段时期,香椿种子的呼吸速率迅速增加,由开始的0.549 μmol/(g·min)上升到10.645 μmol/(g·min);随后在27—45 h内,香椿种子呼吸速率呈现平缓上升趋势;45 h以后呼吸速率又再次迅速升高。
香椿干种子的气体交换处于极低水平,其呼吸速率仅为0.549 μmol/(g·min),但随着种子含水量的增加,酶系统不断修复、活化,原本“静止状态”的种子呼吸速率明显加强。培养最初的27 h时间内,呼吸速率上升较快,这可能是干种子吸水膨胀后,与线粒体内柠檬酸循环和电子传递链有关的酶系统活化有关;在种子置床培养27—45 h期间,呼吸速率变化平缓,但在45 h之后,呼吸速率又大幅度升高,并于培养81 h达到最高值,表明胚萌发前期细胞不断分裂增长,新线粒体的合成不断增强,呼吸酶类的活化,致使呼吸作用剧增,呼吸速率也随之迅速升高。从图1还可以看出,香椿种子在萌发初始阶段的含水量与呼吸速率的变化基本一致。
2.2 种胚可溶性糖含量的变化
可溶性糖具有合成纤维素、组成细胞壁、转化并组成其他有机物等作用。可溶性糖的分解产物是其他很多有机物合成的原料。可溶性糖类作为呼吸基质,为各种合成过程和各种生命活动提供了所需的能量。
由图2可知,随着香椿种子培养时间的延长,种胚中可溶性糖含量呈现总体下降的趋势。在培养最初9 h内,可溶性糖含量大幅下降,从1.95%降至1.22%,其原因除了可溶性糖因细胞膜功能尚未恢复而外渗外,更多的是作为呼吸代谢的底物被利用。在9—36 h期间,可溶性糖含量下降较缓慢,从1.22%降至0.98%,一方面原因是细胞膜已逐渐修复,内含物质外渗量减少;另一方面原因是细胞内代谢趋于稳定,糖代谢趋于平衡,可溶性糖降解较少。在36—45 h阶段,可溶性糖含量又出现大幅下降,从0.98%降至0.51%,主要是由于该时期胚的代谢活动增强,需要消耗大量的糖类物质。在45 h以后,可溶性糖含量开始回升,这可能是由于淀粉、脂肪等高分子物质大量转化为小分子糖,进而为种子萌发提供能量。香椿种胚的可溶性糖在萌发初期的这种变化规律表明:随着培养时间的延长,种子内部物质代谢逐渐旺盛,可溶性糖作为能量来源被降解或直接作为原料物质被胚利用。

图2 香椿种子萌发初始阶段种胚可溶性糖含量的变化
2.3 种胚淀粉含量的变化
淀粉作为种子内大分子营养物质之一,在种子发育过程中是通过种子本身的生理生化作用,利用可溶性糖转化而成的,并作为贮存物质在种子中累积起来。种子萌发时,淀粉在淀粉酶的作用下降解为小分子的可溶性物质,同时释放出大量的能量供胚吸收利用,其含量的变化是种子代谢状况的重要指标。
由图3可知,随着香椿种子培养时间的延长,种胚中淀粉含量总体上呈现逐步减少的趋势。在前27 h内,淀粉含量呈下降趋势,从48.96%降至41.98%,尤其是9 h内下降幅度较大,从48.96%降至44.88%。培养27—45 h期间,淀粉含量又有所回升,可能是其他贮藏物质降解后转化为淀粉的原因。在种子培养45 h后,淀粉含量再次大幅度下降,并于胚萌发时,含量降至最低28.01%,比干种子淀粉含量低20.95%。萌发初期的这种变化说明随着置床时间的延长,种子中的代谢活动逐渐加强,生理生化反应加剧,种子对碳水化合物的需求也不断增加,因此与此相对应,淀粉含量就不断下降。
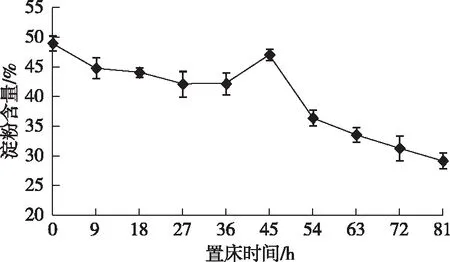
图3 香椿种子萌发初始阶段种胚淀粉含量的变化
2.4 种胚可溶性蛋白质含量的变化
蛋白质是构成生物大分子的一类重要物质,又是酶的基本组成成分。种子萌发时,种子中的贮藏蛋白质在各种酶的作用下降解为小分子氨基酸,供给新蛋白质和其他代谢物质合成之用。因此,蛋白质代谢是种子生命活动的基础,其含量变化可以间接反映种子代谢的强弱。
由图4可知,可溶性蛋白含量随着置床时间的延长,降解生成小肽或氨基酸,含量总体降低。培养0—18 h期间,可溶性蛋白质含量迅速下降,从0.74%降至0.45%,这是由于种子吸水后,贮藏蛋白大量外渗所致。培养18—63 h期间,可溶性蛋白质含量呈现不断下降趋势,从0.45%降至0.32%,主要是由于此段时间种子内的代谢较强,不断生长的胚轴对可溶性蛋白质需求增加,促使贮藏蛋白不断水解的缘故。随后至胚根突破种皮,可溶性蛋白质含量又有所回升,主要是由于在这一时期子叶内粗脂肪等物质降解转化为蛋白质较多的缘故,且贮藏蛋白质之前分解成的氨基酸又重新构成蛋白质。

图4 香椿种子萌发初始阶段种胚可溶性蛋白质含量的变化
2.5 种胚粗脂肪含量的变化
粗脂肪是种子的重要贮藏物质,它在种子内的转化直接影响到种子萌发所需的能量供应和中间产物的来源。种子萌发过程中,脂肪在脂肪酶的作用下水解成脂肪酸和甘油,再进一步转化为糖类,供胚利用。种胚内的脂肪,是种子的重要能源,对种子的萌发有重要作用。
由图5可知,香椿种胚内的脂肪不断降解,其含量总体上呈现下降的趋势。在9 h之前,种胚内物质代谢尚未启动,粗脂肪含量的降低主要是外渗所致,从27.10%降至25.01%。在9—27 h期间,粗脂肪含量缓慢下降,从25.01%降至24.52%,该阶段种子吸水膨胀,酶系统均已活化,但代谢活动很弱,粗脂肪含量变化不大。在27—54 h期间,香椿种子的生理活动已逐渐趋向旺盛,种胚需要更多的养料和能量来满足生长的需要,脂肪既可以通过乙醛酸循环转化为糖,作为呼吸基质被消耗掉,又可作为原料物质供胚利用,所以导致粗脂肪含量的继续下降,从24.52%降至20.05%。而在54 h时出现短暂的回升,可能是种子代谢过程中的一些中间产物又生成脂肪的缘故,随后粗脂肪含量继续下降。

图5 香椿种子萌发初始阶段种胚粗脂肪含量的变化
3 结论与讨论
3.1 种子含水量和呼吸速率变化
种子中的水分是种子生理代谢作用的介质和控制因子。在种子发育、成熟、发芽以及收获以后的不同时期,种子的物理性质和生化变化都与水分的状态及含量有密切的关系。传统的理论把种子吸水分为3个阶段[2]:急剧吸水阶段、滞缓吸水阶段和重新迅速吸水阶段。
香椿种子萌发初期前9 h内,种子吸收的水分较多,速度快,以后随着时间的增加,吸水速度放慢,并逐渐趋于稳定,可见香椿种子吸水过程与传统理论不是完全一致的,只有前2个阶段,或者说第2阶段并不明显。此外,传统理论认为种子吸水的第1阶段是由种子内亲水胶体的吸胀作用引起的,是一个纯粹的物理过程,没有化学变化。但本研究发现,尽管可溶性物质在吸胀阶段并未有明显的变化,但呼吸强度的快速增强可以说明化学变化的存在。由此可知,即使是在第1阶段的水分吸胀期,种子已经开始活化,已经为准备萌发发生各种生化反应。
此外,香椿种子在前9 h内迅速吸水,从11.59%到51.94%,吸水率为348.14%,说明香椿种子具有快速吸水的生物学特性。农作物中吸水率最高的是豌豆,可以达到186%,香椿种子吸水率之高,也是香椿种子容易萌发的主要原因之一,也可能与香椿喜温暖湿润气候有关。各类种子的吸胀速度和吸胀力有明显差异,这与种子的结构和成分有关。
完熟的植物种子组织处于高度脱水状态,因此种子在萌发的初级阶段便是吸水阶段,并伴有明显的呼吸作用。呼吸作用包含着种子内在贮藏物质如淀粉、脂肪、蛋白质等有机物质的氧化分解,同时产生大量ATP能量,用于分解和合成所需酶类的生理活动。因此呼吸作用越强,物质的分解和运转速度越快,生长也就越旺盛。
由香椿种子呼吸速率的变化趋势,可以看出其符合呼吸作用一般理论的3段式规律,即:呼吸速率急剧升高、呼吸作用滞缓期、再次迅速升高。这3个阶段的时间长度因不同物种种子的吸胀速率、种皮对氧气的通透性和代谢速率等因素而异;而且,也因外界条件特别是温度不同而发生显著的变化。香椿种子呼吸速率的变化呈现“S”形曲线,而且出现了从27—45 h的呼吸滞缓期。在少数种子如野燕麦(Avenafatua)、蓖麻 (Ricinuscommunis)[10]等中,氧气的吸收没有明显的滞缓期(第2阶段),其原因尚不清楚,但可能是由于种子伴随着吸胀,较早地建立了有效的呼吸系统,从而确保O2持续利用的增加;而且,在萌发完成之前,种皮的不可透性可能不限制O2的吸收。由此也可以看出,香椿种子含水量的变化与呼吸强度的变化步调并不完全一致。
3.2 种胚贮藏物质的变化
胚乳或子叶中贮藏有丰富的营养物质,主要为糖类、蛋白质和脂肪。不同植物种子中,这3类有机物的含量有很大差异。据此,将种子区分为淀粉种子(淀粉含量多)、油料种子(脂肪含量多)和豆类种子(蛋白质含量多)。这些贮藏物质在种子萌发时分解为简单的有机物,以可利用的形式运输到胚中,作为幼胚生长的营养物质。
香椿种子属小粒种子,胚相对较小,贮藏营养物质的器官主要为子叶。香椿干种子胚中各贮藏物质成分如下:可溶性糖1.95%,淀粉48.96%,可溶性蛋白0.74%,粗脂肪27.1%。可见,香椿种子主要营养物质为淀粉、粗脂肪和可溶性糖,属于淀粉种子。
随着培养时间的延长,香椿种子中可溶性糖、淀粉、可溶性蛋白、粗脂肪的含量均呈现出总体下降的趋势。在种子培养初期,由于细胞膜功能尚未修复,种胚内含物都有不同程度的外渗现象,这在种子中是普遍现象。这些营养物质在培养最初9 h均有不同程度的外渗。随后伴随着种子含水量和呼吸速率的升高,胚中的物质和能量代谢逐渐活跃,贮藏的营养物质迅速被利用,为胚轴的伸长生长提供了充足的物质和能量。
香椿种胚的贮藏物质含量变化的总趋势基本相同,但是它们的降解进程却呈现出一定的差异性。其中可溶性蛋白质动员和粗脂肪最先降解较快,它们在置床27 h后,含量即迅速下降;而淀粉和可溶性糖于培养 36 h后代谢趋向旺盛。可溶性蛋白质和粗脂肪动员较快,是香椿种子萌发初始阶段首先利用的贮藏物质。但在北美黄杉种子萌发过程中,脂类物质最先动员,其次为淀粉和可溶性糖,蛋白质降解最迟[10]。
在种子萌发初始阶段,胚中贮藏的营养物质彼此之间会发生相互转化。如香椿种子在培养45—81 h期间,可溶性糖含量的升高,就可能是淀粉、可溶性蛋白质和粗脂肪转化为糖类物质所致。蛋白质可以通过脱氨基作用转化为糖类[11];粗脂肪则通过乙醛酸循环转变为可溶性糖或者其水解产物甘油逆转为己糖。粗脂肪在降解过程中不但可以转化为糖类,在一定情况下还可转化为蛋白质;香椿种子在培养63—81 h期间可溶性蛋白质含量的增加,可能与粗脂肪的降解转化有关。
