雌雄间性柳树自交子代当年生幼苗表型观测
2021-08-16王保松
周 洁,王保松
(江苏省林业科学研究院,江苏 南京 211153)
柳树为杨柳科柳属的总称,全世界520多种,我国有257种,122变种,33种变型[1]。柳树分布广泛,种类繁多,生长速度快,耐水湿,耐低度盐碱,造林范围广,是集观赏价值、生态价值和经济价值于一体的多功能树种[2]。柳树一般是雌雄异体植物,虽有报道发现柳树存在雌雄同序和两性花[3],但是雌雄同序三性型柳树的自交尚未见报道。
植物表型性状是物种资源研究最基本的方法和途径,是资源保护与评价的关键指标,也是育种的重要基础。关于园林植物观赏价值评价方法的研究比较多,大多数学者从美学角度着手,采用百分制法、层次分析法、心理物理学法、感官评分法、加权评分法、灰色关联分析法和模糊相似优先比法等评价方法,但由于不同植物的观赏重点不同,以及评价过程中各种定性与定量指标权重和分值的不确定性,导致暂且未形成公认的比较成熟的评价方法和评价体系[4-9]。
自交是创制优异中间材料和种质资源的有效方式之一。通过自交可使一些隐性基因得到表现,从而获得丰富的基因变异及表型差异[10]。本研究通过自交的方法建立了柳树的自交F1代,在自交群体中发现了性状的分离,出现枝条的变异、叶片的皱缩、植株的矮化、分枝的变化等表型的改变。通过对表型,如株高、叶片长、叶片宽、枝条弯曲程度及比例的描述和评价,分析柳树自交群体的性状分离程度,为柳树重要性状的分离和基因的鉴定奠定重要的基础。
1 材料与方法
1.1 试验材料
试验材料为雌雄间性柳树品种‘苏柳795’(如图1)自交所获得的子代群体。于2019年1月中采集‘苏柳795’花枝进行水培,置于江苏省林业科学研究院柳树种质资源圃温室,对花枝进行修剪,去除多余的叶片和花芽,3—4月待开花后进行人工杂交。杂交时为避免花粉互相传播,利用纱网将花枝包住,待花序完全开放后,采用人工涂抹的方式进行杂交。将全雄花序和雌雄同序的雄花花粉给全雌花序和雌雄同序的柱头授粉,获得自交种子,收获种子后播于装有泥炭的穴盘中,获得自交苗。待5月苗生长健壮时移栽入装有泥炭珍珠岩混合基质的无纺布袋子中,对所有个体进行编号,以待观测。

a.‘苏柳795’雄花序;b. ‘苏柳795’雌雄同序;c. ‘苏柳795’雌花序。图1 ‘苏柳795’不同性别花序
1.2 表型性状的观测与统计
表型性状的观测于杂交当年秋季10月待苗木生长健壮时统计。叶长、叶宽、叶柄长的取样部位为从顶部开始往下数第3片完整老熟具有代表性的叶片。株高的测量为从植株顶部至基质表面的高度。于当年秋季和次年的春季分别进行测量。
1.3 数据统计分析
使用Excel 2010,SPSS 20.0软件,分别对各种表型性状的数据进行遗传变异度分析。数量性状直方图使用R(V4.0.2版本)软件中自带的hist()函数进行作图。具体参数说明如下:x,输入数据;probability=T绘制样本频次;若为F则绘制密度;col设置直方图中柱子的颜色,xlab设置横轴标签;main设置图标签,xfit,设置横轴显示范围(最小值到最大值),yfit 根据x变量生成拟合y值,lines()函数根据x和y画线,lwd设置线宽。并用R中自带的ks.test()函数和shapiro.test()函数对数据进行处理。
2 结果与分析
2.1 表型性状特征
在自交子代群体中,表型性状枝条、叶片、干型、分枝等都出现了较大的变异。枝条颜色有绿色(b,d)、金色(c)和红色(a,e)(如图2)。枝条姿态有直伸(j)、弯曲(k,n)、扭曲(b,c)、矮化(g)(株高<5 cm)等不同的类型。分枝数和分枝角度也出现了较大的分离,h—n较为高大,有通直没有分枝的类型j,也有分枝数较多的如i和m;分枝部位不同,有基部分枝f,上部分枝的h和中部分枝的i;分枝角度也出现了较大变化,如i的分枝几近平展,l和m的分枝角度较窄。
在自交F1代中,叶片的叶形出现了多种类型(见图3)。‘苏柳795’为披针形,叶柄绿色,无托叶。子代中叶形有披针形(231,292),卵圆形(310,350,140),长披针形(242,11,299),变小(96),叶柄颜色有红绿(242,11)和绿色(66),矮化的叶片没有叶柄,部分植株出现了托叶(271)。叶片正面颜色有深绿(242)、浅绿(48)、黄绿(297)。叶片基部形状‘苏柳795’为契形,140为圆形,299为窄契形。
2.2 自交子代表型性状统计
自交共获得存活的子代478个,生长正常。在植株的表型中,出现了枝条、叶片等的性状分离。干型有通直、弯曲、扭曲3种类型,经统计其中通直的占10.46%,扭曲的为18.41%,弯曲的为71.13%。叶片宽圆形的占13.81%,披针形的占31.17%,线形的为12.13%,叶片缩小的比例为25.73%,且出现了皱缩的现象。有托叶的占2.3%,矮化的占11.72%(见表1)。

图2 自交F1代株型变异

注:叶片编号对应于子代个体编号图3 自交子代叶片性状特征

表1 自交子代各性状比例
2.3 数量性状变异分析
对自交子代478个个体数量性状株高、叶长、叶宽、托叶等进行统计分析。当年杂交苗秋季最高的株高为120 cm,最矮的为1 cm,平均株高为31.89 cm。杂交后第2年春季对株高再次测量,最高株高为205 cm,最矮的为2 cm,平均株高为52.76 cm。叶最长为15.1 cm,最短为0.2 cm,叶宽最长为1.7 cm,最小为0.1 cm,叶片长宽比最大比为15,最小为2;托叶最长为1.7 cm,最小为0.2 cm,托叶宽为0.1—0.2 cm,托叶长宽比为0—12。其中变异最大的为株高,当年株高变异系数为66%,第2年株高变异系数达77%(见表2)。

表2 柳树自交子代数量性状变异分析
2.4 自交子代数量性状频率分布
在对478个自交子代的3个数量性状变异情况统计分析的基础上,绘制了数量性状的分布频次图(见图4)。图4显示叶长、叶宽、株高、叶片长宽比均未呈现正态分布,均呈偏态分布。
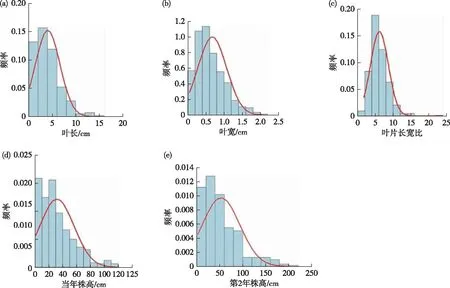
注:a—e分别为叶长、叶宽、叶片长宽比、当年株高、第2年株高分布曲线图4 数量性状分布图
3 结论与讨论
植物表型性状的鉴定与描述是研究种质资源的最简单、易行的基本方法,也是检测遗传变异最传统、最直观的方法[11-12]。柳树在种质资源评价方面已进行了耐盐[13]、耐热[14]、观赏[15]等方面的评价。柳树作为雌雄异株植物,利用自交子代进行数量性状和质量性状的变异分析和评价还是首次。种质资源的遗传变异系数越大,其遗传多样性程度越高。自交子代的性状分离是研究等位基因的重要材料,具有较丰富的遗传变异,变异系数达36%—66%,最高的为株高(达66%),说明自交子代在株高、叶长、叶宽等方面存在丰富的遗传多样性,是种质资源多样性的重要遗传基础,对柳树育种生长迅速、干型好的育种选择具有重要的指导价值。
生物的连续性或间断性变量大多遵从正态分布[16]。本研究发现,柳树自交群体中株高、叶长、叶宽未较好地遵从正态分布,可见在柳树的自交试验中,偏态分布可能也是柳树表型性状中重要的分布形式之一。偏态分布的性状与环境和遗传因素有关。本研究中所有子代为同一批杂交子代,栽培环境一致,来源一致,所以偏态分布和环境因素没有一定的关系。一方面主要和遗传相关,自交使基因发生了纯合,导致了植株的矮化,出现了矮化株高<5 cm的植株,当年杂交苗株高<10 cm的频次大大增加,和平均株高相同;另一方面可能是由于人为选择方向所致[17],即在乔木柳选育工作中,生长快、植株高一直是选择的方向和目标,因为这种选择压力,使得柳树自交群体向此特定方向进化,相应性状的个体增加,形成拖尾现象。
乔木柳的自交群体中出现了株高、叶片、托叶、枝条、皮色等重要性状分离,扭曲的枝条占18.41%,矮化的占11.72%,有托叶的为2.30%。龙爪柳[Salixmatsudanavar.matsudanaf.tortuosa(Vilm.) Rehd.]为旱柳的变种,但是对于枝条扭曲的成因尚不清楚,在自交群体中,扭曲的占近20%,为枝条扭曲的柳树提供了另外一种解释。植株的矮化一般由单一基因控制,水稻中OsDWL1[18]、高粱矮化基因Dw3[19]等多种作物中的矮化基因被鉴定,同时矮化突变体降低了茎秆的同化作用,能将更多的光合产物转移到穗部,显著增加产量。柳树自交群体中的矮化植株占11.72%,为矮化机理的研究和矮化基因的鉴定提供了理想的研究材料。皮色、叶片大小、托叶均为数量性状,为多基因联合调控。柳树SxMAX基因能显著增加分枝数[20],而在自交群体中基部分枝的占8%,可以通过克隆自交材料的SxMAX基因,找到功能区和突变位点,为基因编辑技术提供基础。枝条颜色是林木冬季重要的观赏性状。目前在西葫芦皮色、南瓜果皮等果蔬中已鉴定到控制皮色的基因[21-22],但林木枝条颜色的基因还未见报道。在自交子代中,出现了皮色金色、红色、紫色、绿色等的分离,可为枝条颜色的变异规律和基因克隆提供重要的基础。
柳树观赏性状的评价在于枝条的颜色、枝条的形态,直立和扭曲枝条的性状分离是观赏柳树新的研究性状特性,所以矮化、皮色等性状的鉴定和分枝数、托叶、叶片长、叶片宽、株高等数量性状的鉴定有利于为柳树重要基因的鉴定提供理论基础,自交材料的建立和性状的评价为今后培育柳树高生物量、枝条颜色艳丽、形态各异的柳树提供重要的物质基础。
