巴氏新小绥螨对水稻干尖线虫控制能力的评估
2021-08-11杨思华徐春玲
杨思华,李 曼,陈 淳,徐春玲,谢 辉
(华南农业大学植物保护学院植物线虫研究室/华南农业大学广东省微生物信号与作物病害防控重点实验室,广州 510642)
水稻干尖线虫Aphelenchoidesbesseyi是世界农业上重要的病原线虫之一[1],其为害水稻造成每年约10%~71%的产量损失[2]。水稻干尖线虫是迁移性寄生线虫[3],寄主范围广泛,可以寄生35属200多种植物,其中水稻Oryzasativa和草莓Fragariaananasa是其最主要寄主[4-6]。
水稻干尖线虫病是种传病害,带病种子是此病的主要初侵染源[7]。由于耕作方式和气候的变化,我国水稻干尖线虫病的流行及危害程度加重,研究和实施有效的防治措施显得非常重要。目前实施检疫、采用合理栽培措施、对稻种进行浸种处理和田间施用化学农药是防治水稻干尖线虫病的常用方法,其中最有效的防治方法是水稻种子播种前采用温水或药剂浸种[8-10]。然而,由于种子热处理易影响种子的发芽率,而农药使用所造成的环境问题越来越受到人们的关注,其使用已经受到严格的限制。因此,近年来,植物线虫病害生物防治方法的研究和应用倍受关注。目前植物线虫生物防治的研究主要集中在利用天敌真菌和细菌控制线虫[11-15]。在自然界一些捕食性螨也是植物线虫的潜在生物防治天敌[16,17],已报道的对植物线虫具有防控作用的捕食螨主要有Gaeolaelapsaculeifer[18]、Sancassaniaultima[19]、Tyrophagusputrescentiae[20,21]、Lasioseiusathiasae[22]、Protogamasellusmica[23]、Pergalumnasp.[24]、Blattisociusdolichus[25,26]和Stratiolaelaps scimitus[27]。
巴氏新小绥螨Neoseiulusbarkeri隶属于寄螨目 Parasitiformes、植绥螨科 Phytoseiidae、新小绥螨属Neoseiulus,主要分布在温带地区,可栖息在土壤、仓储食物和多种植物上[29]。该螨能捕食多种害螨和昆虫,可用来防治多种农业害虫[13,30],如西花蓟马Frankliniellaoccidentalis[30]、柑橘全爪螨Panonychuscitri[31]、朱砂叶螨Tetranychuscinnabarinus[32]等。其中,巴氏新小绥螨在防控柑橘全爪螨方面能取得较为显著的效果,已形成较为成熟的商品化运作[33]。周万琴等[34]报道了巴氏新小绥螨可以捕食多种线虫,包括南方根结线虫Meloidogyneincognita二龄幼虫、水稻干尖线虫、腐烂茎线虫Ditylenchusdestructor、小杆线虫Rhabditissp.和香蕉穿孔线虫Radopholussimilis。尚素琴等[35]报道巴氏新小绥螨具有防治腐烂茎线虫的潜力。Yang等[36]报道巴氏新小绥螨能作为南方根结线虫和香蕉穿孔线虫的生物防治天敌。但有关巴氏新小绥螨以水稻干尖线虫为食的发育和繁殖特性和对水稻干尖线虫的捕食功能和控制能力尚未有研究报道。本研究通过测定巴氏新小绥螨以水稻干尖线虫为食的生命表以及捕食密度、饥饿时间和温度对巴氏新小绥螨捕食线虫的的影响,研究巴氏新小绥螨对水稻干尖线虫的捕食作用及其是否是水稻干尖线虫的潜在天敌。
1 材料与方法
1.1 材料
本研究供试的水稻干尖线虫来自华南农业大学植物线虫研究室,腐食酪螨由中国农业科学院植物保护研究所提供,巴氏新小绥螨采自华南农业大学校园内植物的根际土壤。其中巴氏新小绥螨采用腐食酪螨饲养,腐食酪螨采用酵母片粉和麦麸饲养。酵母片粉购于广州市天河区五山路药房,用榨汁机粉碎成酵母粉后使用。
1.2 个体饲育器的制作
根据周万琴等[34]方法并加以改进制作螨的个体饲育器,具体如下:将2 mL离心管的盖子剪下,在离心管盖中注入2%的琼脂至离心管盖高度的1/2处,待琼脂冷凝后,将装有琼脂的离心管盖用黑色胶布固定在直径为35 mm的培养皿上。用厚度约为1 mm的黑色硬纸剪成培养皿底一般大小的圆纸片,放置在该35 mm的培养皿内,使其与培养皿的底部服帖,再倒入5 mL清水将纸片浸泡约1~2 h,倒去多余的水。将植物寄生线虫及巴氏新小绥螨移入到琼脂表面后往培养皿中加水,使水面高度和离心管盖的高度差不多持平,形成水栅防止巴氏新小绥螨跑出,作为巴氏新小绥螨完成生活史和测定捕食功能的饲养小室。
1.3 巴氏新小绥螨以水稻干尖线虫为食的发育繁殖和生命表
用笔尖沾水的毛笔挑出巴氏新小绥螨雌成螨 100~150头,放入饲养小室中培养。将巴氏新小绥螨雌成螨在12 h内产的卵取出进行单个孵化培养。每隔12 h观察并记录其生长发育情况,在卵及幼螨期间不供食。当卵孵化发育成若螨之后,移入到放有足量水稻干尖线虫和腐食酪螨的饲养小室中。每1 d更换一次条件完全相同的饲养小室,出现新的蜕皮壳即可确定螨态已进入下一阶段。在若螨发育至成螨后将雌、雄螨配对,每个饲养小室内各放1头雌成螨和1头雄成螨。试验阶段若有雄螨逃逸或死亡则进行补充。记录雌螨产卵前期、产卵期、产卵后期、日产卵量及寿命。
根据上述获得的数据,计算生命表参数,制作巴氏新小绥螨以水稻干尖线虫和以腐食酪螨为食的生命表,计算生命表参数。
生命表参数的计算公式[37]:净增殖率R0=Σlxmx;平均世代周期T=Σxlxmx/R0;内禀增长率rm=lnR0/T;精准计算 Σe−rmlxmx=1;周限增长率λ=erm;种群倍增时间Dt=ln2/rm。其中,e为自然对数,X表示以天(d)为单位的时间间隔,lx表示雌成螨在X期间的存活率,mx表示在X期间平均每头雌成螨产下的雌性后代数。
1.4 巴氏新小绥螨对线虫的捕食功能
巴氏新小绥螨捕食线虫功能的测定参考Yang等[36]的方法并加以改进。在每个饲养小室中分别放入水稻干尖线虫,水稻干尖线虫密度设为100、200、300、400、500条。在每个饲养小室分别放入1头经1 d饥饿处理的巴氏新小绥螨雌成螨。在黑暗培养1 d后观察记录水稻干尖线虫被捕食的数量。每个线虫密度为1个处理,共5个处理,每个处理5次重复。试验数据用Holling[38]圆盘方程Na=aTN0/(1+aThN0)进行拟合,式中N0为供试猎物初始密度,Na为猎物被捕食数量,a为瞬间攻击系数,T为试验持续时间(本试验中T为1 d),Th为捕食者处理1头猎物所需要的时间。将N0及Na数据带入圆盘方程,运用最小二乘法计算,得到各功能反应的反应参数,并用a/Th值来评价巴氏新小绥螨的捕食能力。
1.5 饥饿时间和温度对巴氏新小绥螨捕食水稻干尖线虫的影响
在每个饲养小室分别放入 400条水稻干尖线虫,然后进行以下两个试验:(1)在每个饲养小室中分别放入1头经饥饿处理1、2、3、4和5 d的巴氏新小绥螨雌成螨。在(25±1)℃条件下黑暗培养1 d后观察记录水稻干尖线虫被捕食的数量。每个饥饿时间处理的巴氏新小绥螨在每个线虫密度条件下的捕食量为1个试验处理,共25个处理,每个处理5次重复;(2)将培养箱的温度分别设为15 ℃、20 ℃、25 ℃、30 ℃、35 ℃,在每个饲养小室中分别放入1头饥饿处理1 d的巴氏新小绥螨雌成螨,在黑暗培养1 d后观察记录水稻干尖线虫被捕食的数量。每个温度处理的巴氏新小绥螨在每个线虫密度条件下的捕食量为 1个试验处理,共25个处理,每个处理5次重复。
1.6 数据统计与分析
试验数据的统计分析采用SPSS 14.0和EXCEL 2013软件,平均值和标准误的分析采用单因素方差分析(One-Way ANOVA),多重比较采用Tukey法,双重比较采用t检验,差异显著水平为p=0.05。
2 结果与分析
2.1 巴氏新小绥螨以水稻干尖线虫和腐食酪螨为食的发育繁殖
供试捕食螨分别以两种不同的供试猎物为食的发育历期及生殖力测定结果表明(表1,表2),通过捕食水稻干尖线虫和腐食酪螨,巴氏新小绥螨能够正常发育并完成整个生活史。巴氏新小绥螨从卵发育到成螨一共经历5个阶段,依次为卵、幼螨、前若螨、后若螨和成螨。与取食腐食酪螨相比,巴氏新小绥螨取食水稻干尖线虫时的前若螨历期2.59 d显著短于取食腐食酪螨的2.08 d(p<0.05),其他发育历期差异不显著(p>0.05)。以这两种不同的猎物喂食的巴氏新小绥螨整个发育历期差异不显著(p>0.05)。除此之外,巴氏新小绥螨取食水稻干尖线虫时,产卵前期4.41 d显著小于取食腐食酪螨的4.83 d(p<0.05),产卵期、产卵后期和雌螨寿命均与取食腐食酪螨的差异不显著(p>0.05);日均产卵量大于取食腐食酪螨,但差异不显著(p>0.05)。

表1 巴氏新小绥螨以水稻干尖线虫和腐食酪螨为食的发育历期Table 1 The developmental duration of Neoseiulus barkeri fed on Aphelenchoides besseyi and Tyrophagus putrescentiae

表2 以水稻干尖线虫和腐食酪螨为食的巴氏新小绥螨产卵前期、产卵期、产卵后期、寿命和日均产卵量Table 2 Pre-oveposition, ovipositon, post-oviposition, oviposition amount, longevity and daily oviposition of Neoseiulus barkeri fed on Aphelenchoides besseyi and Tyrophagus putrescentiae
2.2 巴氏新小绥螨以水稻干尖线虫和腐食酪螨为食的生命表及其参数
根据试验获得的数据分别计算供试捕食螨以两种不同的供试猎物为食的存活率及平均产雌数,分别制作得到捕食螨以两种不同猎物为食的生命表,求得生命表参数(表 3),巴氏新小绥螨以水稻干尖线虫为食的净增殖率、内禀增长率和周限增长率分别为31.4605、0.2313/d和1.2603,均大于取食对照腐食酪螨的净增殖率、内禀增长率和周限增长率(R0=26.8800;rm=0.1973/d;λ=1.2181);取食水稻干尖线虫的巴氏新小绥螨种群的平均世代周期(T=19.9342 d)和种群倍增时间(Dt=2.9962 d)均小于取食对照腐食酪螨的种群平均世代周期(T=20.8423 d)和种群倍增时间(Dt=3.5126 d)。
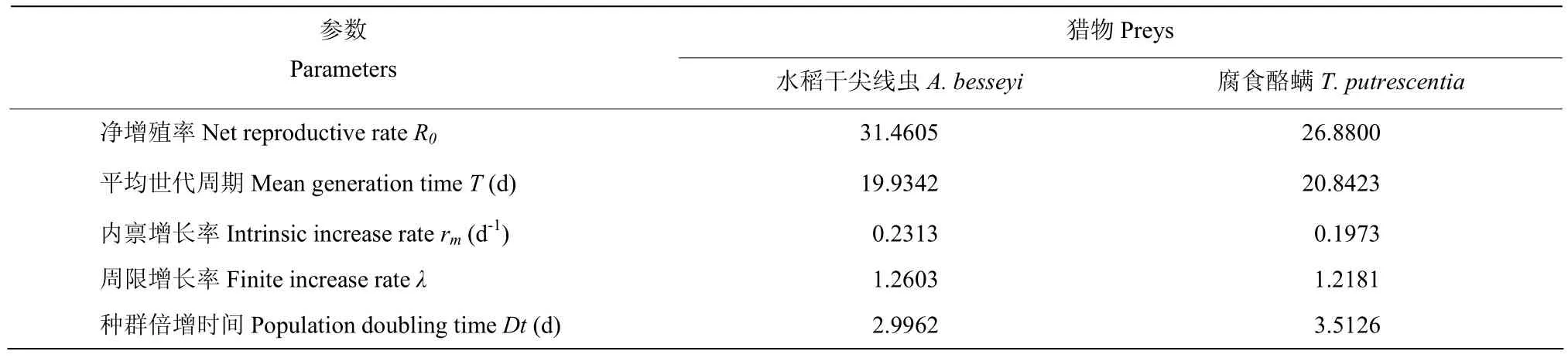
表3 巴氏新小绥螨雌成螨以水稻干尖线虫和腐食酪螨为食的生命表参数Table 3 The life table parameters of Neoseiulus barkeri fed on Aphelenchoides besseyi and Tyrophagus putrescentiae
2.3 巴氏新小绥螨捕食水稻干尖线虫的功能反应
在不同的线虫密度条件下,饥饿1 d的巴氏新小绥螨对水稻干尖线虫的捕食量随着线虫密度的增加而增大,但捕食量的增加速率逐渐降低,捕食量趋于稳定(图1)。当水稻干尖线虫密度为100~400条范围时,巴氏新小绥螨对线虫的捕食数量随着线虫密度的增加而显著增加(p<0.05);水稻干尖线虫密度为400和500条时,巴氏新小绥螨对线虫的捕食量无显著差异(p>0.05)。因此,巴氏新小绥螨捕食水稻干尖线虫的功能反应为负加速模型,属于逆密度制约关系,可用Holling-Ⅱ型圆盘方程Na=aTN0/(1+aThN0)进行功能反应拟合。结果表明,巴氏新小绥螨对水稻干尖线虫的瞬时攻击率a=0.5645,处理时间Th=0.0044 d,捕食能力a/Th=128.2954,圆盘方程公式为Na=0.5645N0/(1+0.002484N0)。理论捕食量与实际捕食量相关系数 R2为 0.9990,经过卡方适合性检验后发现误差不显著(χ2=0.0921<χ20.05=9.49),说明试验估算出的理论模型拟合良好,能够客观地反映出巴氏新小绥螨对不同密度水稻干尖线虫的捕食情况。当线虫密度N0→∞时,1头巴氏新小绥螨在1 d中,捕食水稻干尖线虫的最大理论值为227.27条。
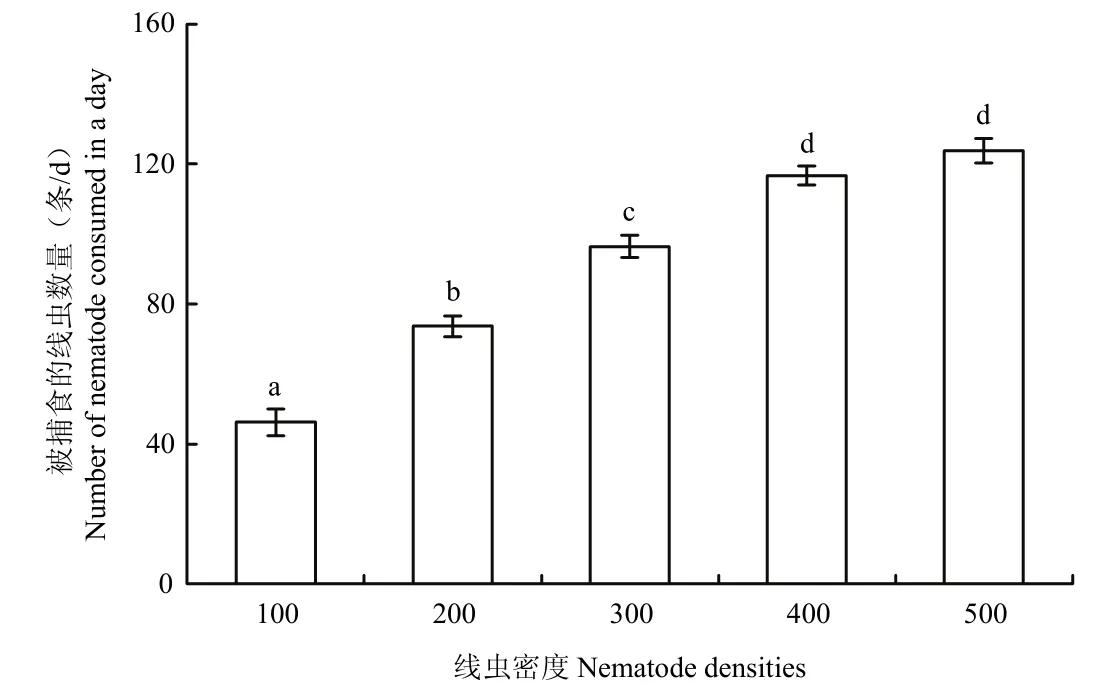
图1 线虫密度对巴氏新小绥螨捕食水稻干尖线虫的影响Fig.1 Effects of nematode density on the predation of Aphelenchoides besseyi by an adult female Neoseiulus barkeri
2.4 饥饿程度对巴氏新小绥螨捕食水稻干尖线虫的影响
在水稻干尖线虫密度为400条的条件下,巴氏新小绥螨对水稻干尖线虫的捕食量随饥饿时间的延长呈先增加后减少的趋势(图2),在饥饿为4 d时,捕食量达到最大,达到165条/d。在饥饿时间为1~3 d范围时,随着饥饿时间的延长巴氏新小绥螨对水稻干尖线虫的捕食量显著增加(p<0.05);饥饿时间为3 d和4 d时,巴氏新小绥螨对水稻干尖线虫的捕食量无显著差异,饥饿时间为5 d时,巴氏新小绥螨对水稻干尖线虫的捕食量显著降低(p<0.05)。
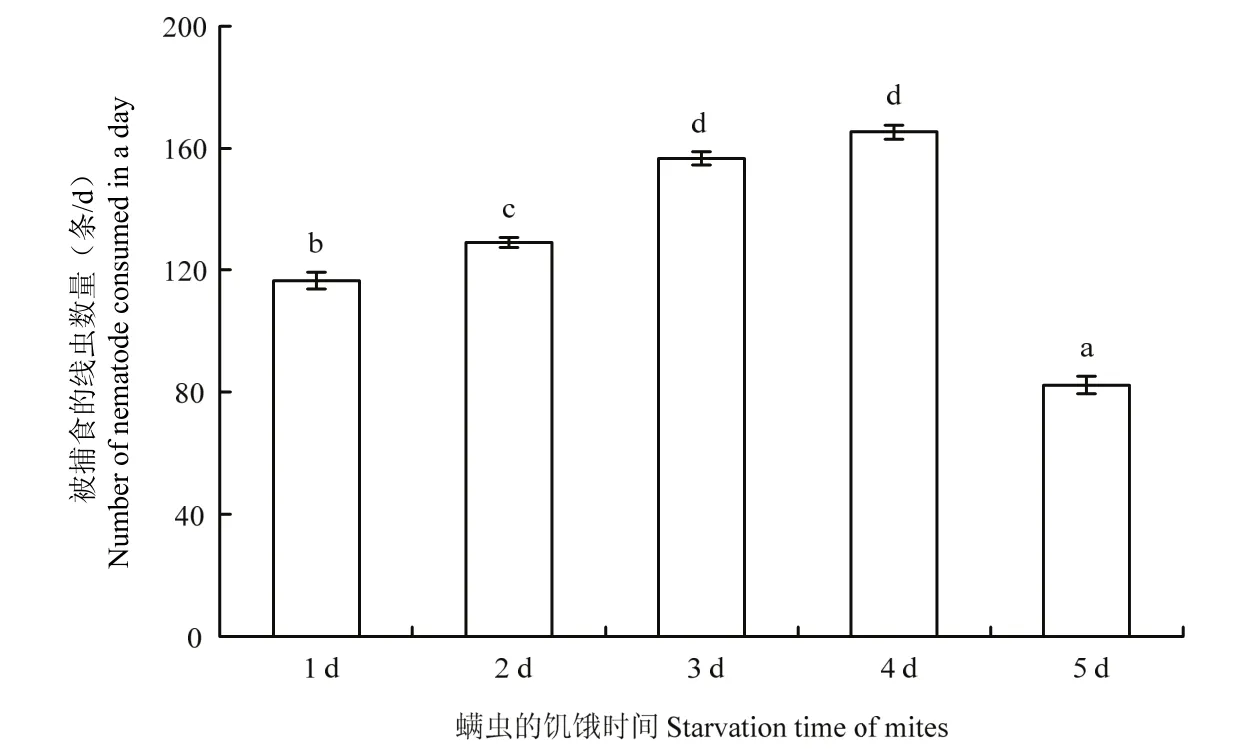
图2 饥饿时间对巴氏新小绥螨捕食水稻干尖线虫的影响Fig.2 Effects of starvation time on the predation of Aphelenchoides besseyi by an adult female Neoseiulus barkeri
2.5 温度对巴氏新小绥螨捕食水稻干尖线虫的影响
水稻干尖线虫密度为400条时,巴氏新小绥螨对水稻干尖线虫的捕食量随温度的升高呈先增加后减少的趋势(图3),在处理温度为25 ℃时,捕食量达到最大,达到117条/d。处理温度在1 ℃~25 ℃范围时,巴氏新小绥螨对水稻干尖线虫的捕食量随温度升高而逐渐升高,15 ℃处理的线虫捕食量显著小于 20 ℃和25 ℃处理(p<0.05),20 ℃和25 ℃处理的线虫捕食量差异不显著(p>0.05);处理温度在25 ℃~35 ℃范围时,巴氏新小绥螨对水稻干尖线虫的捕食量随温度升高而显著降低(p<0.05)。
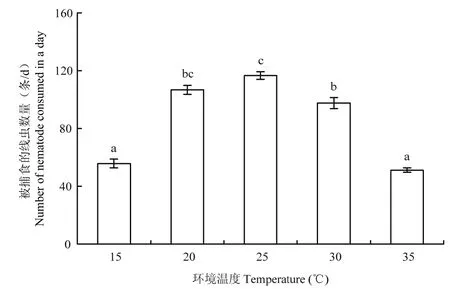
图3 温度对巴氏新小绥螨捕食水稻干尖线虫的影响Fig.3 Effects of temperature on the predation of Aphelenchoides besseyi by an adult female Neoseiulus barkeri
3 讨论
本研究分别以水稻干尖线虫和腐食酪螨为巴氏新小绥螨的猎物,构建巴氏新小绥螨生命表,计算其生命表参数,发现巴氏新小绥螨捕食水稻干尖线虫和腐食酪螨均可正常发育并完成整个生活史,其各阶段的发育历期、寿命及日均产卵量没有明显差异,但前者的前若螨期显著长于后者。这一结果与周万琴等[34]报道巴氏新小绥螨以南方根结线虫为食时的前若螨期长于取食腐食酪螨时的前若螨期的结果一致。除此之外,巴氏新小绥螨以水稻干尖线虫为食的产卵前期显著短于取食腐食酪螨时的产卵前期,其取食水稻干尖线虫时的净增值率、内禀增长率及周限增长率均大于取食腐食酪螨,但其种群倍增时间小于取食腐食酪螨,表明巴氏新小绥螨取食水稻干尖线虫比取食腐食酪螨更适合其生长发育和繁殖。内禀增长率是反应种群在理想条件下的繁殖能力,周限增长率反应的是种群在一定时间内的增长情况[37]。若种群的繁殖能力较强,其内禀增长率及周限增长率也相对较大。本研究结果显示,巴氏新小绥螨取食水稻干尖线虫的内禀增长率和周限增长率(rm=0.23/d,λ=1.26)不仅大于取食腐食酪螨(rm=0.20/d,λ=1.22),而且大于取食南方根结线虫2龄幼虫的内禀增长率和周限增长率(rm=0.18/d,λ=1.20)[34]、取食腐烂茎线虫的内禀增长率和周限增长率(rm=0.16 /d,λ=1.16)[35]以及取食西花蓟马的内禀增长率和周限增长率(rm=0.19/d,λ=1.21)[30]。因此,巴氏新小绥螨以水稻干尖线虫为食具有较强的增殖能力。种群倍增时间可反应种群数量加倍的速率。巴氏新小绥螨以水稻干尖线虫为食的种群倍增时间(Dt=3.00 d)不仅小于对照组取食腐食酪螨(Dt=3.51 d),而且小于文献报道的该螨以南方根结线虫2龄幼虫(Dt=3.76 d)、腐烂茎线虫(Dt=4.56 d)及西花蓟马(Dt=4.64 d)为食[30,34,35]的种群倍增时间。因此,巴氏新小绥螨以水稻干尖线虫为食的种群数量能够快速增加。这些结果表明,巴氏新小绥螨具有作为水稻干尖线虫天敌的潜能。
捕食者与被捕食者的相互作用受被捕食者密度、饥饿时间和环境温度的影响。研究结果表明,在一定范围内,随着线虫密度的增加,巴氏新小绥螨对水稻干尖线虫的捕食量显著增加,但增加速率逐渐降低,猎物密度400条和500条两个处理的捕食量差异不显著,表明猎物密度达到400条后,捕食量趋于稳定,这与捕食螨对猎物最常见的捕食反应形式相似[39]。有研究表明巴氏新小绥螨对柑橘始叶螨Eotetranychus kankitus[40]和腐烂茎线虫[35]等的功能反应均可以用Holling-II模型拟合。本研究结果表明,巴氏新小绥螨对水稻干尖线虫的捕食功能反应为负加速模型,属于逆密度制约关系,同样可以使用 Holling-II型圆盘方程进行功能反应拟合。巴氏新小绥螨对水稻干尖线虫的捕食能力a/Th达128.30,大于巴氏新小绥螨对柑橘始叶螨幼虫的捕食能力(a/Th=68.40)[40]和腐烂茎线虫的捕食能力(a/Th=63.68)[35]。此外,当线虫密度N0→∞时,1头巴氏新小绥螨在1 d中,捕食水稻干尖线虫的最大理论量为227.27条。证明巴氏新小绥螨具有较强捕食水稻干尖线虫的能力。
在一定范围内,巴氏新小绥螨捕食水稻干尖线虫的数量随饥饿时间和温度的增加而增加,但到达一定程度后,随着饥饿时间和温度的持续增加,捕食量降低。Yang等[36]报道巴氏新小绥螨是香蕉穿孔线虫的天敌捕食螨,该研究中的饥饿时间和温度对巴氏新小绥螨捕食香蕉穿孔线虫的影响趋势与本研究一致。在本研究中,饥饿4 d,巴氏新小绥螨对水稻干尖线虫的捕食量达到最大,饥饿超过4 d后,捕食量显著降低(p<0.05)。Yang等[36]报道的巴氏新小绥螨捕食香蕉穿孔线虫的最佳饥饿时间也为4 d。这可能是由于长期饥饿会使螨类消耗过多,导致它们移动、搜索和处理猎物的能力降低。在供试的温度范围内,25 ℃是巴氏新小绥螨捕食水稻干尖线虫的最佳捕食温度。据报道,水稻干尖线虫的适宜活动温度为20 ℃~26 ℃[41],巴氏新小绥螨适合生长温度范围为15 ℃~37 ℃[42],均与巴氏新小绥螨捕食水稻干尖线虫的最佳捕食温度一致,这有利于巴氏新小绥螨对水稻干尖线虫的控制。因此,本研究证实了巴氏新小绥螨对水稻干尖线虫具有较强的捕食作用和控制能力,可以作为水稻干尖线虫潜在天敌捕食螨。巴氏新小绥螨对水稻干尖线虫的防控效果和应用方法尚有待研究。
