江原钝绥螨对三种橘园害虫的捕食能力
2021-08-11李晓杨赵文娟郑薇薇张宏宇
李晓杨,赵文娟,郑薇薇,张宏宇
(华中农业大学园艺植物生物学教育部重点实验室,武汉 430070)
柑橘是我国南方最主要的水果之一,全国有20个省、市、自治区1000多个县种植柑橘,产区大多气候高温多雨、温暖湿润,加之柑橘物候期长,使得柑橘病虫害种类繁多、发生为害较重[1]。近年来随着化学农药的广泛使用,尤其对高毒化学农药的不合理使用,害虫抗药性不断提高,使得原来的一些次要害虫上升为主要害虫[2,3],螨类、粉虱、蚧类、蓟马和蚜虫等“五小害虫”发生为害日趋严重,对我国柑橘绿色无公害生产造成严重威胁[4]。其中,害螨在全国各地柑橘园中普遍发生较重,尤其以柑橘全爪螨PanonychuscitriMcGregor最为严重。柑橘全爪螨是世界性的柑橘害螨[5-7],在我国广泛分布于各柑橘种植区,其幼螨、若螨、成螨均能吸食柑橘叶片、嫩枝、幼果等部位的汁液,使受害部位由绿转白,更可造成落叶、落花、落果,在生产上造成巨大损失[8],是我国柑橘园最重要的害虫之一。柑橘粉虱DialeurodescitriAshm是我国柑橘的主要害虫,以成虫、若虫吸食植物汁液、分泌大量蜜露和诱发霉污病等影响柑橘生产[9,10]。黄胸蓟马ThripshawaiiensisMorgan是我国危害柑橘的主要蓟马种类之一,产卵于植物幼嫩组织内, 若虫在植物花或嫩梢内取食,蛹期入土化蛹,成虫聚集于花或植物幼嫩器官内[11]。
生物防治作为一种安全、无污染的防治手段日益受到重视。捕食螨是柑橘园小型害虫的重要天敌之一,除捕食叶螨、瘿螨、跗线螨外还可以捕食其他小型的无脊椎动物,如蓟马、蚧壳虫、小蛾类的卵和线虫等。捕食螨具有个体小、发育快、生殖潜能大、捕食能力强、易饲养和地理分布广等优势,对害螨的生物防治具有重要的意义[12]。江原钝绥螨AmblyseiuseharaiAmitaietSwirski隶属植绥螨科Phytoseiidae,是华中地区橘园中的捕食螨优势种,主要以柑橘全爪螨为猎物,饥饿状态下也可以取食蚧壳虫虫卵、柑橘粉虱卵和低龄若虫[13,14],分布于柑橘、茶树等多种作物上,是极具潜能的生物防治天敌。利用捕食螨控制害螨是近年来柑橘害螨绿色防控的重要措施[15,16],但在田间释放之前,江原钝绥螨的捕食能力仍需进一步的研究。
天敌捕食作用研究中常通过功能反应来讨论其优劣,对捕食者的评价应根据其环境适应能力、功能和数值反应以及时空同步下与猎物的关系来进行。功能反应即单个捕食者在单位时间内在不同猎物密度下所能捕获的猎物数量变化,是研究天敌对害虫捕食能力的重要尺度之一[17],分为四种基本类型[18]。目前已研究过的捕食螨功能反应大多属于HollingⅡ型,如巴氏钝绥螨AmblyseiusbarkeriHughes雌成螨、雄成螨和若螨在不同温度下对椭圆食粉螨AleuroglyphusovatusTroupeau和柑橘全爪螨的功能反应均属于HollingⅡ型[19]。胡瓜钝绥螨NeoseiuluscucumerisOudermans、尼氏真绥螨EuseiusnicholsiEhara和巴氏钝绥螨对柑橘全爪螨[20,21]、朱砂叶螨TetranychuscinnabarinusBoisduval[22]和西花蓟马Frankliniellaoccidentalis[23-25]、烟蓟马ThripstabaciLind[26]等的功能反应也都符合HollingⅡ型。少数植绥螨的功能反应属于其他类型,例如,真桑钝绥螨AmblyseiusmakuwaEhara的功能反应符合Ⅰ型[27]。数值反应是捕食者数量与猎物密度变化之间的关系,主要有3种方式:行为数值反应、发育数值反应和繁殖数值反应[28]。张艳璇[20]用数值反应和实验种群生命表分析胡瓜钝绥螨对柑橘全爪螨的控制能力,发现在21 ℃~31 ℃和1~9头猎物密度条件下,随着温度升高胡瓜钝绥螨的捕食量增大;同一温度条件下,随着猎物密度增加捕食量增大,均呈正相关关系。
前人的研究主要集中在江原钝绥螨与柑橘全爪螨的相互关系上,重点是以柑橘全爪螨为猎物的发育、繁殖潜能及捕食功能等研究。本文综合研究了江原钝绥螨在不同温度下对柑橘全爪螨、柑橘粉虱、黄胸蓟马的捕食功能反应,以及其对3种猎物的数值反应和选择性,综合上述研究分析其捕食功能,为有效地利用江原钝绥螨防治害螨提供重要的理论依据和现实指导。
1 材料与方法
1.1 供试材料
1.1.1 供试昆虫(螨) 试验所用江原钝绥螨的螨源采自湖北省宜昌市当阳农户的柑橘树上,置于25 ℃,相对湿度RH 85%,光周期16L∶8D条件下以柑橘全爪螨为食进一步饲养纯化而得。柑橘全爪螨采自华中农业大学柑橘园的柑橘树上,饲养并建立试验种群,作为供试材料。黄胸蓟马采自华中农业大学柑橘园的柑橘树上,饲养并建立种群,取初孵若虫进行试验。柑橘粉虱采自华中农业大学柑橘园的柑橘树上,饲养并建立种群,取初孵若虫进行试验。
1.1.2 试验仪器和材料 人工智能气候培养箱、双筒解剖镜、0号小毛笔、大培养皿(直径6 cm,高1.5 cm)、小培养皿(直径3 cm,高1 cm)、滤纸、记号笔、海绵、黑色塑料薄膜。
1.2 江原钝绥螨对不同猎物的功能反应
功能反应试验在恒温培养箱内进行,供试温度分别为16 ℃、20 ℃、24 ℃、28 ℃、32 ℃,RH 85%±5%,光周期16L∶8D。
试验采用的方法是水盘法,在培养皿(直径90 mm)内,直径由大到小,依次放入剪成圆形的海绵、滤纸、黑膜,海绵和滤纸用水充分浸透,用昆虫针在黑膜边缘扎小孔,培养皿内保持一定水位,约为海绵厚度的1/2。
在各供试温度下,每个小饲养水盘上放 1 头经 24 h 饥饿处理的江原钝绥螨雌成螨,柑橘全爪螨(成螨/幼若螨/卵三种螨态分别处理)和柑橘粉虱(1龄若虫)的密度设置为5、10、15、20、25、30头;黄胸蓟马(1龄若虫)密度设置为3、5、10、15、20、25头,每个处理重复10次,对照组不放入捕食螨,24 h后记录其捕食量。
1.3 江原钝绥螨对不同猎物的数值反应
数值反应试验在恒温培养箱25 ℃,RH 85%±5%,光周期16L∶8D条件下进行,试验前将雌成螨与同种的雄成螨放在同一小试管里,用供试猎物饲喂两天,再饥饿24 h后转移至试验器皿中。每天观察时将雌成螨当日所产卵取出,如有剩余猎物及时清除,然后加入规定的猎物数,记录捕食者产卵量和死亡数。连续观察10 d。柑橘全爪螨(成螨)密度设置为1、3、6、9、12、15头;柑橘粉虱(1龄若虫)密度设置为5、10、15、20、25、30头;黄胸蓟马(1龄若虫)密度设置为3、5、10、15、20、25头。每个处理重复6次,对照组不放入捕食螨。
1.4 江原钝绥螨对不同猎物的选择性
1.4.1 江原钝绥螨对柑橘全爪螨不同螨态的选择性 往培养皿中挑入各螨态柑橘全爪螨各 20头和经饥饿24 h处理后的江原钝绥螨雌成螨1头,24 h后观察记录江原钝绥螨雌成螨对猎物各螨态的日捕食量,每处理设10次重复,温度设置为25 ℃,RH 85%±5%。(螨态根据柑橘全爪螨的日龄判断,一般在25 ℃±1 ℃及相对湿度为75%~85%的条件下,卵期2 d,幼螨期、前若螨期和后若螨期各1 d)。
1.4.2 江原钝绥螨对不同猎物的选择性 往培养皿中挑入柑橘全爪螨成螨、黄胸蓟马(1龄若虫)、柑橘粉虱(1龄若虫)各20头和经饥饿24 h处理后的江原钝绥螨雌成螨1头,24 h 后观察记录江原钝绥螨雌成螨对三种猎物的日捕食量,每处理设10次重复,温度设置为25 ℃,RH 85%±5%。
1.5 数据统计与分析
1.5.1 功能反应 功能反应是一个捕食者在单位时间内对猎物的捕食量随猎物的不同密度而变化。Holling根据对功能反应的研究提出了圆盘方程,其中HollingⅡ型圆盘方程为:Na=aTN/(1+aThN)[18]。式中,N为猎物的初始密度,Na为猎物被捕食数量,a为天敌瞬间攻击系数,T为捕食者可利用以发现猎物的时间,本试验中T为l d,Th为处理1头猎物所需要的时间,a/Th用来评价捕食者的捕食能力,1/Th为日最大捕食量。
1.5.2 选择性 天敌对害虫(螨)嗜食性的高低用选择系数Q来表示:Q=R1/R2(其中R1表示某猎物被捕食数与被捕食总数之比;R2表示某猎物数与猎物总数之比。若Q>1时,表示天敌对该猎物是嗜食的;若Q<1时,表明是非嗜食的;而Q=1时,表明天敌对该猎物是随机捕食的[21]。所有数据分析利用Excel 2007和SPSS 19.0统计软件完成。
2 结果与分析
2.1 江原钝绥螨对不同猎物的功能反应
江原钝绥螨雌螨在不同温度下对不同密度的柑橘全爪螨各螨态、柑橘粉虱1龄若虫、黄胸蓟马1龄若虫的捕食功能反应结果所作散点趋势图见图1~3。图中结果表明:在同一温度处理内,江原钝绥螨对3种猎物的捕食量随猎物密度的增加而增加,但当猎物密度增加到一定限度后,江原钝绥螨的捕食量增幅逐渐减缓;且在同一温度、同一密度处理下,江原钝绥螨对柑橘全爪螨各螨态的捕食量为幼若螨>成螨>卵。在同一密度处理下,在16~28 ℃范围内,随着温度的升高,江原钝绥螨对3种猎物的捕食量逐渐增大,28 ℃时猎物捕食量达到最大,温度高于28 ℃捕食量开始减少,这表明温度过高或过低均会影响江原钝绥螨的捕食能力,28 ℃为江原钝绥螨最佳捕食温度。

图1 不同温度条件下江原钝绥螨雌螨对柑橘全爪螨的捕食功能反应Fig.1 Functional responses of adult females of A.eharai on three stages of P.citri at the different temperatures
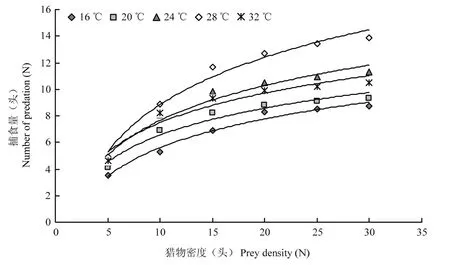
图2 不同温度条件下江原钝绥螨雌螨对柑橘粉虱1龄若虫的捕食功能反应Fig.2 Functional responses of adult females of A.eharai on the first instar larvae of D.citri at the different temperatures

图3 不同温度条件下江原钝绥螨雌螨对黄胸蓟马1龄若虫的捕食功能反应Fig.3 Functional responses of adult females of A.eharai on the first instar larvae of T.hawaiiensis at the different temperatures
江原钝绥螨对柑橘全爪螨、柑橘粉虱和黄胸蓟马的功能反应均属于 HollingⅡ型。用 HollingⅡ圆盘方程Na=TN/(1+aThN) 拟合的功能反应数据如表1所示。在16 ℃~32 ℃内,江原钝绥螨对柑橘全爪螨各螨态、柑橘粉虱1龄若虫和黄胸蓟马1龄若虫均有较强的捕食能力;猎物密度在一定范围内,江原钝绥螨的捕食量随猎物密度的增大而增加,但当猎物增加到一定密度后,其捕食量则在一定范围内波动。在16 ℃~28 ℃范围内,江原钝绥螨对这3种猎物的天敌瞬间攻击系数、控制能力均随着温度上升而增大,猎物处理时间随着温度的上升而缩短,28 ℃时江原钝绥螨对猎物的瞬间攻击系数达到最大值,猎物处理时间最短,控制能力同样显示 28 ℃时江原钝绥螨的捕食能力最强。当温度上升到 32 ℃,江原钝绥螨对猎物的瞬间攻击系数和控制能力均明显下降,这表明温度过高或过低均会影响江原钝绥螨的捕食能力,当温度达到28 ℃时,江原钝绥螨捕食能力最强。

表1 江原钝绥螨雌成螨对柑橘全爪螨、柑橘粉虱和黄胸蓟马的功能反应方程和参数Table 1 Functional responses equation and parameters of adult females of A.eharai on P.citri, D.citri and T.hawaiiensis
2.2 江原钝绥螨对不同猎物的数值反应
江原钝绥螨对柑橘全爪螨(成螨)、柑橘粉虱(1龄若虫)和黄胸蓟马(1龄若虫)的繁殖数值反应见表2。表中结果显示江原钝绥螨的日均产卵量在低猎物密度下随着猎物密度的增加而显著增加;在高猎物密度下,随着猎物的密度增加,日均产卵量增加的速率逐渐减慢,没有显著性增加。江原钝绥螨在捕食柑橘全爪螨、柑橘粉虱和黄胸蓟马时的日均产卵量与猎物密度均能用HollingⅡ型模型拟合,其方程分别如下。柑桔全爪螨:Na=1.1210N/(1+0.7143N)(R²=0.9476);柑橘粉虱:Na=0.0890N/(1+0.0299N)(R²=0.9907);黄胸蓟马:Na=0.2391N/(1+0.1158N)(R²=0.978)。

表2 不同江原钝绥螨密度下捕食柑橘全爪螨、柑橘粉虱和黄胸蓟马的日均产卵量Table 2 The reproduction number of P.citri, D.citri and T.hawaiiensis under different density of A.eharai
2.3 江原钝绥螨对猎物的选择性
2.3.1江原钝绥螨对柑橘全爪螨不同螨态的选择性 江原钝绥螨雌成螨对柑橘全爪螨各螨态选择捕食作用数据见表 3。江原钝绥螨对柑橘全爪螨各螨态的嗜食性显著不同。江原钝绥螨雌成螨对柑橘全爪螨的幼螨和若螨的选择系数Q分别为2.21和l.40,均大于1,而对柑橘全爪螨的卵和成螨的选择系数分别为0.14和0.25,均小于 1。结果表明,柑橘全爪螨的幼螨、若螨是江原钝绥螨的嗜食螨态,且幼螨为最嗜好虫态。当柑橘全爪螨各螨态同时存在时,江原钝绥螨会优先捕食柑橘全爪螨幼螨和若螨。
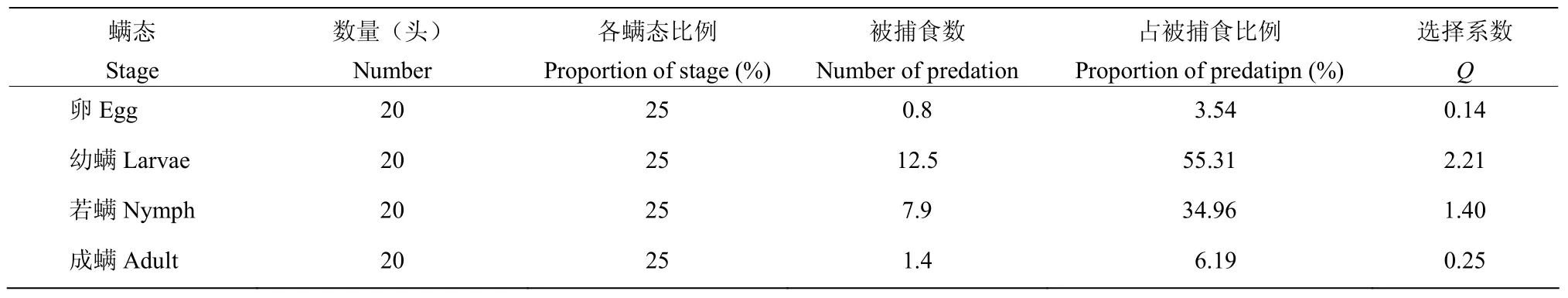
表3 江原钝绥螨雌成螨对柑橘全爪螨各螨态的选择捕食作用Table 3 Prey stage preference of adult females of A.eharai to P.citri
2.3.2 江原钝绥螨对不同猎物的选择性 江原钝绥螨雌成螨对柑橘全爪螨成螨、柑橘粉虱1龄若虫、黄胸蓟马1龄若虫这3种猎物的选择捕食作用数据见表4。结果表明,江原钝绥螨雌成螨对不同猎物的嗜食性显著不同。在3种猎物中江原钝绥螨对柑橘全爪螨最嗜食,选择系数为3.29(Q>1)。而江原钝绥螨雌成螨对柑橘粉虱和黄胸蓟马的选择系数分别为0.09和0.63,均小于1。在这3种猎物都存在的情况下,江原钝绥螨的主要捕食对象为柑橘全爪螨,其次为黄胸蓟马。

表4 江原钝绥螨雌成螨对柑橘全爪螨、柑橘粉虱和黄胸蓟马的选择捕食作用Table 4 Prey preference of adult females of A.eharai to P.citri, T.hawaiiensis and D.citri
3 讨论
本研究表明,江原钝绥螨对柑橘全爪螨各螨态、柑橘粉虱、黄胸蓟马的功能反映均符合Holling II型圆盘方程,这与前人研究中江原钝绥螨雌成螨对烟粉虱卵的捕食功能反应一致[14]。大多数捕食螨对叶螨的捕食功能反应均符合 Holling II 型圆盘方程,如加州新小绥螨NeoseiuluscalifornicusMcGregor对苹果全爪螨PanonychusulmiKoch[29]、朱砂叶螨[30],巴氏钝绥螨对二斑叶螨TetranychusurticaeKoch[31]、柑橘全爪螨[21]。江原钝绥螨对柑橘全爪螨、柑橘粉虱和黄胸蓟马这三种猎物均具有较强的捕食能力。
本研究发现温度是影响江原钝绥螨捕食效能的重要因素,在温度16 ℃~28 ℃,江原钝绥螨对这三种猎物的控制能力均随温度升高而增大,在28 ℃时达到最大值,其后控制能力随温度升高而下降。说明江原钝绥螨在 28 ℃时具有较高的捕食效能,温度过高会减弱江原钝绥螨对猎物的捕食效能,这可能是因为高温下江原钝绥螨活性降低,前人的研究也表明当温度达到32 ℃会引起江原钝绥螨死亡[13]。季洁等[14]研究了江原钝绥螨对烟粉虱卵的捕食能力,在16 ℃~28 ℃,其捕食能力随温度升高而增大,但当温度达到32 ℃时,捕食量明显下降,该结果与本研究结果基本一致。张艳璇[20]研究了胡瓜钝绥螨对柑橘全爪螨的捕食能力,在27 ℃~28 ℃时胡瓜钝绥螨的捕食能力最高。凌鹏等[21]研究了巴氏钝绥螨对柑桔全爪螨的捕食能力,在28 ℃时巴氏钝绥螨的捕食能力最高。以上研究表明捕食螨捕食效能均受温度影响。
江原钝绥螨对猎物的选择性试验结果表明,在柑橘全爪螨、柑橘粉虱和黄胸蓟马这3种猎物共同存在的情况下,江原钝绥螨会优先捕食柑橘全爪螨。通过本试验的研究结果,可以发现江原钝绥螨主要捕食柑橘全爪螨,同时也能捕食一些柑橘粉虱和黄胸蓟马。可以推测,当柑橘全爪螨缺乏时,江原钝绥螨会捕食柑橘粉虱和蓟马来维持种群的稳定,从而稳定自身种群数量,达到长期控制柑橘全爪螨为害的目的。江原钝绥螨雌成螨对柑橘全爪螨各螨态的取食具有选择性,其幼若螨是江原钝绥螨的嗜好螨态。可能是由于幼若螨个体小,行动迟缓,所需的处理时间短;而成螨个体大,行动较快,所需的处理时间长;卵有坚硬而相对光滑的卵壳保护,不利于江原钝绥螨的取食。江汉华等[32]、陈守坚等[33]通过研究分别发现,江原钝绥螨在对柑橘全爪螨取食时,均偏好柑橘全爪螨的幼若螨,与本试验的选择性结果基本一致。大部分捕食螨对害螨的卵、幼螨、若螨有嗜食性,对成螨无嗜食性,如加州新小绥螨对苹果全爪螨卵、幼螨和第1若螨有嗜食性,对苹果全爪螨的第2若螨和雌成螨无嗜食性[29];巴氏钝绥螨对柑橘全爪螨幼螨和若螨有嗜食性[20];巴氏钝绥螨对二斑叶螨的卵和若螨表现为嗜食,对雌成螨表现为非嗜食[31]。
