松材线虫病除治迹地营造的15年生木荷林生长分析*
2021-08-11陈献志楚秀丽黄盛怡程卫斌冯贻勇王斌周志春
陈献志,楚秀丽,黄盛怡,程卫斌,冯贻勇,王斌,周志春
(1.浙江省临海市自然资源和规划局,浙江 临海 317000;2.中国林业科学研究院亚热带林业研究所,浙江 杭州 311400;3.临海市括苍镇农业综合服务中心,浙江 临海 317000;4.临海市涌泉镇农业综合服务中心,浙江 临海 317000)
松材线虫病(Bursaphelenchusxylophilus)是世界上最危险的森林病害之一[1],不仅影响社会经济的发展,而且也严重破坏自然景观和生态环境[2-3],是林业上必须高度重视预防的重大病虫害。国内外学者从化学措施防治、物理防治、生物防治及抗病育种等方面对松材线虫防治进行了大量的研究,但由于其传播、发生及危害的特殊性,现有的防控技术还不能很好地控制松材线虫病的蔓延,导致我国松材线虫病发生面积不断扩大[4]。干净、彻底地清理松材线虫病病死树是目前最经济、最有效的防治措施,由此形成大面积的松材线虫病除治迹地。立地条件较差的除治迹地,其植物群落生物多样性降低,垂直分层结构不合理,只剩灌木层和草本层,原有松林向灌丛方向退化,生态功能显著下降,受损的群落很难自然恢复与更新[5]。因此,尽快恢复除治迹地的森林植被,将其改培为生产力更高、生态功能优质的森林,是当前我国森林建设的重要内容之一。
补植阔叶树是目前各地普遍采用的松材线虫病除治迹地生态修复措施,通过有效的人工造林,可极大加快除治迹地植被恢复进程[6]。在此过程中,生态修复树种的选择与配置模式起着至关重要的作用。适于我国亚热带区域马尾松(Pinusmassoniana)林生态修复的阔叶树种有木荷(Schimasuperba)、青冈(Cyclobalanopsisglauca)、枫香(Liquidambarformosana)、杜英(Elaeocarpusdecipiens)、千年桐(Verniciamontana)、火力楠(Micheliamacclurei)和山乌桕(Sapiumdiscolor)等,其中以木荷最具代表性,其与枫香、甜槠(Castanopsiseyrei)、青冈等共同组成复层群落类型,在陡坡和岩石较多的生境中也能形成较为单纯的群落类型[7]。木荷为山茶科(Theaceae)木荷属(Schima)常绿大乔木,喜光,幼年稍耐庇荫,不仅是我国生物防火林带建设的当家树种和重要生态防护树种,而且也是主要的速生、珍贵、优质阔叶用材树种之一[8],同时也是当今松材线虫病除治迹地更新首选的生态修复树种[9],在生态修复补植造林中的表现极佳。枫香作为我国重要的速生、优质乡土树种和亚热带阔叶林中的优良先锋树种,与木荷混交也是生产中常用的一种造林模式。木荷纯植和与枫香混植2种林分具有不同的林内光照条件,木荷的生长是否会因此而有所差异?本研究选择浙江东部松材线虫病除治迹地营造的15 a木荷林,分析其在不同坡位及与枫香混交条件下的生长表现,以期为松材线虫病除治迹地生态修复及木荷高质量林分的培育提供参考。
1 材料与方法
1.1 试验地概况
研究区位于浙江省临海市(120°49′~121°41′E,28°40′~29°04′N),属亚热带季风气候,年平均气温为18.2 ℃,年平均降雨量1 710.4 mm,无霜期241 d,年平均日照时数1 699.3 h,年积温5 370 ℃。境内西北部为山地丘陵,中南部为河谷平原及低丘缓坡,东部为沿海平原及岛屿,海拔跨度较大,在10~1 200 m之间,山地丘陵土壤以红壤为主,河谷平原以水稻土为主。21世纪初,临海市松材线虫病大面积爆发,马尾松林遭受严重危害,因此,以仅保留极少数健康木(约为90 株/hm2)的方式采伐病害木,形成大面积的除治迹地。2005年,临海市在西北部松材线虫病除治迹地上(立地条件中等偏下,海拔50~280 m)栽种1 a生木荷容器苗进行改培(当年造林面积共327.93 hm2),改培模式包括木荷纯林、木荷与枫香1∶1混交林两种模式,造林株行距2 m×3 m。
2019年样地调查显示,改培的木荷林中有天然更新的枫香、青冈和白栎(Quercusfabri)等,地表植被多为地衣、杂灌及草丛。经过15 a的生长,部分木荷和枫香被自然淘汰,2019年调查时纯林模式保留的木荷为825 株/hm2,混交模式保留的木荷与枫香分别为558 株/hm2和456 株/hm2(比例为1.23∶1)。混交林内枫香平均树高9.20 m,平均胸径10.54 cm,平均单株材积0.045 m3,平均树干通直度3.46。
1.2 样地设置与调查
2019年底在2005年实施生态修复的松材线虫病除治迹地内选择海拔150~200 m,林地条件和抚育措施等基本一致的木荷林,按照纯林模式和混交模式,分别在上、中和下坡位设置样地,3次重复,共18块样地,每个样地面积400 m2(20 m×20 m);改培模式(2水平)和坡位(3水平)共同构成复因子完全随机设计,共计6个处理组合(表1)。

表1 样地基本情况Tab.1 General information of the sample plots
每木测定样地内木荷树高、胸径、冠幅、分叉干数、最大分枝粗(最粗的一级分枝基部直径)、最大分枝角(最粗一级分枝与主干的夹角)和树干通直度。
分叉干数取值:单一主干无分叉,记为0;除主干外距基部1 m以下有1个分叉干,记为1,有2个分叉干记为2,依此类推。
树干通直度分为通直、较通直、一般、弯曲、严重弯曲5 级,分别记分为5、4、3、2 和1,数字越大越通直。
1.3 数据处理与分析
木荷单株材积按V=0.000 068 01D1.865 613H0.918 129估算,式中V为单株材积,D为胸径,H为树高[10]。
植株高径比为树高与胸径之比,比值越大树干越圆满;冠径比为树冠直径与胸径之比,称树冠系数,比值越小林木空间利用越好;枝粗指标为最大分枝粗与胸径之比,比值越小林木自然整枝越好。
采用Weibull分布函数对径阶分布进行拟合[11-12]。用d=r/R公式(r为林木胸径,R为林分平均胸径)计算个体的相对胸径d(单木竞争指标)。各样地统计分级木比例,林木分级标准为:Ⅰ级木,d≥1.336;Ⅱ级木,1.026≤d<1.336;Ⅲ级木,0.712≤d<1.026;Ⅳ级木,0.383≤d<0.712;V级木,d<0.383[13]。
采用Microsoft Excel 2018进行描述性统计分析;采用SPSS 25.0单因素方差分析(one-way ANOVA)和Duncan检验法进行方差分析和多重比较(α=0.05)。分叉干数经(X+0.5)1/2数据转换,树干通直度经X1/2数据转换,最大分枝角和各级林木株数比例经反正弦数据转换后进行方差分析[14]。
2 结果与分析
2.1 松材线虫病除治迹地木荷林生长特征分析
表2为松材线虫病除治迹地15 a木荷林生长特征分析结果。从性状变异系数来看,木荷分叉干数变异最大(72.42%),主要是因为木荷林分叉干数分布极度不均,80%以上的木荷林木没有分叉,极少数林木有3个分叉;其次是单株材积(49.23%),该值与树高和胸径有关,而树高和胸径分别受改培模式和坡位影响显著,因此有着较大的变幅;树高、胸径和冠幅变异系数均在20%左右;树干通直度变异系数最小(10.36%),松材线虫病除治迹地种植木荷时因存在部分上层的马尾松,其树干都较为通直。变异来源分析表明,木荷树高、单株材积、最大分枝角、树干通直度和高径比受改培模式影响显著,树高、胸径、单株材积和最大分枝粗受坡位影响显著。改培模式和坡位对最大分枝粗、最大分枝角度和枝粗指标的互作效应显著。可见,除治迹地营造木荷林时,需同时考虑改培模式及坡位的影响,应根据坡位条件选择适合的改培模式,以获得最佳的修复和改培效果。

表2 除治迹地木荷林生长特征分析Tab.2 Analysis on the growth traits and variation of S.superba forests in the clear-cutting forestland
2.2 不同改培模式及不同坡位的木荷林生长性状比较
不同改培模式及坡位下木荷林生长性状的比较分析结果见表3。从改培模式来看,混交模式下木荷树高、胸径、冠幅和单株材积等生长指标较纯林模式分别提高了33.14%、13.55%、10.05%和57.38%,其中树高显著高于纯林模式,表明除治迹地种植木荷时若与早期速生的枫香混交,能明显地促进木荷的生长。样地调查结果显示,混交模式林地后期天然更新的树种有青冈、冬青(Ilexchinensis)、山槐(Albiziakalkora)、苦槠(Castanopsissclerophylla)、白栎(Quercusfabri)、化香(Platycaryastrobilacea)、榉树(Zelkovaserrata)和檫木(Sassafrastzumu)等10余种,而纯林模式林地内天然更新的树种仅青冈、白栎和冬青等少数几种,表明混交模式在促进木荷生长的同时也增加了林内的生物多样性。此外,混交模式的木荷高径比较纯林模式高18.08%,冠径比、分叉干数和枝粗指标较纯林模式分别低1.58%、32.00%和2.44%,而纯林模式的木荷最大分枝角和树干通直度分别较混交模式高14.53%和11.21%,最大分枝粗较混交模式低9.21%,并且树干通直度和高径比在两种模式间存在显著差异。从木荷生长和形质指标综合来看,木荷-枫香混交模式优于木荷纯林模式。因此,在除治迹地营造木荷林时,可选择与枫香混交栽种,不仅能促进木荷林的生长和优质干材培育,而且可提高林分的景观效果。

表3 不同改培模式及坡位对木荷林生长性状影响Tab.3 Effects of plantation modes and slope position on the growth traits of S.superba forests
从不同坡位来看,木荷林树高、胸径、冠幅和单株材积总体随坡位升高呈下降趋势。其中纯林模式下木荷树高、胸径、冠幅和单株材积下坡位显著高于上坡位,混交模式木荷树高下坡位显著高于上坡位。可见,无论是纯林模式还是混交模式,木荷在除治迹地下坡位都有着最好的生长表现。同时,混交模式下木荷胸径、冠幅和单株材积在不同坡位间差异减少,表明与枫香混交一定程度上能够缓解中、上坡位较差立地条件对木荷生长的影响。纯林模式木荷分叉干数、最大分枝角和树干通直度在不同坡位间均无显著差异,仅最大分枝粗下坡位显著高于上坡位,而混交模式的木荷分叉干数、最大分枝粗和树干通直度在不同坡位间均无显著差异,仅最大分枝角下坡位显著高于中坡位。总体来看,坡位对木荷形质影响较小。
从高径比、冠径比和枝粗指标来看,纯林模式木荷高径比随坡位升高逐渐增加,即坡位越高木荷干形越好。与之相反,混交模式木荷高径比随坡位升高逐渐降低。多重比较结果表明,2种模式不同坡位间高径比差异均不显著,但混交模式中、下坡位高径比显著高于同坡位纯林模式,表明混交模式下木荷不仅长得快而且干形好。木荷林冠径比在不同坡位间均无显著差异,表明坡位对林分空间利用无显著影响。纯林模式下木荷枝粗指标随坡位升高逐渐减小,其中上坡位明显低于下坡位,这与上坡位立地条件较差,木荷最大分枝粗较小有关。混交模式下木荷枝粗指标中坡位最低,但不同坡位间无显著差异,可见与纯林模式相比,混交模式下不同坡位木荷的自然整枝能力相对一致。
2.3 木荷林林木分级比例及径阶分布参数
径阶分布反映林分内各大小直径林木按径阶的分配状态,对森林经营具有重要意义。对木荷林径阶分布采用Weibull分布函数进行拟合,各参数计算结果见表4。确定系数R2反映参数拟合程度,其值越接近于1,说明参数对数据的拟合程度越好。在本研究不同改培模式、不同坡位的15 a木荷林中,表示林分最小直径的位置参数a介于3.215~4.514之间;表示林分直径分布范围的尺度参数b介于4.038~5.022之间;决定林分直径分布偏度的性状参数c仅木荷纯林上坡位小于1,为倒“J”型分布,林分结构稳定,其余木荷林c均大于1,为单峰左偏山状分布,林分结构尚不稳定(表4,图1)。

表4 木荷林直径分布参数及林木分级Tab.4 Diameter distribution and classification of S.superba forests
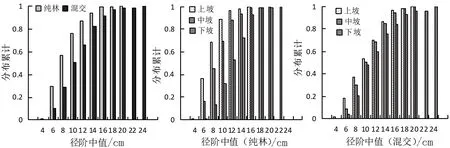
图1 不同改培模式及坡位条件下木荷林直径累积分布Fig.1 Cumulation of diameter distribution of S.superba forests in different mode and slope position
林木分级结果显示,木荷林以Ⅱ、Ⅲ级木为主,其中纯林模式Ⅱ、Ⅲ级木比例为79.15%,混交模式Ⅱ、Ⅲ级木比例为67.71%,均无Ⅴ级木。混交模式中Ⅰ、Ⅳ级木比例高于纯林模式,而Ⅱ、Ⅲ级木比例低于纯林模式,表明除治迹地混交种植木荷和枫香促进了木荷个体分化。不同坡位木荷林Ⅰ、Ⅲ、Ⅳ级木无显著差异,纯林模式下坡位Ⅱ级木显著高于混交模式,但混交模式下坡位Ⅰ级木比例较高,是纯林模式的3.82倍,可见在下坡位优越的立地条件下,混交林中的木荷相比纯林更易促进林木个体分化。
不同改培模式相同分布累积频率范围对应的径阶中值显示,混交模式的径阶中值较纯林模式的更大,再次表明除治迹地木荷-枫香混交的改培模式能够促进木荷的生长(图1)。坡位间相同分布累积频率范围对应的径阶中值表明,不同改培模式木荷径阶中值为下坡>中坡>上坡(图1),与表3中胸径随坡位变化的规律一致。
3 讨论与结论
3.1 讨论
福建建瓯低山丘陵区营造的15 a木荷人工林树高和胸径分别为10.13 m和10.75 cm[15],广东梅州市18 a生木荷人工林平均树高、胸径和单株立木材积分别为7.47 m、8.19 cm和0.024 m3[16],本研究松材线虫病除治迹地营造的15 a木荷林,其生长状况与福建建瓯15 a木荷人工林基本一致,高于广东梅州市18 a木荷人工林。前两者均是采伐迹地造林,本研究相当于林冠下造林,可能早期适度遮阴更有利于木荷人工林生长。
阔叶树与伴生树种混交造林可产生积极或消极影响,选择种间关系协调的伴生树种,不仅可以促进其生长,而且可以改善其干形[17]。本研究木荷分叉干数变异较大,可能与选用容器苗造林有关,也可能与除治迹地改培方式有关。混交改培模式的木荷生长指标和形质指标总体优于木荷纯林改培模式,说明与枫香混交改变了木荷林分生长环境,促进了木荷更好地生长,该结论与木荷-杉木或木荷-马尾松混交生长效益研究结果相一致[18-19]。究其原因,主要是因为混交促进了木荷的竞争生长,并且混交模式对光照和养分的利用更充分[19]。同时,混交种植木荷和枫香的林地,其物种多样性明显高于木荷纯林,马尾松林下套种阔叶树也得到类似结论[20],主要是常绿树种木荷和落叶树种枫香混交,形成了更加多样的林内光照条件,为山槐、苦槠、化香、榉树和檫木等喜光树种提供了适宜的生长环境。可见,在除治迹地栽种木荷时,宜采用与枫香、榉树或檫木等进行混交,不仅能促进木荷的生长,同时也能增加林内的生物多样性,有利于形成更加合理和稳定的多树种森林群落,加速除治迹地植被恢复进程。
立地条件对林木生长的影响主要是通过水肥和光照条件的差异引起的。木荷易成活、易成林,有较强的适应松材线虫病除治迹地立地条件的能力,但不同坡位生长状况存在差异,其中下坡位生长表现最佳,主要是因为下坡位立地条件较好,这与王秀花等[15]对木荷、廖德志等[21]对樟树(Cinnamomumcamphora)、苏妮尔等[22]对红皮云杉(Piceakoraiensis)的研究结论一致。纯林改培模式的木荷生长受坡位影响较大,随着坡位的升高,生长有减弱的趋势,树高、胸径、冠幅和单株材积等生长指标,以及最大分枝粗、枝粗指标等形质指标在上坡位与下坡位之间均存在显著差异。与木荷纯林模式相比,木荷-枫香混交改培模式仅树高、单株材积随着坡位的升高呈现减少的趋势,表明除治迹地营造木荷林时与枫香混交能缓解坡位条件对木荷生长的影响,使其在不同坡位间的生长性状差异有所缩小,该结论与木荷-杉木混交生长效益研究结果相一致[15,18],可能与混交林可调节土壤大量营养元素及有效微量元素状况,改良土壤理化特性有关[23]。这也意味着除治迹地特别是中、上坡位宜采用与枫香混交方式营造木荷林。
倒“J”型分布是林分结构稳定的必要特征之一[24],本研究除木荷纯林上坡位为倒“J”型分布外,其余均为单峰左偏山状,c值远低于楚秀丽等[12]对福建建瓯42 a木荷人工林的研究结果,可见本研究除治迹地营造的15 a木荷林远未达到稳定状态,林分结构随林龄增大将持续发生变化。混交模式c值高于纯林模式,即其林分结构相对更不稳定,表明木荷与枫香混交使得木荷林形成稳定结构的时间后延,此外,纯林模式和混交模式c值均表现为下坡>中坡>上坡,即坡位越低林分结构越不稳定。这主要是因为林地上坡位长期受到雨水冲刷,土层较薄,肥力较弱,而中、下坡位积累流失的土壤和养分,具有更好的水肥条件,因此木荷生长更加旺盛[25],导致形成稳定林分结构的时间更长。不同改培模式及坡位条件下木荷林均以Ⅱ、Ⅲ级木为主,Ⅰ、Ⅳ级木比例较低,均未出现Ⅴ级木,表明林分分化程度还较低[12]。研究表明,Ⅰ、Ⅱ级木在林木状态转移中起主要支配作用,因此林分常遵循培育Ⅰ级木,间伐Ⅱ级木,解放Ⅲ级木,移除Ⅳ、Ⅴ级木的原则[26],对本研究的木荷林而言,这一原则同样适用。随着木荷林分年龄的增长,由于林木间相互竞争与分化,会有大量林木逐渐死亡,为避免林分自然稀疏过程中干形良好个体被淘汰,形质低劣个体被保留,后期应通过抚育间伐有目的地选择保留木,用人工选优代替自然选择,以培育优质木荷林。
3.2 结论
立地条件的好坏,树种选择的适当与否,直接影响着森林生态修复的效果。根据本研究结果,松材线虫病除治迹地可改培为高质量的木荷林。对松材线虫病除治迹地通过改培进行生态修复时,在下坡位营造木荷纯林和混交林都有着较好的生长表现;对中、上坡位而言,无论是从减缓坡位条件差异对林分的影响来看,还是从促进木荷更好生长来看,都宜选择木荷与枫香、榉树或檫木等混交模式。同时,混交模式还能增加林内物种多样性。但森林生态修复是一个复杂的过程,本研究所调查样地木荷和枫香初植比例为1∶1,未设立不同混交比例的比较研究,是否还有其它混交比例更有利于木荷和枫香的生长,有待后期进一步研究。除了木荷和枫香之外,适合生态修复的树种还有很多,继续开展松材线虫病除治迹地不同立地条件生态修复树种选择及配置模式研究,可为除治迹地生态修复提供更多选择,更好地服务于林业生产。
