沙棘无性系对干旱胁迫的响应研究
2021-08-11胡建忠刘丽颖
胡建忠,刘丽颖
(1.水利部沙棘开发管理中心,北京 100038;2.沈阳农业大学林学院,辽宁 沈阳 110866)
干旱是我国“三北”地区常见的一种自然现象。植物对干旱的适应能力不仅与干旱强度、速度有关,更受其自身基因的调控[1]。在一定干旱阈值胁迫范围内,很多植物能够进行相关抗旱基因的表达,随之产生一系列生理、生化及形态结构等方面的变化,从而显现出抗旱性的综合性状[2]。限于盆栽研究手段的制约,国内目前对作物或牧草等草本植物干旱胁迫的研究[3-4]较多,而对林木开展的较少,且只能以苗期研究[5-6]为主。沙棘属Hippophae植物是我国“三北”地区广为栽培的一类生态经济型树种,深入研究沙棘的抗旱机理,揭示其抗旱特性,科学选育适宜广大干旱半干旱地区种植的优良沙棘无性系品种,已显得十分重要。但就沙棘而言,有代表性的研究论文涉及到对西藏分布的云南沙棘H.rhamnoidesssp.yunnanensis、江孜沙棘H.gyantsensis和肋果沙棘H.neurocarpa3种沙棘属植物干旱胁迫下光合特性研究[7],对引进大果沙棘“太阳”和中国沙棘H.rhamnoidesssp.sinensis优选类型“丰宁”开展的干旱胁迫下分子机制研究[8],以及对干旱胁迫下中国沙棘脯氨酸含量变化[9]的研究。可见沙棘属植物干旱胁迫领域还有许多工作要做,有鉴于此,本文就5个杂交沙棘无性系开展了干旱胁迫响应的有关研究探讨。
1 材料与方法
试验地设在沈阳农业大学校内试验场,5个沙棘无性系为“杂雌优1号”、“杂雌优2号”、“杂雌优10号”、“杂雌优12号”和“杂雌优54号” ,是用不同蒙古沙棘Hippophaerhamnoidesssp.mongolica亚种做母本、中国沙棘做父本杂交选育而成。
采用盆栽试验方法。3月下旬,将当年早春采集的沙棘硬枝插条经过生根粉浸泡4 h后,插在深33 cm、容积约8 L的花盆中,每个无性系30盆,共150盆,每盆扦插2株。所用土壤为培养土与石英砂1∶1(体积比)的混合物,培养土为采自东北针叶林林下的自然土壤。扦插后将花盆随机摆放到温室大棚中培养。
经过4个多月的生长,从8月16日起,对5个参试沙棘无性系进行干旱胁迫处理。设计3个水分处理,分别为正常水分(对照)、中度干旱、重度干旱,其对应的土壤含水率分别为田间持水量的80%、50%和30%。
测定指标:叶面积和干质量,分别采用扫描仪和称重法测定;叶片净光合速率、蒸腾速率、气孔导度、胞间CO2浓度和气孔限制值等光合特性值,采用Li-6400光合仪(美国,Li-Cor公司)测定;长期水分利用效率,即叶片的稳定碳同位素比值(δ13C值)采用稳定同位素比质谱仪MAT-253型(Finigan公司)测定;超氧化物歧化酶(SOD)、抗坏血酸过氧化物酶(APX)、过氧化氢酶(CAT)、过氧化物酶(POD)4类抗氧化酶的活性,采用Chakraborty等[10]和李忠光等[11]的方法测定。所有数据均用SPSS 20.0进行统计分析。
2 结果与分析
2.1 叶性状响应
叶面积、叶干质量等性状是沙棘适应环境所体现在叶片上的特征参数,对环境变化具有重要指标意义。干旱胁迫造成沙棘无性系叶干质量及叶面积的下降,结果见表1。
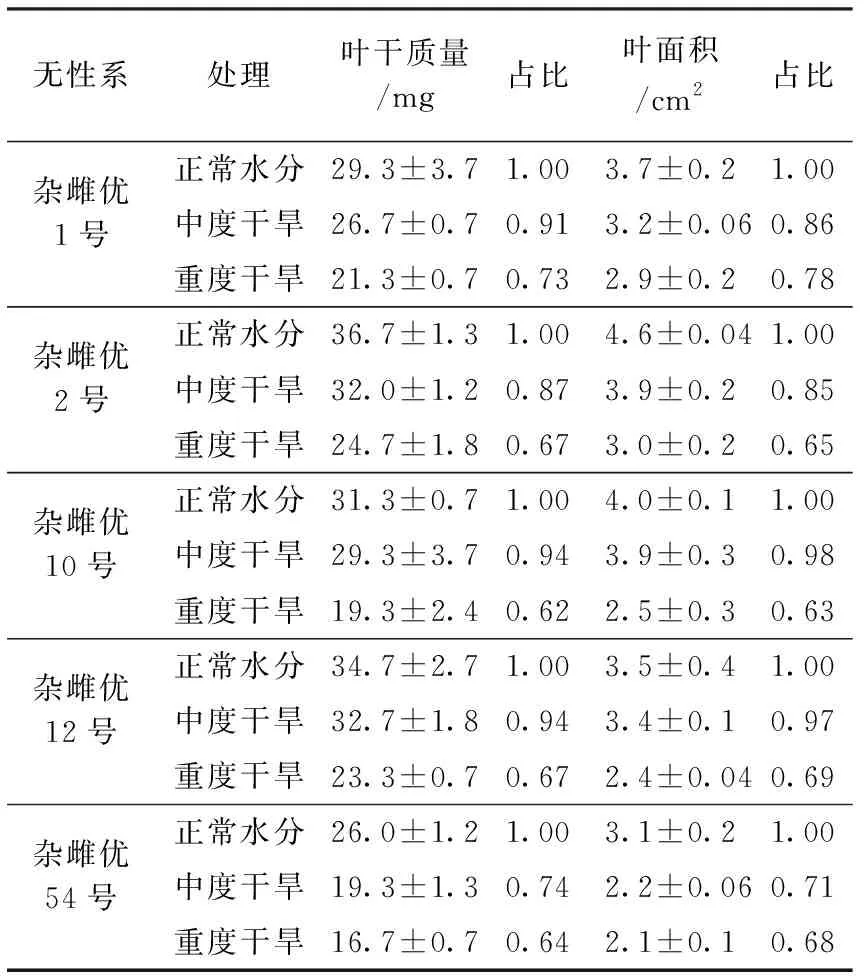
表1 不同干旱胁迫条件下沙棘无性系叶片性状
从表1可以看出,在中度干旱条件下,叶干质量占正常水分条件下的88%(74%~94%);在重度干旱条件下,叶干质量仅占正常水分条件下的67%(62%~73%)。同样,在中度干旱条件下,叶面积占正常水分条件下的87%(71%~98%);在重度干旱条件下,叶面积仅占正常水分条件下的68%(63%~78%)。叶干质量和叶面积在中度、重度条件下的占比较为接近。
2.2 光合特性响应
从表2可以看出,5个沙棘无性系净光合速率平均值,正常水分条件下为20.59 μmol·m-2·s-1,中度干旱条件下为12.32 μmol·m-2·s-1,重度干旱条件下为5.40 μmol·m-2·s-1,中度、重度分别相当于正常水分条件下的60%和26%。可见在干旱胁迫条件下,净光合速率随胁迫程度高低而相应有所下降。
5个沙棘无性系蒸腾速率平均值,在正常水分条件下为4.74 mmol·m-2·s-1,在中度干旱条件下为2.41 mmol·m-2·s-1,在重度干旱条件下为1.02 mmol·m-2·s-1,分别相当于正常水分条件下的51%和21%。在干旱胁迫条件下沙棘无性系部分气孔关闭,降低了蒸腾速率,蒸腾拉力下降,用于光合作用的水分减少,生长量也随之减少。
5个沙棘无性系气孔导度平均值,在正常水分条件下为0.22 mol·m-2·s-1,在中度干旱条件下为0.11 mol·m-2·s-1,在重度干旱条件下为0.04 mol·m-2·s-1,中度、重度分别相当于正常水分条件下的50%和19%。
5个沙棘无性系胞间CO2浓度平均值,在正常水分条件下为212.67 μmol·mol-1,在中度干旱条件下为169.98 μmol·mol-1,在重度干旱条件下为199.20 μmol·mol-1,中度、重度分别相当于正常水分条件下的80%和94%,不降反升,主要原因是此时净光合速率低,对CO2利用较少。
5个沙棘无性系胞间气孔限制值,在正常水分条件下平均值为0.46,在中度干旱条件下平均值为0.56,在重度干旱条件下平均值为0.54。结合不同干旱胁迫下沙棘胞间CO2浓度和气孔限制值的变化,可以判断沙棘在中度干旱胁迫下净光合速率的下降主要由气孔限制引起。而重度干旱胁迫下,参照有关研究[12],主要由非气孔限制引起,如叶绿体结构、叶绿素荧光参数、光合色素和酶活性、活性氧代谢等。
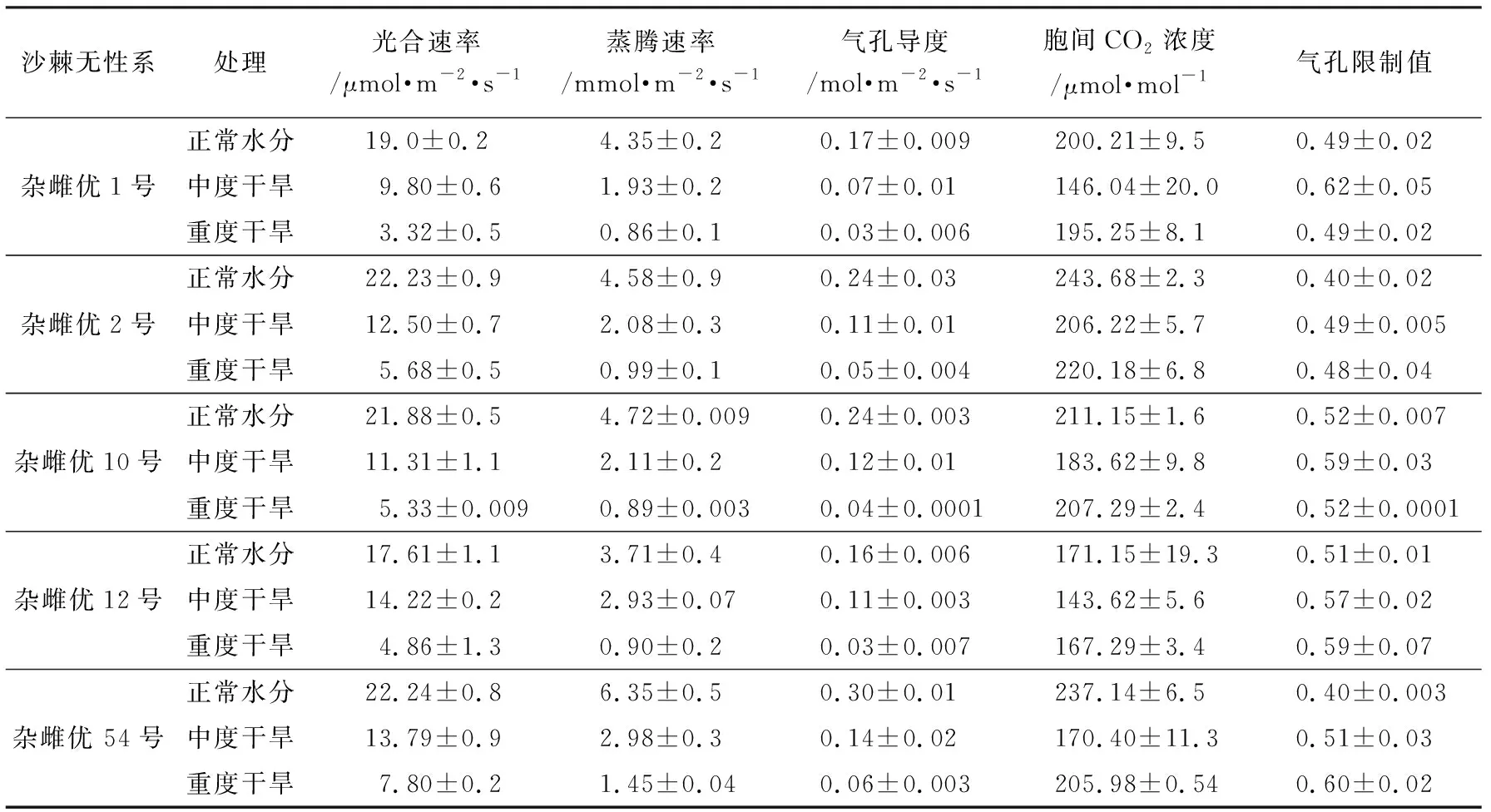
表2 不同干旱胁迫条件下沙棘无性系的光合特征
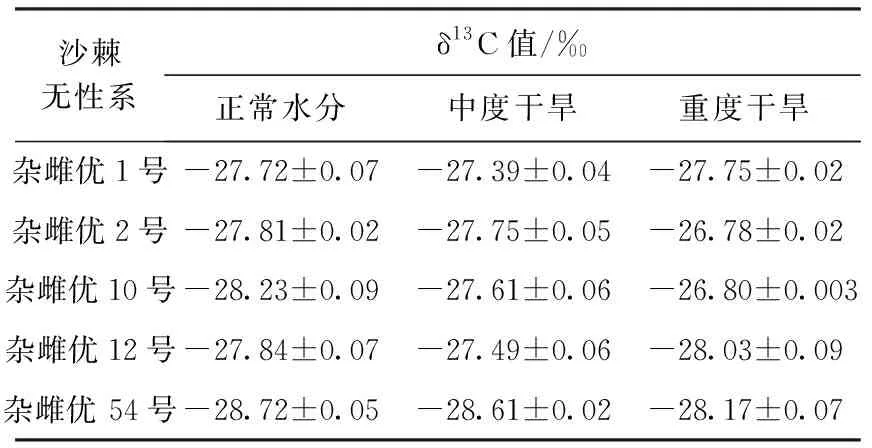
表3 不同干旱胁迫条件下沙棘无性系叶片δ13C值
从表3可以看出,在正常水分条件下,沙棘无性系δ13C值按从大到小的排序为“杂雌优1号”>“杂雌优2号”>“杂雌优12号”>“杂雌优10号”>“杂雌优54号”。5个沙棘无性系在受到中度干旱胁迫时均会提高水分利用效率,表明其有较强的抗旱性。在受到重度干旱胁迫时,“杂雌优1号”和“杂雌优12号”的δ13C值会降低,而 “杂雌优2号”、“杂雌优10号”和“杂雌优54号”的δ13C值还会继续增加,表明这3个沙棘无性系抗旱性能较强。
2.3 酶活性响应
从表4可以看出,5个沙棘无性系在中度干旱胁迫下,均会提高SOD、APX、CAT、POD等抗氧化酶活性,表明在干旱条件下这5个沙棘无性系均具有较敏感的应激反应,能够通过提高抗氧化酶活性以抵抗干旱胁迫对自身带来的损伤。随着干旱程度的加剧,在重度干旱胁迫时,“杂雌优1号”和“杂雌优12号”的抗氧化酶活性较中度干旱胁迫时有所下降(但仍高于正常水分条件下酶活性),这可能是由于沙棘体细胞受到了一定伤害使得抗氧化酶活性降低,或其他原因所致;而“杂雌优2号”、“杂雌优10号”和“杂雌优54号”的抗氧化酶活性仍继续升高,表明这3个沙棘无性系在持续干旱的条件下仍具有较高的抗性。

表4 不同干旱胁迫条件下沙棘无性系叶片的酶活性
3 结论与讨论
5个沙棘无性系叶片干质量在中度干旱条件下,占正常水分条件的88%(74%~94%);在重度干旱条件下,占正常水分条件的67%(62%~73%)。同样,叶面积在中度干旱条件下,占正常水分条件下的87%(71%~98%);在重度干旱条件下,占正常水分条件下的68%(63%~78%)。叶干质量、叶面积在中度、重度干旱条件下占比基本相同。
5个沙棘无性系的净光合速率、蒸腾速率、气孔导度都以正常水分条件下最高,重度干旱条件下最低。在田间持水量80%以下,随着土壤水分含量的减少,这些指标也随之减少。胞间CO2浓度在中度干旱胁迫下会显著降低,而在重度干旱胁迫下反而上升,接近对照处理,并未像净光合速率一样随着干旱加剧而持续降低。这表明在重度干旱胁迫下沙棘光合速率的下降并非完全是由于气孔关闭引起CO2不足所致,应该是非气孔因素在起作用。气孔限制值在中度干旱条件下会增加,重度干旱条件下降低。在重度干旱胁迫时, “杂雌优2号”、“杂雌优10号”和“杂雌优54号”的长期水分利用效率(δ13C值)、抗氧化酶活性仍继续升高,表明这3个沙棘无性系在持续干旱条件下仍具有较强的抗旱性,可用于“三北”地区生态经济型沙棘林建设。
研究干旱条件下沙棘生理生化特征变化,对今后培育抗旱沙棘新品种具有非常重要的理论和实践意义。干旱胁迫下导致沙棘无性系叶面积和干质量下降,主要是通过降低气孔导度和抑制叶绿体光合活性造成的。沙棘对干旱信号的最先反应是调整气孔开度,防止水分散失,本文的研究结果也证明了这一点。而在重度干旱条件下,是否会使沙棘生物膜结构与功能遭到破坏,随之叶绿素、核酸和蛋白质等大分子遭到破坏和损伤,进而引起一系列生理生化功能的改变,最终导致光合器官活性下降[13],本文没有涉及,将是需要后续开展研究的重要内容。
