牛乳、驴乳乳清蛋白二级结构及其功能对比研究
2021-08-09刘爱成李墨翰张正翰张秀敏刘一鸣陈佳丽热罕古丽岳喜庆
刘爱成,李墨翰,张正翰,张秀敏,刘一鸣,陈佳丽,热罕古丽,郑 艳,岳喜庆,*
(1.沈阳农业大学食品学院,辽宁 沈阳 110866;2.北京食品科学研究院,北京 100068;3.沈阳农业大学外语教学部,辽宁 沈阳 110866)
驴乳是一种特种乳,其基本组成与人乳较为相近。驴乳中乳清蛋白相对含量较高,VC含量丰富,钙磷含量超过人乳[1];驴乳脂肪含量较低,脂肪酸组成丰富,营养价值高,亚麻酸和亚油酸含量高于牛乳和羊乳[2];驴乳中活性成分较多,如溶菌酶和乳铁蛋白等,具有增强免疫力、消除炎症等作用,适合多类人群食用及多种配方食品的生产,具有较大的开发潜力[3-5]。牛乳营养价值丰富,受众范围广,占据了乳类消费市场的绝大多数份额[6]。牛乳中乳糖、蛋白质和脂肪含量较高,可以为人体提供能量[7],牛乳中的α-乳白蛋白能为人体提供重要的必需氨基酸,对婴幼儿的成长发育极为重要[8]。目前婴幼儿配方乳粉多用牛乳作为基料,但是由于一部分婴幼儿对牛乳蛋白过敏,会产生一些不良反应[9],所以驴乳受到人们的关注,并逐渐被用于婴幼儿配方食品的生产。在国外,驴乳被用于治疗牛乳蛋白过敏症状,并且取得了较好的疗效,因此驴乳也被称为“低致敏性”乳[10]。
新生儿的能量补充来自蛋白质、碳水化合物和脂质等,蛋白质能够补充营养及发挥功能作用,促进生长发育[11]。乳清蛋白易于消化吸收,营养价值高,适合作为婴幼儿配方食品基料,如通过添加乳清蛋白调整蛋白比例,使婴幼儿配方乳粉的组成更接近母乳[12-13]。α-螺旋和β-折叠是蛋白质中常见的二级结构[14],每种结构都具有一定的功能,其中α-螺旋结构十分稳定,能够作为识别基序,介导蛋白质间相互作用[15]。傅里叶变换红外光谱法、核磁共振技术、圆二色光谱法等是目前较为常用的测定蛋白质二级结构的技术[16]。在上述方法中,傅里叶变换红外光谱法因其适用性强,能在多种环境下对蛋白质等生物大分子进行定量分析从而被广泛使用。近年来,国内外对牛乳和驴乳组成成分和营养价值研究较多,但对于牛乳和驴乳蛋白质二级结构及其蛋白质功能的对比分析较少。
本研究首先通过傅里叶变换红外光谱分析驴初乳、驴常乳、牛初乳、牛常乳乳清蛋白的二级结构,通过OMNIC 8.2和Peakfit 4.12软件分析谱图,随后利用液相色谱-质谱联用技术对上述4 种乳样的乳清蛋白进行鉴定,并进一步利用基因本体论(gene ontology,GO)功能注释和KEGG(Kyoto Encyclopedia of Genes and Genomes)代谢通路分析其功能及差异。比较上述4 种乳样中乳清蛋白二级结构及功能性质的差异,将有助于加深对不同乳源及不同泌乳期乳清蛋白成分的认识,为开发更适合婴幼儿需求的配方乳粉提供一定的参考。
1 材料与方法
1.1 材料与试剂
牛初乳、牛常乳采自沈阳沈北新区伊利牧场,驴初乳、驴常乳采自大连圣鸿道荙驴业科技发展有限公司养殖场,其中初乳采集时间均为分娩后0~7 d,常乳均为分娩后30~60 d。上述4 种乳样各采集30 份,于超低温冰箱(-80 ℃)贮存待用,在实验前解冻并将同种乳样充分混合,以排除个体差异[17]。
1.2 仪器与设备
FTIR-650傅里叶变换红外光谱仪 苏州贝锐仪器科技有限公司;5804R多功能台式离心机 德国艾本德股份公司;Q-Exactive质谱仪 赛默飞世尔科技有限公司;UniStar 2真空冷冻干燥机 北京天利联合有限公司;1290 Infinity Ⅱ液相色谱仪、强阳离子交换色谱(SCX)柱 美国安捷伦科技公司。
1.3 方法
1.3.1 乳清蛋白二级结构测定
1.3.1.1 样品前处理
每组称取30 mL乳样进行离心(4 ℃、10 000 r/min、10 min),除去上层脂质;随后将样品再次离心(4 ℃、12 000 r/min、60 min),使用低温丙酮法对上清液进行处理,除去酪蛋白,再将上清液冻干,制成乳清蛋白冻干粉[18]。
1.3.1.2 傅里叶变换红外光谱图分析
傅里叶变换红外光谱检测前将冻干样品通过溴化钾压片法制样,制样后使用衰减全反射全范围扫描。通过OMNIC 8.2软件对样品谱图的酰胺Ⅲ带(1 220~1 330 cm-1)和酰胺Ⅰ带(1 600~1 700 cm-1)进行处理,再使用Peakfit 4.12软件对所得的曲线进行最终的处理分析,确定各子峰对应的二级结构,根据峰面积求出相对含量。
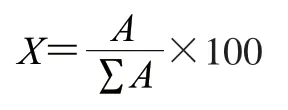
式中:X为二级结构相对含量/%;A为各二级结构的峰面积;ΣA为各二级结构峰面积总和。
1.3.2 蛋白质功能测定
参考张熙桐等[19]的处理方法,提取乳清蛋白后,使用过滤辅助蛋白质组制备法进行酶解,随后采用毛细管高效液相色谱与电喷雾质谱法对蛋白进行鉴定。再利用GO功能注释及KEGG代谢通路对乳清蛋白的生物学功能及参与的代谢通路进行分析。
1.4 数据处理
所有实验均至少重复3 次,数据以平均值±标准差的形式表示,选用SPSS(version 22.0)统计分析软件及Design-Expert 8.0.6软件进行数据处理。
2 结果与分析
2.1 乳清蛋白二级结构分析

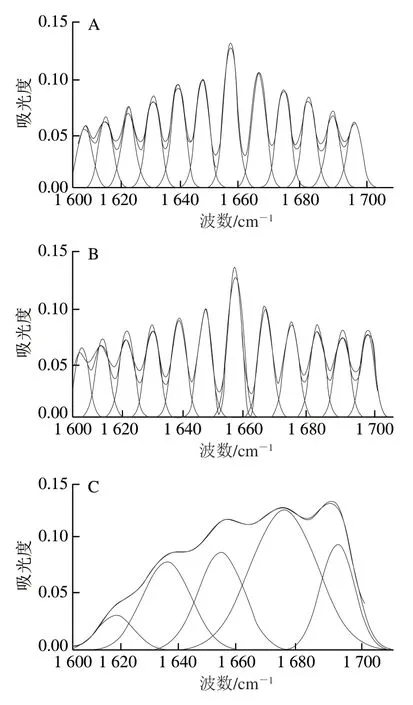
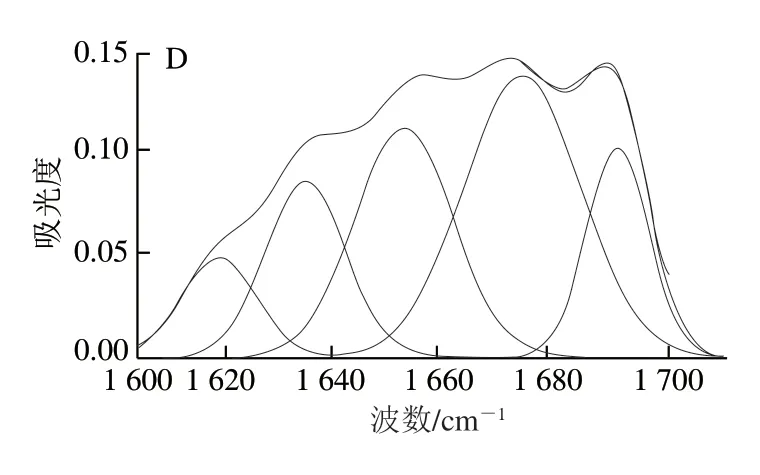
图2 驴初乳(A)、驴常乳(B)、牛初乳(C)和牛常乳(D)乳清蛋白酰胺Ⅰ带拟合效果Fig. 2 Amide I band curve-fitting results for whey proteins in donkey colostrum (A), donkey milk (B), bovine colostrum (C) and bovine milk (D)
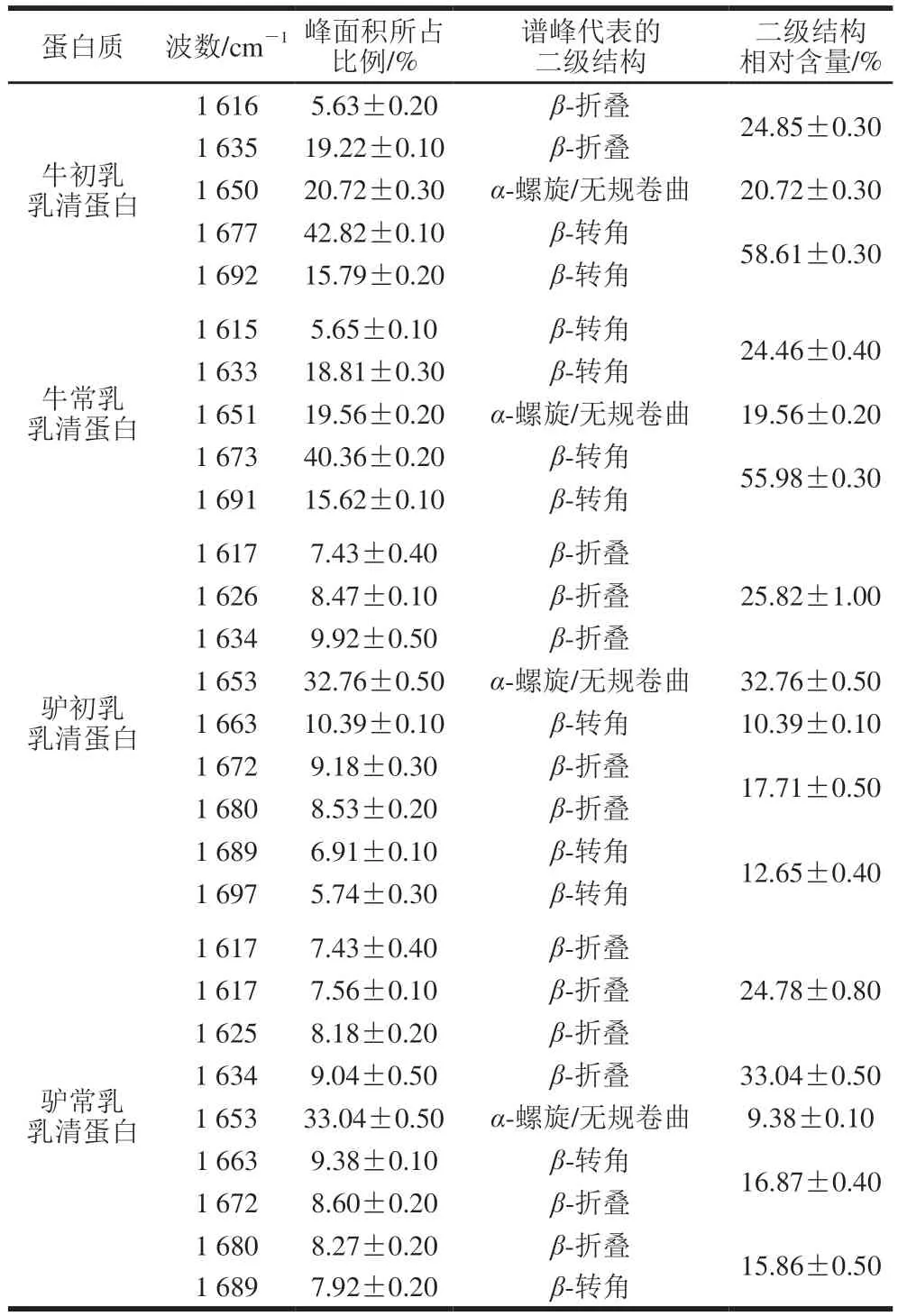
表1 驴初乳、驴常乳、牛初乳和牛常乳乳清蛋白酰胺Ⅰ带分析结果Table 1 Secondary structure contents of whey proteins in donkey colostrum, donkey milk, bovine colostrum and bovine milk determined from amide I band
由图1可知,4 种乳样光谱图的变化规律大致相似,将4 种乳样原谱图通过OMNIC 8.2软件和Peakfit 4.12软件进行处理(图2),对酰胺Ⅰ带进行分析,得出每种乳样的二级结构及对应的相对含量(表1),为更好区分某些二级结构基序及更准确定量,对酰胺Ⅲ带进行分析[20](表2)。分析结果显示,驴乳和牛乳乳清蛋白二级结构存在差异。驴乳乳清蛋白中α-螺旋及β-折叠相对含量均高于同期牛乳,而β-转角相对含量则低于牛乳。与牛初乳相比,牛常乳乳清蛋白中的二级结构相对含量均降低;与驴初乳相比,驴常乳中β-折叠结构相对含量降低而α-螺旋结构相对含量升高。驴乳中β-折叠结构相对含量较高,研究发现,乳中含有的缬氨酸和异亮氨酸能促进β-折叠结构的形成,同时,驴乳中所含的脯氨酸也有利于β-转角结构的形成[21]。而β-转角结构过多会导致α-螺旋结构相对含量降低,间接影响人体吸收利用,驴常乳乳清蛋白中α-螺旋结构相对含量最高,β-转角结构相对含量最低,因此最适于人体吸收。
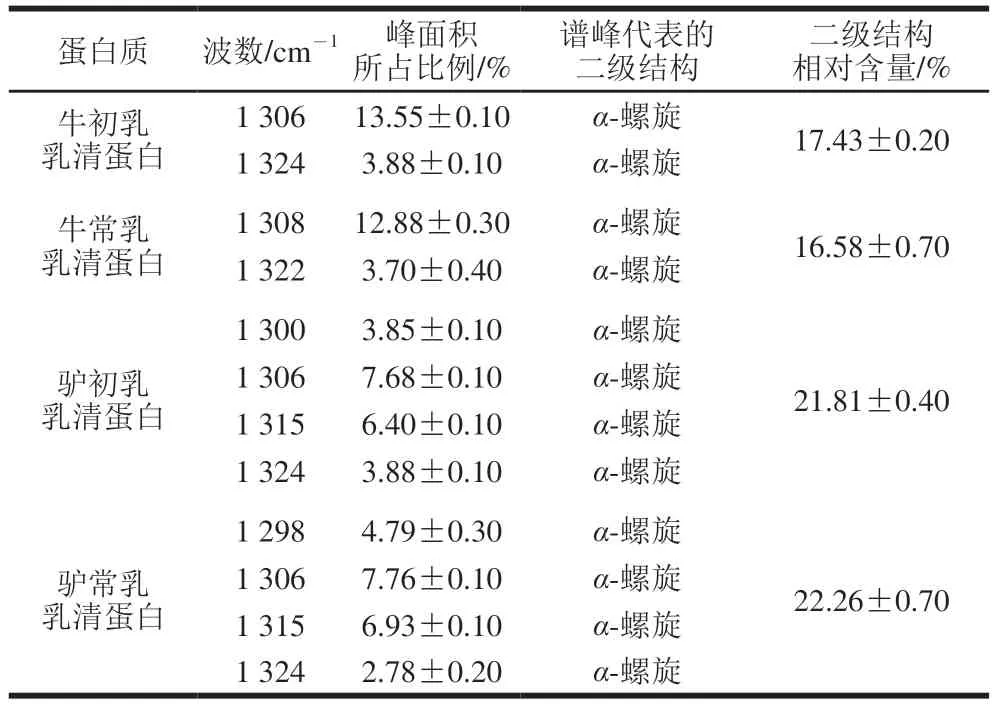
表2 驴初乳、驴常乳、牛初乳和牛常乳乳清蛋白酰胺Ⅲ带分析结果Table 2 Secondary structure contents of whey proteins in donkey colostrum, donkey milk, bovine colostrum and bovine milk determined from amide III band
蛋白质变性后,二级结构中的α-螺旋会发生转变,从而导致β-折叠和β-转角结构含量增加[22]。由于驴乳和牛乳在不同泌乳期的成分存在一定的差异,如驴乳中的蛋白质含量在泌乳期有先降低后稳定上升的趋势[23],因此也可能导致驴乳乳清蛋白二级结构的变化。
2.2 乳清蛋白GO功能注释分析
2.2.1 参与的生物过程分析

图3 4 种乳乳清蛋白参与的生物过程Fig. 3 Biological processes involving whey proteins in four milks
由图3可知,4 种乳样乳清蛋白参与的主要生物过程包括代谢过程、对刺激的反应、定位及免疫系统过程等,其中参与代谢过程的乳清蛋白种类数最多,其次为参与对刺激的反应过程的乳清蛋白种类数较多。在上述4 种乳样的乳清蛋白中,驴常乳中参与生物过程的乳清蛋白种类数最多,其次是驴初乳,牛常乳与牛初乳差别不大。整体来看,牛乳中参与生物过程的乳清蛋白种类数均低于驴乳,随着泌乳期的延长,驴乳和牛乳参与生物过程的乳清蛋白种类数均增加,但牛乳增加幅度不大。因此,相较于牛乳,驴乳在生物功能方面更具优势。由于婴幼儿的免疫力较低,相较于牛乳,驴乳能更好地增强免疫力,所以配方乳粉可以使用驴乳为基料进行研发,刺激婴幼儿肠道菌群生成,增强肠道功能,促进免疫系统发育[24-25]。良好的应激反应也有利于婴幼儿适应外界环境,维持机体稳定。驴常乳在各过程中参与的乳清蛋白种类数均最多,发挥的作用最明显,说明驴常乳相较于牛乳及驴初乳在生物功能方面更具优势。
2.2.2 分子功能分析
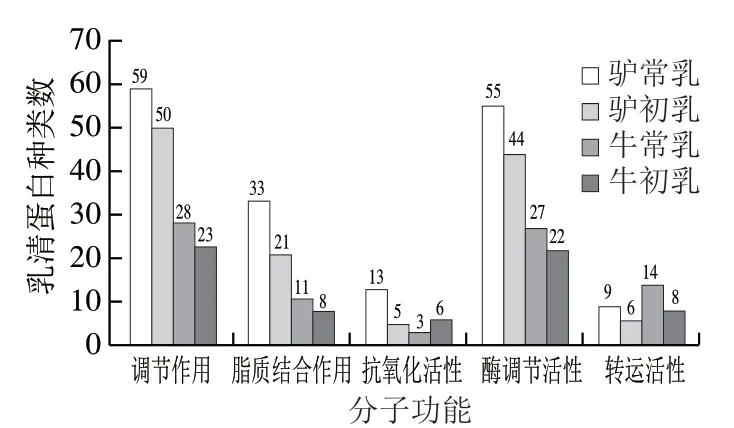
图4 4 种乳乳清蛋白参与的分子功能Fig. 4 Molecular functions involving whey proteins in four milks
由图4可知,调节作用是4 种乳样的乳清蛋白参与最多的分子功能,其次是酶调节活性、脂质结合作用、抗氧化活性和转运活性等。整体而言,4 种乳中参与分子功能的乳清蛋白种类数存在一定的差距,除转运活性外,牛乳中参与分子功能的乳清蛋白种类数均少于驴乳;随着泌乳期的延长,除抗氧化活性外,牛乳和驴乳中参与分子功能的乳清蛋白种类数均增加;驴乳中参与调节作用和酶调节活性的乳清蛋白种类数较多,且高于牛乳,目前已有研究表明,驴乳能够调节胰岛B细胞活力及与糖代谢有关酶的活性,对治疗2型糖尿病具有重要作用[26]。苏德奇等[27]的研究也证明了驴乳的抗氧化功效,能够对机体损伤部位产生一定的保护作用。驴常乳除参与转运活性的乳清蛋白种类数低于牛常乳,参与其余分子功能的乳清蛋白种类数均最多,相较于牛乳和驴初乳,驴常乳在分子功能上发挥的作用更明显。
2.2.3 细胞组成分析
由图5可知,驴乳与牛乳乳清蛋白主要参与的细胞组成有胞外区部分、细胞器、胞外体、胞囊等,驴乳和牛乳中参与胞外区的乳清蛋白种类数最多。在血液微粒组成上牛乳和驴乳参与的乳清蛋白种类数差距较小。整体而言,驴常乳乳清蛋白参与细胞组成的乳清蛋白种类数最多,驴初乳次之,牛初乳最少。随着泌乳期的延长,驴乳和牛乳乳清蛋白参与细胞组成的乳清蛋白种类数逐渐增多,驴乳增加幅度较大,牛乳增加幅度较小。胞外区和细胞器区是细胞组成中2 个主要区域,很多细胞激活反应及分子结合均发生在胞外区。膜性细胞器是一种相对局域化的空间结构,由膜包裹,在机体内有序行使细胞分裂、增殖、呼吸作用等生物学功能[28]。牛乳乳清蛋白参与细胞组成的乳清蛋白种类数与驴乳乳清蛋白存在明显差距,由此可见,驴常乳更有助于促进婴幼儿机体内的细胞组成,且很大程度上优于牛乳。
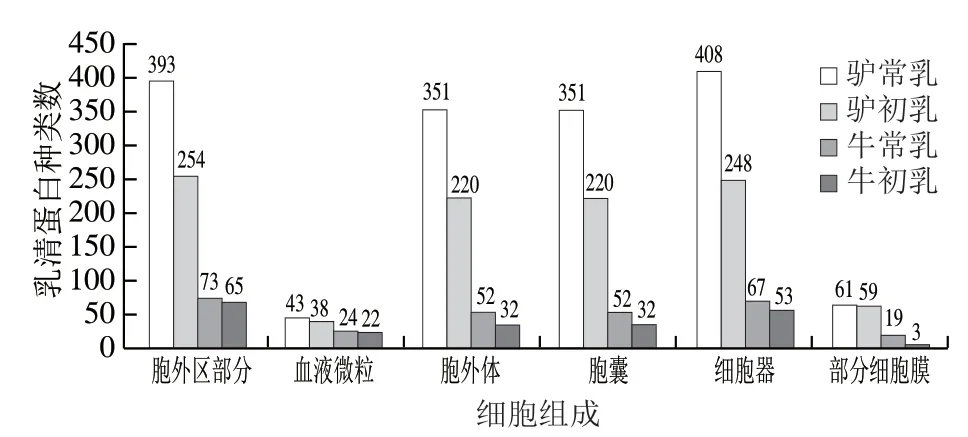
图5 4 种乳乳清蛋白参与的细胞组成Fig. 5 Cellular components involving whey proteins in four milks
2.3 4 种乳乳清蛋白参与的KEGG通路分析

表3 4 种乳乳清蛋白参与KEGG通路种类数Table 3 Number of KEGG pathways involving whey proteins in four milks
由表3可知,通过KEGG通路分析得出,驴乳乳清蛋白参与的KEGG通路数量多于牛乳,驴初乳、驴常乳乳清蛋白分别参与17、28 条KEGG代谢通路,而牛初乳、牛常乳则分别参与3、10 条KEGG代谢通路。随着泌乳时间的延长,牛乳和驴乳乳清蛋白参与的KEGG通路数量均增加,4 种乳存在3 条共同的KEGG通路,且都与代谢、免疫和疾病相关,关乎婴幼儿的成长发育。以系统性红斑狼疮为例,儿童系统性红斑狼疮的发病机制与多因素有关,包括遗传、环境和免疫系统失调等,导致机体组织器官出现炎症和损伤[29]。牛乳和驴乳中存在的吞噬通路、补体和凝血级联反应通路,能够调节免疫,起到一定的预防作用。
吞噬及补体和凝血级联反应是驴常乳乳清蛋白主要参与的功能,乳清蛋白能够增强巨噬细胞的吞噬能力,增强人体免疫功能[30]。补体系统和凝血系统相互关联,维持体内平衡,补体C3为补体成分之一,能够激活凝血功能[31]。从整体来看,驴常乳乳清蛋白参与的KEGG通路最为丰富,且参与内质网蛋白质加工、溶酶体、抗生素生物合成等,对于人体蛋白质的新陈代谢及免疫功能等方面具有重要价值。综上可知,驴常乳相较于驴初乳、牛初乳和牛常乳更适合作为婴幼儿功能性食品原料。
3 结 论
目前,通过傅里叶变换红外光谱法对比乳清蛋白二级结构的研究较少。本研究通过对驴常乳、驴初乳、牛常乳和牛初乳乳清蛋白的结构和功能性质进行分析,对比它们的二级结构和功能差异性,为进一步利用不同泌乳期的牛乳与驴乳改良现有乳粉、开发更适用于婴幼儿体质的新型配方乳粉提供了一定的数据参考。通过傅里叶变换红外光谱对蛋白质二级结构进行分析可知,驴乳和牛乳乳清蛋白二级结构存在一定的差异,驴乳乳清蛋白中常见的二级结构相对含量高于牛乳,驴初乳与驴常乳乳清蛋白中α-螺旋结构相对含量相差不大,牛常乳中α-螺旋结构相对含量最低。与人乳中常见的二级结构进行对比,驴常乳整体与人乳最为接近。通过GO功能注释和KEGG通路分析发现,驴乳和牛乳在生物学功能和参与的代谢通路方面也存在一定的差异。驴乳乳清蛋白参与的生物学功能和代谢通路的数量均高于牛乳,且驴常乳和牛常乳参与的KEGG通路种类数均多于初乳。整体而言,随着泌乳期的延长,4 种乳中参与生物学功能和代谢通路的乳清蛋白种类数均增多,其中驴常乳乳清蛋白发挥的功能作用最明显,更有益于婴幼儿配方乳粉的改良。
