番茄嫁接植株根系内生细菌和真菌群落的组成特征
2021-08-05覃仁柳庞师婵唐小付黄子粤杨尚东
覃仁柳,庞师婵,唐小付,黄子粤,肖 健,杨尚东
(广西大学农学院/植物科学国家级实验教学示范中心,广西 南宁 530004)
【研究意义】番茄是广西秋冬蔬菜生产的主要品种之一,也是广西“南菜北运”和“西菜东调”的重要蔬菜种类。我国番茄连作现象较普遍,土传病害和连作障碍十分严重,阻碍了番茄产业的可持续发展[1]。嫁接是降低土传病害、克服连作障碍的一项有效技术措施,嫁接技术已在番茄[2]、辣椒[3]、茄子[4]、黄瓜[5]等瓜菜类作物生产上得到较广泛应用。已有研究分别从组织结构[6]、生理生化[7]和抗性诱导[8]等方面阐述了嫁接可增强番茄植株抗性的机制,并发现植物病害的发生不仅与根际微生物群落组成和功能存在联系,与植株根系内生微生物也存在显著相关性。因此,研究嫁接对番茄植株根系内生微生物多样性及群落结构特征的影响,对解析嫁接番茄根系内生微生物多样性与抗性间的联系具有重要意义。【前人研究进展】番茄嫁接植株根际土壤中更丰富的细菌多样性和更多样的特有优势细菌属是嫁接植株抗性增强的重要原因之一[9]。植物内生微生物是微生物研究的重要领域,微生物结构组成和数量与植物养分循环、病虫害发生及植物健康紧密相关[10]。植物根系的内生细菌主要入侵和定殖在植物根区,并利用植物的代谢产物生存和生长,促进宿主植物生长发育,增加总生物量,提升植物抗性、产生抗生素类物质[11],同时增加植物的固氮能力和对钾、钙和磷等营养元素的吸收[12]。植物根系内生真菌通过产生生长素、赤霉素和细胞激动素等植物生长素促进宿主植物生长[13],有益内生真菌可发挥抗虫作用,其中枝顶孢属(Acrmonium)菌类可控制蚜虫危害[14]。内生真菌作为杀菌剂对脉孢菌属(Neurosporasp.)、镰刀菌(Fusariumsp.)、木霉(Trichodermasp.)等病原真菌具有生长抑制活性[15]。【本研究切入点】目前,针对嫁接提高番茄植株抗性的研究多数是从生理生化和抗性诱导等方面进行分析,关于嫁接对番茄植株根系内生微生物群落结构影响的研究鲜见报道。【拟解决的关键问题】基于Illumina MiSeq高通量测序技术分析番茄嫁接植株根系内生微生物多样性及群落结构特征,旨在更深入地探究嫁接番茄植株的抗性机制,为番茄生产上筛选和开发利用有益微生物提供理论依据与技术支撑。
1 材料与方法
1.1 试验材料
砧木分别采用番茄普通品种新组合1号和宝砧1号,接穗为桃太郎。种子均购自农友种苗(中国)有限公司。
1.2 试验方法
1.2.1 试验设计 试验于2019年9月10日至12月30日在广西大学农学院蔬菜生产基地(东经108°17′25″,北纬22°51′02″)温室进行。设5个处理,分别为:新组合1号番茄砧木自根植株(简称新砧处理);以桃太郎为接穗、新组合1号为砧木的嫁接植株(简称桃太郎/新砧处理);宝砧1号番茄砧木自根植株(简称宝砧处理);以桃太郎为接穗、宝砧1号为砧木的嫁接植株(简称桃太郎/宝砧处理);桃太郎番茄接穗自根植株(简称桃太郎处理)。每处理种植60株,3个重复。当砧木长至3叶1心时采用插接法[16]嫁接,植株成活后进行盆栽定植(盆直径30 cm,高40 cm)。田间管理按常规方法进行。
1.2.2 样品采集 定植80 d后,收集根系样品:采用“S”采样方法,3株混样,每处理3次重复;将盆栽翻转,倒出整个植株用蒸馏水缓慢冲洗根系,并对根表面进行消毒,收集根系装入无菌袋,用于植株根系内生细菌和真菌多样性[ACE指数、Chao1指数、Shannon指数、Simpson指数、Heip指数、覆盖度(Coverage)、群落组成和Venn图]检测。
1.2.3 总DNA提取、PCR扩增和序列测定 根系样品总DNA提取、PCR扩增和序列测定由上海美吉生物医药科技有限公司完成。测序具体类型与引物序列见表1。
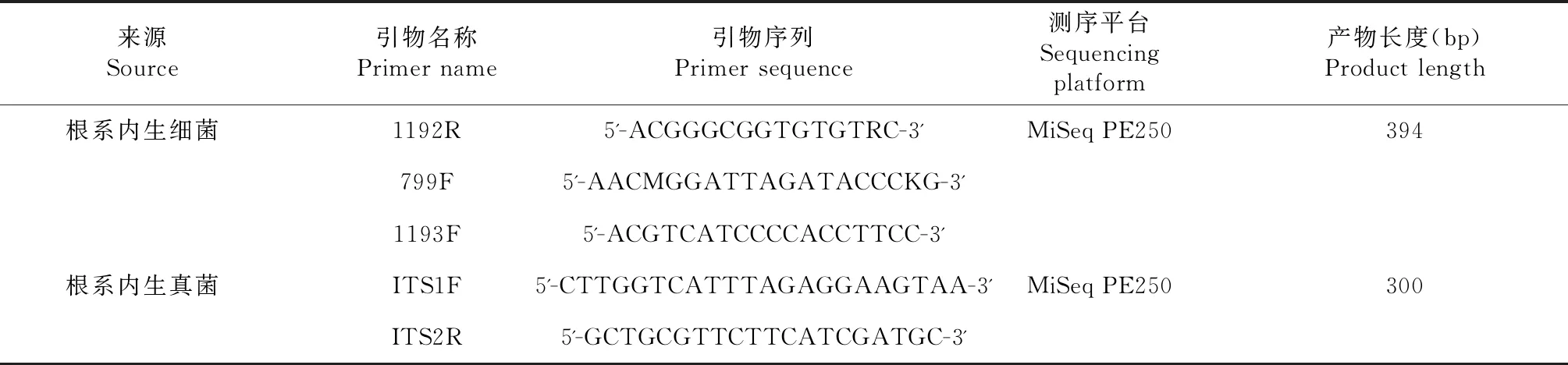
表1 用于PCR扩增的引物序列
1.3 统计分析
采用Excel 2010和SPSS 19.0对试验数据进行统计分析,高通量检测数据通过Usearch(vsesion 7.0)及I-sanger平台进行分析,并构建微生物群落组成图。
2 结果与分析
2.1 番茄嫁接和自根植株样本的测序序列及分类
由表2可知,桃太郎、新砧、宝砧、桃太郎/新砧和桃太郎/宝砧处理的根系内生细菌数分别为591、841、639、526和601 OTUs;根系内生真菌数分别为276、293、263、186和306 OTUs;各处理根系的内生细菌数和真菌数在门、纲、目和科分类水平上无明显差异,而在属和种分类水平上,桃太郎/新砧和桃太郎/宝砧处理根系的内生细菌数明显少于其相应的新砧、宝砧和桃太郎处理根系。可见,番茄嫁接植株根系的内生细菌OTU数明显高于内生细菌OTU数,说明番茄嫁接植株根系的内生细菌OTU数受砧木种类影响较明显。

表2 各品种番茄及其嫁接植株样品的测序序列统计及分类结果
2.2 番茄嫁接和自根植株根系内生细菌和真菌的Alpha多样性特征
由表3可知,番茄各处理植株根系的Coverage均在98.00 %以上,表明测序结果可代表样本中微生物的真实情况;桃太郎/新砧处理和桃太郎/宝砧处理根系内生细菌的ACE指数分别为393.53和473.20,Chao1指数分别为478.54和451.62,均明显小于相应的新砧、宝砧和和桃太郎处理,说明嫁接番茄植株根系的内生细菌丰富度低于其相应的砧木和接穗实生自根根系;桃太郎/新砧、桃太郎/宝砧、新砧、宝砧和桃太郎处理根系内生真菌的Shannon指数分别为1.34、1.80、1.93、1.77和2.46,相互间无显著差异(P>0.05,下同),桃太郎/新砧和桃太郎/宝砧处理根系内生细菌的Shannon指数与其相应新砧和宝砧处理的Shannon指数间也无显著差异,但桃太郎/新砧和桃太郎/宝砧处理根系内生细菌的Shannon指数较大,分别为3.58和3.55,均显著大于桃太郎处理(P<0.05,下同);各处理根系内生真菌的Simpson指数间也无显著差异,但内生细菌的Simpson指数以桃太郎处理最大,且显著大于桃太郎/新砧、桃太郎/宝砧、新砧和宝砧处理,而桃太郎/新砧和桃太郎/宝砧处理内生细菌的Simpson指数均小于新砧和宝砧处理,但相互间差异不显著。说明嫁接可改变番茄植株根系的内生微生物多样性特征,尤其可提高根系内生细菌的多样性。
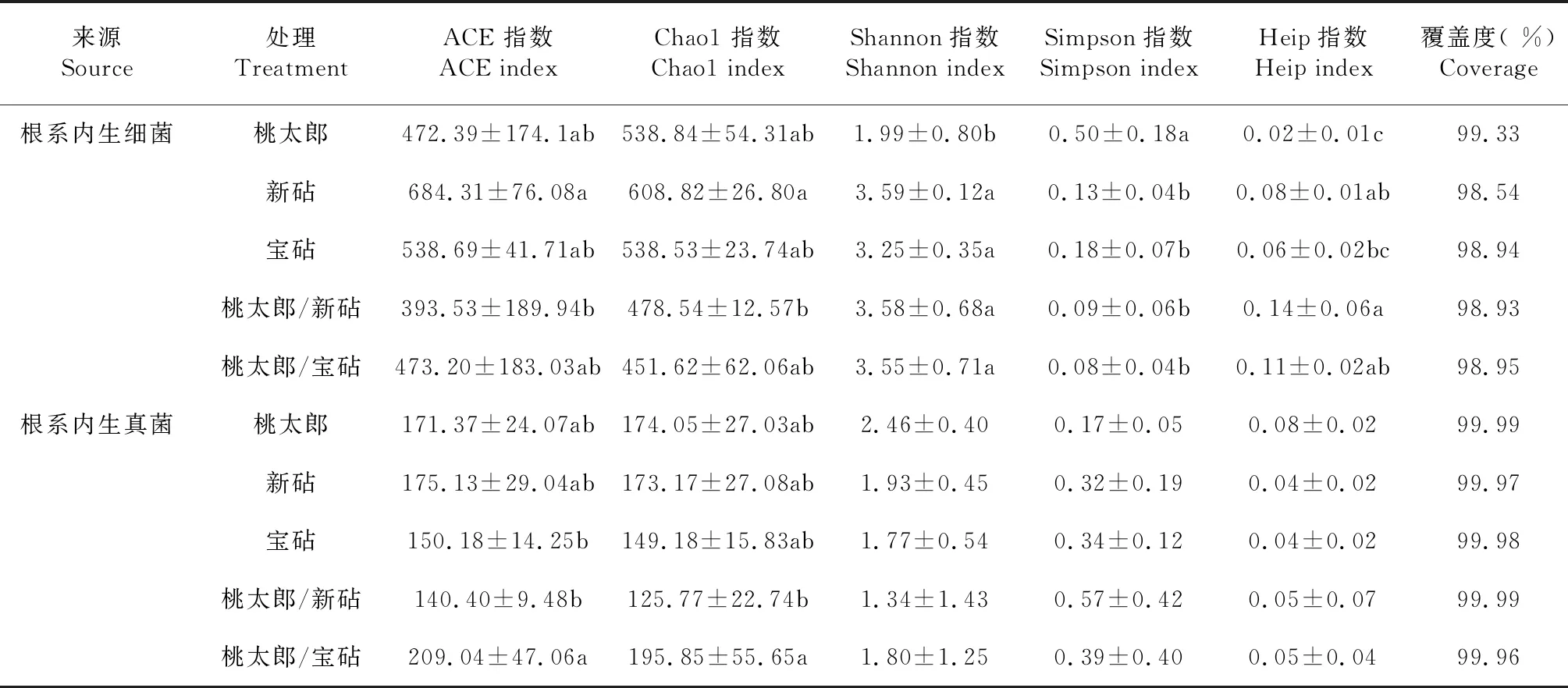
表3 各品种番茄及其嫁接植株根系内生细菌和真菌的多样性指数比较
从表3还可看出,桃太郎、新砧、宝砧、桃太郎/新砧和桃太郎/宝砧处理根系内生真菌的Heip指数分别为0.08、0.04、0.04、0.05和0.05,处理间无显著差异,而桃太郎/新砧和桃太郎/宝砧处理根系内生细菌的Heip指数分别为0.14和0.11,均显著大于桃太郎处理,明显大于相应的新砧和宝砧处理,说明番茄嫁接植株的均匀度较高,即嫁接也有助于改善番茄植株根系内生细菌的群落结构。
2.3 番茄嫁接和自根植株根系内生细菌和真菌的群落组成
2.3.1 门分类水平的优势菌群分析 对比图1各处理番茄植株根系内生细菌门分类水平群落的相对丰度可知,变形菌门(Proteobacteria)、放线菌门(Actinobacteria)、拟杆菌门(Bacteroidetes)和厚壁菌门(Firmicutes)是各品种番茄及其嫁接植株根系内生细菌的共有优势菌门。由表4可知,桃太郎/新砧和桃太郎/宝砧处理根系的变形菌门在其内生细菌组成结构中占比分别为61.23 %和71.21 %,厚壁菌门在其内生细菌组成结构中占比分别为0.74 %和0.40 %,均低于其相应新砧、宝砧和桃太郎处理根系变形菌门和厚壁菌门在其内生细菌组成结构中的占比;而桃太郎/新砧和桃太郎/宝砧处理根系的放线菌门在其内生细菌组成结构中占比分别为20.74 %和18.36 %,拟杆菌门在其内生细菌组成结构中的占比分别为16.60 %和8.71 %,均高于其相应新砧、宝砧和桃太郎处理根系放线菌门和拟杆菌门在其内生细菌组成结构中的占比。可见,嫁接具有改变番茄植株根系内生细菌组成结构占比的作用,且砧木种类不同,其嫁接植株根系内生细菌的组成占比存在差异。
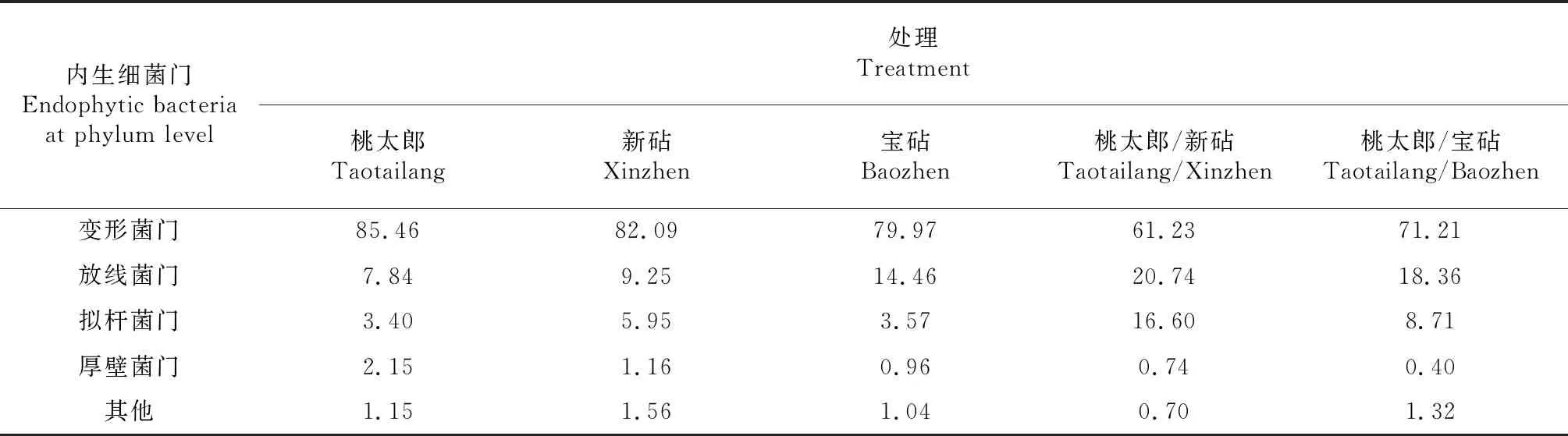
表4 各品种番茄及其嫁接植株根系内生细菌在门分类水平的占比比较
对比图2各处理番茄植株根系内生真菌门分类水平群落的相对丰度可知,子囊菌门(Ascomycota)、油壶菌门(Olpidiomycota)、担子菌门(Basidiomycota)和unclassified_k__Fungi是各处理番茄植株根系内生真菌的共有优势菌门。由表5可知,桃太郎/新砧处理根系的子囊菌门在其内生真菌组成结构中占比为46.73 %,较桃太郎处理低6.57个百分点;担子菌门在其内生真菌组成结构中的占比为41.48 %,较新砧处理高34.42个百分点;而油壶菌门在其内生真菌组成结构中占比为3.65 %,较新砧处理低43.50个百分点,较桃太郎处理低3.49个百分点。从表5还可看出,桃太郎/宝砧处理根系的子囊菌门在其内生真菌组成结构中占比为41.33 %,分别较宝砧和桃太郎处理低7.23和11.97个百分点;担子菌门在其内生真菌组成结构中占比为18.02 %,较宝砧处理高8.21个百分点,较桃太郎处理低19.63个百分点;而油壶菌门在其内生真菌组成结构中占比为36.03 %,较桃太郎处理高28.89个百分点,较宝砧处理低11.12个百分点。可见,采用不同砧木嫁接成活的番茄植株,其根系优势内生真菌组成的占比受砧木种类和接穗共同影响。由此判断,砧穗组合不同是番茄嫁接植株抗性存在差异的重要原因。
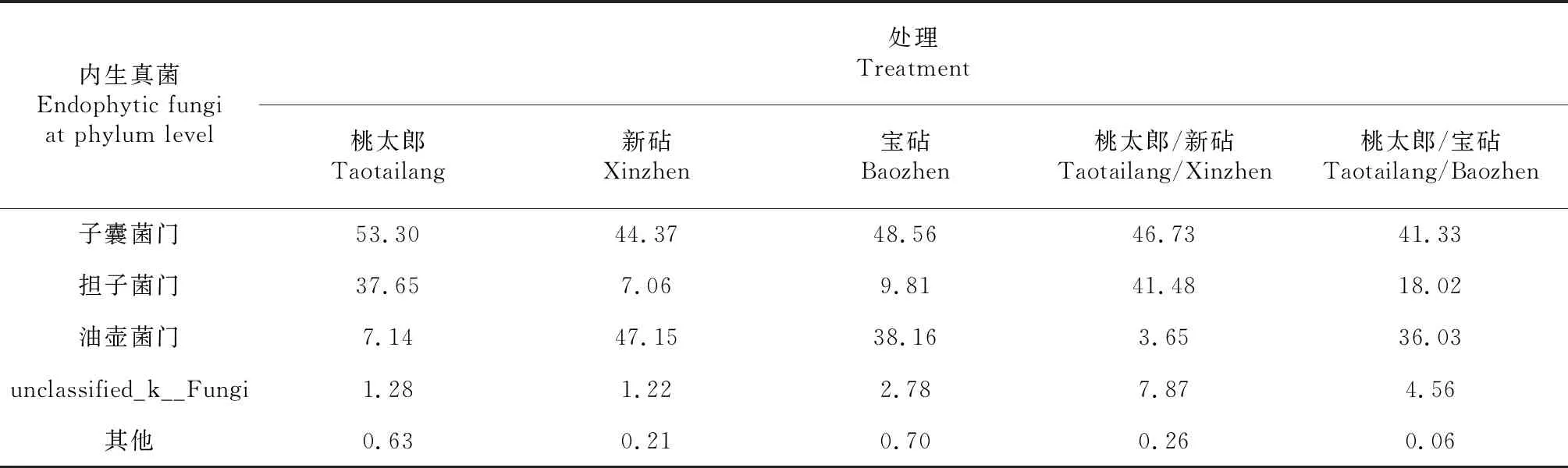
表5 各品种番茄及其嫁接植株根系内生真菌门分类水平的占比比较
2.3.2 属分类水平的优势菌群分析 对比图3各处理番茄植株根系内生细菌属分类水平群落的相对丰度可知,有命名信息的番茄各处理根系内生细菌优势菌属有假单胞菌属(Pseudomonas)、黄杆菌属(Flavobacterium)、波氏杆菌属(Bordetella)、不粘柄菌属(Asticcacaulis)、分支杆菌属(Mycobacterium)、寡养单胞菌属(Stenotrophomonas)、鞘氨醇杆菌属(Sphingobacterium)、微杆菌属(Microbacterium)、热单胞菌属(Thermomonas)、申氏杆菌属(Shinella)、纤维弧菌属(Cellvibrio)和链霉菌属(Streptomyces)。由表6可知,桃太郎/新砧处理根系的假单胞菌属、unclassified_f__Burkholderiaceae、波氏杆菌属和unclassified_f__Rhodanobacteraceae在其内生细菌组成结构中占比分别为19.37 %、2.80 %、1.93 %和2.74 %,较新砧处理相应属在其内生细菌组成结构中的占比分别降低16.32、2.57、3.64和1.90个百分点,而黄杆菌属在其内生细菌组成结构中占比为7.04 %,较新砧处理提高2.1个百分点;与桃太郎处理相比,桃太郎/新砧处理根系假单胞菌属在其内生细菌组成结构中占比降低49.80个百分点,而黄杆菌属和unclassified_f__Burkholderiacea在其内生细菌组成结构中占比分别提高5.19和1.41个百分点。从表6还可看出,桃太郎/宝砧处理根系的假单胞菌属和unclassified_f__Rhodanobacteraceae在其内生细菌组成结构中占比分别为12.22 %和3.34 %,分别较宝砧处理降低33.53和0.97个百分点,而黄杆菌属、unclassified_f__Burkholderiaceae、波氏杆菌属和纤维弧菌属在其内生细菌组成结构中占比分别为2.48 %、5.51 %、7.97 %和7.68 %,较宝砧处理分别提高0.26、2.69、5.84和6.48个百分点;与桃太郎处理相比,桃太郎/宝砧处理根系的假单胞菌属在其内生细菌组成结构中占比降低56.95个百分点,而黄杆菌属和unclassified_f__Burkholderiaceae在其内生细菌组成结构中占比分别提高0.63和4.12个百分点。同时,桃太郎/新砧和桃太郎/宝砧处理根系均富集了鞘氨醇杆菌属、Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium和纤维弧菌属细菌。可见,嫁接番茄植株根系优势内生细菌在属分类水平组成的种类均多于相应的砧木和接穗实生自根植株,这也可能是嫁接番茄植株在正常情况下抗性得以增强的原因之一。
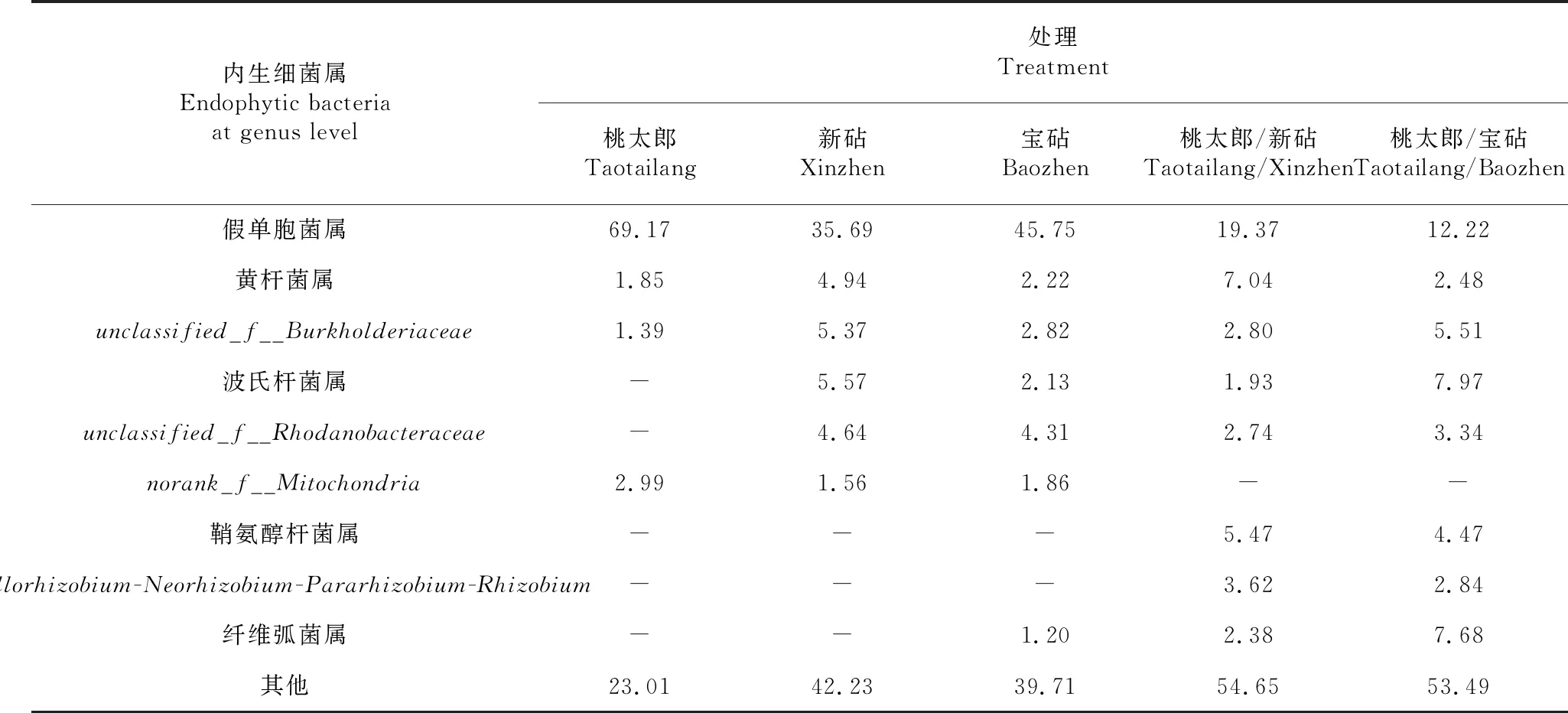
表6 各品种番茄及其嫁接植株根系内生细菌属分类水平占比比较
对比图4各处理番茄植株根系内生真菌属分类水平群落的相对丰度可知,各处理根系内生真菌的优势菌属主要有油壶菌属(Olpidium)、Cutaneotrichosporon、球囊菌属(Plectosphaerella)、红酵母菌属(Rhodotorula)、枝孢菌属(Cladosporium)、镰刀菌属(Fusarium)、木霉菌属(Trichoderma)、杯梗孢属(Cyphellophora)、柄孢壳属(Zopfiella)、微座孢属(Microdochium)、弯孢属(Curvularia)和链格孢属(Alternaria)等。由表7可知,桃太郎/新砧处理根系的油壶菌属和unclassified_f__Chaetomiaceae在其内生真菌优势菌属中占比分别为3.65 %和1.55 %,分别较新砧处理降低43.50和1.35个百分点,而Cutaneotrichosporon、球囊菌属、红酵母菌属、木霉菌属和unclassified_k__Fungi在其内生真菌优势菌属中占比分别为12.26 %、29.20 %、29.10 %、3.65 %和7.87 %,分别较新砧处理提高6.36、26.12、28.0、2.65和6.65个百分点;同时,枝孢菌属和unclassified_o__Branch06真菌富集,unclassified_f__Didymellaceae、镰刀菌属、unclassified_f__Nectriaceae和unclassified_c__Sordariomycetes真菌缺失;与桃太郎处理相比,桃太郎/新砧处理根系的油壶菌属、Cutaneotrichosporon、枝孢菌属、木霉菌属和unclassified_f__Chaetomiaceae在其内生真菌优势菌属中占比分别降低3.49、7.68、4.34和3.64个百分点,但球囊菌属、红酵母菌属和unclassified_k__Fungi在其内生真菌优势菌属中占比分别提高11.75、27.71和6.09个百分点;同时,unclassified_o__Branch06真菌富集、unclassified_f__Nectriaceae真菌缺失。从表7还可看出,桃太郎/宝砧处理根系的油壶菌属、Cutaneotrichosporon、枝孢菌属和镰刀菌属在其内生真菌优势菌属中占比分别为36.03 %、5.17 %、2.37 %和8.41 %,分别较宝砧处理降低2.13、3.38、17.17和10.65个百分点,而球囊菌属、红酵母菌属、unclassified_k__Fungi、unclassified_f__Nectriaceae和青霉菌属在其内生真菌优势菌属中占比分别为2.28 %、12.39 %、4.56 %、2.60 %和4.60 %,分别较宝砧处理提高0.17、11.31、1.8、0.38和3.59个百分点;此外,unclassified_f__Didymellaceae属真菌富集;与桃太郎处理相比,桃太郎/宝砧处理根系的Cutaneotrichosporon、球囊菌属和枝孢菌属在其内生真菌优势菌属中占比分别分别降低30.29、15.17和11.06个百分点,而油壶菌属、红酵母菌属、unclassified_k__Fungi和unclassified_f__Nectriaceae在其内生真菌优势菌属中占比分别提高28.89 %、11.00 %、2.78 %和1.14 %;同时,unclassified_f__Didymellaceae、镰刀菌属和青霉菌属真菌富集,木霉菌属和unclassified_f__Chaetomiaceae真菌缺失。综上所述,与砧木和接穗实生自根植株相比,嫁接番茄植株根系内生真菌优势属的占比虽然发生了一定变化,但嫁接植株根系优势内生真菌属的数量与砧木或接穗实生自根番茄植株基本一致,未呈现如内生细菌一样增加的变化趋势。由此推测,嫁接番茄植株根系的内生真菌可能比内生细菌更难定殖。
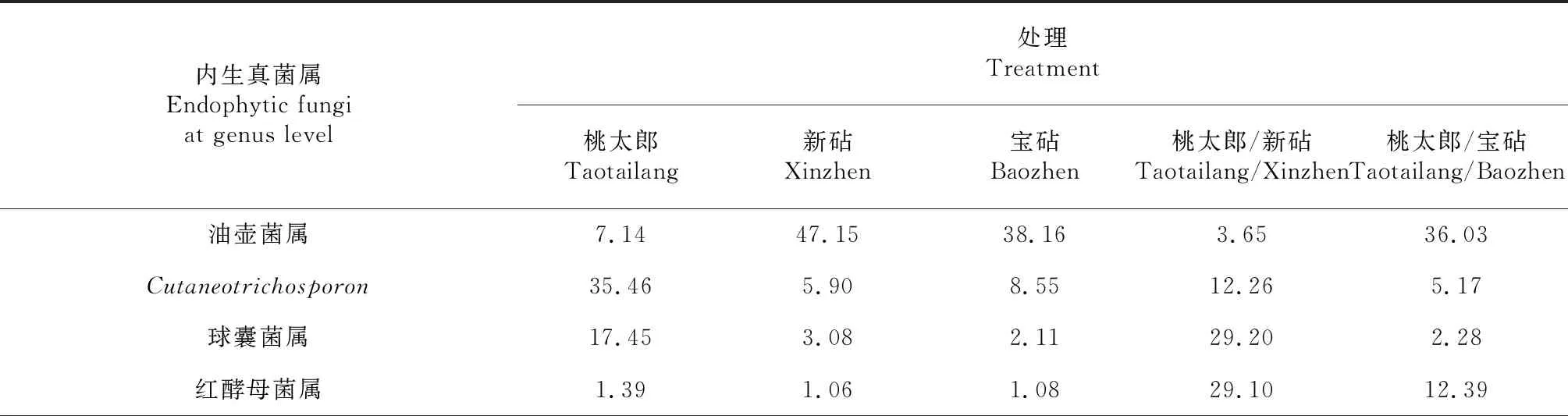
表7 各品种番茄及其嫁接植株根系内生真菌属分类水平的占比比较
2.4 番茄嫁接和自根植株根系内生细菌和真菌基于OTU水平的Venn图分析
基于OTU水平的Venn图(图5)显示,桃太郎、新砧和桃太郎/新砧处理根系的共有优势细菌OTU为362个,新砧处理特有的OTU为90个,桃太郎处理特有的OTU为81个,桃太郎/新砧处理特有的OTU为43个;桃太郎、宝砧和桃太郎/宝砧处理根系的共有优势细菌OTU为409个,宝砧处理特有的OTU为55个,桃太郎处理特有的OTU为58个,桃太郎/宝砧处理特有的OTU为43个。从图6可看出,桃太郎、新砧和桃太郎/新砧处理根系的共有优势真菌OTU为107个,新砧处理特有的OTU为110个,桃太郎处理特有的OTU为103个,桃太郎/新砧处理特有的OTU为31个;桃太郎、宝砧和桃太郎/宝砧处理根系的共有优势真菌OTU为118个,宝砧处理特有的OTU为64个,桃太郎处理特有的OTU为99个,桃太郎/宝砧处理特有的OTU为90个。可见,嫁接番茄植株根系中特有的OTU可能是其抗性增强的原因之一,同时,嫁接植株根系内生细菌和真菌的种类受砧木种类和接穗共同影响。
3 讨 论
根系健康的微生物一方面可抑制植物病原菌过度繁殖,另一方面可酶解产生诱导物质并积累大量次生抗病物质,提高作物的抗病性[17]。植物根系的内生细菌在植物生长发育、病虫害生防及生物修复等方面发挥着重要作用,其多样性是植物内生菌与植物间微生态关系平衡的重要保障[18]。植物根系内生真菌能与宿主植物形成互利共生关系,产生次生代谢物质提高宿主植株的抗逆性[19]。对于嫁接植株,砧木的基因型直接影响接穗对营养物质的吸收和转化[20]。因此,从内生微生物角度探究嫁接对番茄植株抗性的影响,对番茄生产上筛选和开发利用有益微生物具有重要的指导作用。
任奎瑜等[21]研究认为,Ace指数和Chao1指数越大,说明物种的丰富度越高;Shannon指数越大,Simpson指数越小,说明物种的多样性越高;Heip指数越大,说明物种的均匀度越高。本研究中,嫁接番茄植株根系内生细菌的Shannon指数显著大于其桃太郎接穗实生自根植株,说明嫁接有助于提高番茄植株根系内生细菌的多样性。已有研究表明,植株根系的内生细菌通过产生抗生素类物质、水解酶、生长调节剂和诱导植物产生抗性等途径控制植物病害[22]。由此推测,多样性更丰富的根系内生细菌会促进番茄植株分泌抗生素类物质,诱导植物产生抗性,从而提高番茄嫁接植株的抗性。本研究还发现,嫁接番茄植株根系内生细菌的Heip指数均显著大于其桃太郎接穗实生自根植株,明显大于相应的砧木实生自根植株,说明嫁接番茄植株根系内生细菌的均匀度较高,群落结构更均匀;番茄嫁接植株根系的内生真菌种类及其多样性不仅受砧木品种影响,还受接穗类型影响,其中,与相应的新砧和宝砧处理相比,桃太郎/新砧处理根系内生真菌的多样性和丰富度呈下降趋势,而桃太郎/宝砧处理根系内生真菌的多样性和丰富度呈增加趋势,与文才艺[23]等的研究结果一致,可能与不同品种番茄植株内生真菌占据不同生态位、植物体内或表现为分离频率高、数量大的优势种群,或为一些稀有种群紧密相关。
本研究基于门分类水平分析发现,变形菌门、放线菌门、拟杆菌门和厚壁菌门细菌虽然都是番茄接穗、砧木和嫁接植株根系的优势内生细菌门,但嫁接后番茄植株根系内生放线菌门和拟杆菌门细菌在其内生细菌组成结构中占比增加,变形菌门细菌在其内生细菌组成结构中占比降低,与Mendes等[24]研究认为放线菌门的相对丰度与抗病能力呈正相关、对植物病害具有防御作用的观点一致,说明嫁接能增加有益细菌门的数量,从而提高植株抗性;子囊菌门、油壶菌门、unclassified_k__Fungi和担子菌门真菌都是番茄接穗、砧木和嫁接植株根系的优势内生真菌门,但番茄嫁接植株子囊菌门在其内生真菌组成结构中的占比低于其接穗实生自根植株,其中,寄生型子囊菌门可引起植物病害[25],说明嫁接会降低有害真菌门的数量,这可能是嫁接可提高番茄植株抗性的原因之一。
本研究基于属分类水平分析发现,假单胞菌属、黄杆菌属、波氏杆菌属、unclassified_f__Burkholderiaceae、不粘柄菌属、分支杆菌属、寡养单胞菌属、鞘氨醇杆菌属、微杆菌属、热单胞菌属、申氏杆菌属、纤维弧菌属和链霉菌属细菌都是番茄砧木、接穗和嫁接植株根系的优势内生细菌属。桃太郎/新砧和桃太郎/宝砧处理植株根系假单胞菌属在其内生细菌属组成结构中占比低于相应的砧木和接穗实生自根植株,假单胞菌属细菌作为植物病原菌[26],会危害番茄、黄瓜和甜菜等作物生长,说明嫁接可降低番茄植株根系病原菌数量;同时,嫁接番茄植株根系均富集了具有解磷功能的鞘氨醇杆菌属和固氮功能的Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium[27],说明嫁接能富集对番茄植株根系生态循环有益的微生物;与砧木和接穗实生自根植株相比,嫁接番茄植株根系具有更丰富的功能内生细菌,同时可降低致病内生细菌数量,这可能是嫁接番茄植株抗性得以提高的主要原因之一。
本研究中,油壶菌属、Cutaneotrichosporon、球囊菌属、红酵母菌属、枝孢菌属、镰刀菌属、木霉菌属、杯梗孢属、柄孢壳属、微座孢属、弯孢属和链格孢属真菌都是番茄接穗、砧木和嫁接植株根系的优势内生真菌属;与砧木和接穗实生自根植株相比,番茄嫁接植株根系中优势内生真菌属在其内生真菌优势属组成结构中占比虽然发生了一定的变化,但番茄嫁接植株根系优势内生真菌属种类的数量基本与砧木或接穗实生自根植株一致;嫁接番茄植株根系的油壶菌属在其内生真菌优势属组成结构中占比低于相应的砧木实生自根植株,而油壶菌属是一种在植株根系转性寄生的传毒真菌[28],说明嫁接番茄植株根系的有害真菌数量较少;嫁接番茄植株根系红酵母菌属占比显著大于接穗和砧木实生自根植株,而红酵母菌属中的微生物具有促进宿主对矿物质和有机质养分的吸收[29],提高宿主植物抗逆性[30]的作用。基于OTU水平的Venn分析可知,嫁接番茄植株根系特有的OTU可能是其抗性增强的原因之一。
综上所述,番茄嫁接可改变植株根系内生微生物的多样性特征,尤其能提高根系内生细菌的多样性,改善根系内生细菌的均匀结构;在门分类水平,嫁接番茄植株根系内生细菌具有抗病功能的放线菌门占比增加,可引起植物病害的子囊菌门占比降低;在属分类水平,嫁接番茄植株根系内生细菌病原菌假单胞菌属和传毒真菌油壶菌属的占比降低;嫁接番茄植株根系除拥有与砧木和接穗实生自根植株相同的优势内生细菌属外,还富集了具有诱导植物产生抗性物质的优势内生细菌属,以及根据不同砧木种类富集或缺失部分优势真菌属。目前人们对内生真菌与病原真菌及内生真菌与宿主植物之间的关系仍知之甚少[31],因此,番茄嫁接植株根系富集与缺失的优势内生真菌属功能仍有待深入探究。
4 结 论
番茄嫁接植株根系具有较其砧木和接穗实生自根植株根系更丰富的内生细菌多样性和更均匀的群落结构;在门和属分类水平,番茄嫁接植株根系的内生细菌和真菌数量增加,病原微生物数量减少,并富集有解磷和固氮等促生功能的有益内生菌属,拥有更丰富的微生物OTU和特有OTU,是番茄嫁接植株抗性增强的重要原因之一。
