甘南草原3种啮齿动物的食性及其营养生态位特征
2021-08-03张彩军王小燕姚宝辉康宇坤苏军虎
张彩军, 王小燕, 姚宝辉, 王 缠, 康宇坤, 张 倩, 苏军虎
(甘肃农业大学草业学院/草业生态系统教育部重点实验室/甘肃省草业工程实验室/中美草地畜牧业可持续发展研究中心, 甘肃农业大学-新西兰梅西大学草地生物多样性研究中心, 国家林业草原高寒草地鼠害防控工程技术研究中心, 甘肃 兰州 730070)
生态位理论是生态学研究的基础理论,在阐述物种共存和生物群落构建方面有着重要意义。国内外学者对于生态位理论的研究主要集中在时间生态位[1]、空间生态位[2]和营养生态位[3]等方面。其中,营养生态位对于生态系统的研究至关重要,营养生态位是指动物对食物资源的利用,一方面表现其自身的营养需求特征,另一方面反映出它在生态系统中所处的营养层和功能[4]。但动物的营养生态位会随着所处环境的改变而改变,栖息地、食物资源的分布和质量以及种内和种间的竞争等因素都会影响动物的食性,进而导致其营养生态位发生变化。营养生态位研究的关键在于动物的食性分析,因为食性是动物根据自身生长发育的需求在长期的进化中对所处环境适应的结果[5]。分析动物的食性及其营养生态位特征对于认识动物与环境的相互关系有着重要意义。
啮齿动物是哺乳动物中最多的一个类群,在草地生态系统中有着重要的地位和作用,其群落结构随着草地生态系统的变化而演替[6]。啮齿动物对于栖息地环境的变化极为敏感,退化的草地为啮齿类提供了适宜的生境,导致其大量繁殖,加速草地退化[7]。然而草地植物群落的逆向演替,又会影响啮齿动物间的食物和空间资源竞争,亦会改变啮齿动物的群落组成[8]。钟文勤[9]发现随着草地的退化,啮齿动物优势种由达乌尔鼠兔(Ochotonadauurica)更替到布氏田鼠(Lasiopodomysbrandtii)再到长爪沙鼠(Merionesunguiculatus)。退化的草地经人工修复(补播)后,高原鼠兔的种群数量显著下降[10]。草地植被组成的改变会使啮齿动物喜食植物减少,而植被盖度和高度的增加,会影响啮齿动物的视野和活动等,增加了被捕食风险,导致其成为啮齿动物不适宜的生境[11]。通过认识环境变化下啮齿动物食性特征,以及种间相互关系和演替过程,对退化草地的合理修复和优化草地管理策略具有重要意义。
甘南草原位于青藏高原东北部边缘,有“亚洲第一牧场”和“中华水塔”之美誉,对当地的畜牧业发展和我国水源涵养与生态安全方面有着重要意义[12]。由于长期过度放牧和缺乏科学的管理等导致草地退化严重,啮齿动物种群数量激增[13]。有报道甘南草原发生鼠害面积占可利用草原面积的33.7%,特别是位于甘南西部、南部和西南部的亚高山草甸,是高原鼠兔(O.curzoniae)、高原鼢鼠(Eospalaxbaileyi)和喜马拉雅旱獭(Marmotahimalayana)危害亚区,也是分布最集中、危害最严重的区域,危害面积73.44 万hm2,占危害总面积的80%,严重影响了当地的生态安全和可持续发展[14]。实践中发现,随着高原鼢鼠种群数量的上升,高原鼠兔开始入侵高原鼢鼠栖息地,破坏并侵占高原鼢鼠洞道,迫使高原鼢鼠转移迁徙,直至共栖地的高原鼢鼠消失。其后高原鼠兔种群数量爆发[15]。研究啮齿动物的种间关系,对于认识啮齿动物种群动态以及其对草地群落演替的影响有着重要意义。特别是在当前生态修复工程的大量实施背景下,如何兼顾草地功能提升,避免鼠害的二次发生,实现工程的可持续性等,具有重要的现实指导意义。
本研究通过对甘南草原3种啮齿动物粪便和胃容物的显微组织观察,分析3种啮齿动物的食物组成及其比例,从食性角度探讨高原鼠兔、喜马拉雅旱獭和高原鼢鼠的生态位重叠情况,进一步探讨3者的种间竞争、共存机制,为优化草地管理策略提供参考。
1 材料和方法
1.1 试验区概况
试验区位于甘肃省甘南藏族自治州碌曲县尕海乡,地理坐标为北纬34°21′27.48″~34°21′38.09″,东经102°17′07.16″~102°17′41.56″。平均海拔3 500 m,境内年平均温度2.6℃,全年无霜期56 d。年降水量633~782 mm。土壤以亚高山草甸土和高山草甸土为主。植被类型为高寒灌丛草甸和高寒草甸,分别占全县草地面积88.68%,11.32%[16]。啮齿动物种数占甘南啮齿动物种数的74.51%,高原鼠兔和高原鼢鼠是该县啮齿动物的绝对优势类群,喜马拉雅旱獭常与其同域分布[17]。
1.2 试验方法
1.2.1样品采集 2019年9月,在面积为42 hm2的研究区域内收集高原鼠兔和喜马拉雅旱獭新鲜粪便(两物种粪便形状、大小差异明显),高原鼠兔粪便按其分布由3~5堆粪便组成一个复合分析样,每一堆粪便只取一小部分。喜马拉雅旱獭的粪便则按照不同便厕、不同洞群收集,以保证所收集样本为不同个体粪便。共收集高原鼠兔粪便复合样50份,喜马拉雅旱獭粪便21份。由于高原鼢鼠终年地下生活,且洞道结构复杂,很难获得粪便样本,故用弓形夹夹捕高原鼢鼠,捕获后解剖,采集高原鼢鼠胃及其内容物,共采集到17份(雌性10只、雄性7只)。采集研究区内出现的所有植物物种,每种植物采集两份(完整植株),一份置于标本夹中,用于鉴定植物种属。另一份置于牛皮纸信封中,带回实验室用于制作显微鉴定的植物对照样本。
1.2.2样片的制备和镜检 参考相关文献[18],将牛皮纸信封中用于制作显微鉴定的植物对照样本的植物,分根、茎和叶3个部位进行粉碎、研磨,研磨后依次经60目和100目网筛筛选,留取100目筛上物作为试验样品。取适量样品置于试管中,加入5~7 ml 10% HNO3,在酒精灯上加热3 min,模拟动物消化过程。之后在200目网筛中进行冲洗。取适量样品于载玻片上,滴加少量甘油,使植物碎片均匀分布,盖上盖玻片,并用中性树脂封片。在100 X显微镜下观察并拍照保存,作为显微鉴定的对照样本。
粪便的烘干、研磨和过筛与植物对照样片制作的操作步骤相同。过筛后取筛上物0.5 g置于培养皿中,加入10% NaClO溶液,8 h后在200目网筛中进行冲洗,筛上物作为试验样品。将收集的胃内容物放入试管中,加入5% H2GrO4浸泡24 h后经200目的网筛冲洗,筛上物移入烧杯中后加入10% NaClO溶液,漂白10 min后冲洗,筛上物作为试验样品[19]。
装片方法参照曹伊凡和苏建平[20]的方法,每份样品制作5张装片。参照沈广爽[21]镜检方法:每张装片在100 X的显微镜下检查20个视野,每份样品共计检查100个视野。观察每个视野中出现的可辨认植物表皮角质碎片,根据不同的细胞形态类别和特点,对比已有的植物显微装片鉴定到属种,并记录植物在视野中的出现频次。
1.3 数据分析与统计
统计所得的各植物角质碎片的出现频次(F),依据公式:F=100×(1-e-D)[22],将其转换为每个视野中各植物可辨认碎片的平均密度(D),即:D=ln(1-100/F)。根据:相对密度(RD)=各植物可辨认碎片平均密度/各植物可辨认碎片平均密度之和×100%,将D转换为RD[22]。RD即为该种植物占食物组成的百分比,并根据RD值排列各物种可食植物的取食序位。
参考相关文献[23],通过香农-维纳(Shannon-Weiner)多样性指数(H)、最大香农-维纳多样性指数(Hmax)、皮洛(Pielou)均匀度指数(J)和食物生态位宽度指数(B)分析3种啮齿动物食物组成的多样性:
Hmax=lnS
式中:Pi为食物i在取食者食物中出现的频率,S代表粪样中存在的植物物种数。
通过Schoener食物重叠度(Cij)分析3种啮齿动物的食物重叠度。
Cij=1-0.5×∑(|Pik-Pjk|)
Pik,Pjk分别为不同啮齿动物共同采食植物在各自取食植物中所占的百分比。
采用SPSS 23.0软件对3种啮齿动物食谱中不同类群植物相对密度进行单因素方差分析,所有数据处理在Excel 2010中完成。
2 结果与分析
2.1 3种啮齿动物食物组成及比例
3种啮齿动物粪便和胃容物的显微组织的观察结果显示(表1),各物种所采食的植物种类分别为:高原鼢鼠采食14科26种,喜马拉雅旱獭14科27种,高原鼠兔14科25种。3种啮齿动物的所采食的植物种类大致相同,但采食同种植物的比例不同,高原鼢鼠采食的植物主要有珠芽蓼(Polygonumviviparum,28.69%)、鹅绒委陵菜(Potentillaanserina,19.64%)和草地早熟禾(Poapratensis,13.84%)。按科统计,蓼科(Polygonaceae)、禾本科(Poaceae),蔷薇科(Rosaceae)和菊科(Asteraceae)植物为高原鼢鼠的主要食物来源,分别占其食谱的28.69%,21.03%,20.56%和15.64%。喜马拉雅旱獭和高原鼠兔所采食的主要植物均为禾本科、菊科和玄参科(Mazaceae)植物,其中,喜马拉雅旱獭采食3科植物的比例分别为28.69%,17.16%和16.02%,高原鼠兔采食比例分别为74.98%,6.27%和5.26%。喜马拉雅旱獭所采食的植物主要为垂穗披碱草(Elymusnutans,24.59%),兰石草(Lanceatibetica,16.02%)和白苞筋骨草(Ajugalupulina,14.21%)。禾本科的垂穗披碱草和草地早熟禾作为高原鼠兔主要的食物来源,分别占其食谱的57.30%,17.68%。按植物功能群统计,高原鼢鼠对于莎草科(Cyperaceae)植物的采食数量显著高于其它两种动物(P< 0.05),而高原鼠兔对禾本科植物的采食数量显著高于其它两啮齿动物(P< 0.05),高原鼢鼠和喜马拉雅旱獭采食的杂类草显著高于高原鼠兔所采食的量(P< 0.05,图1)。
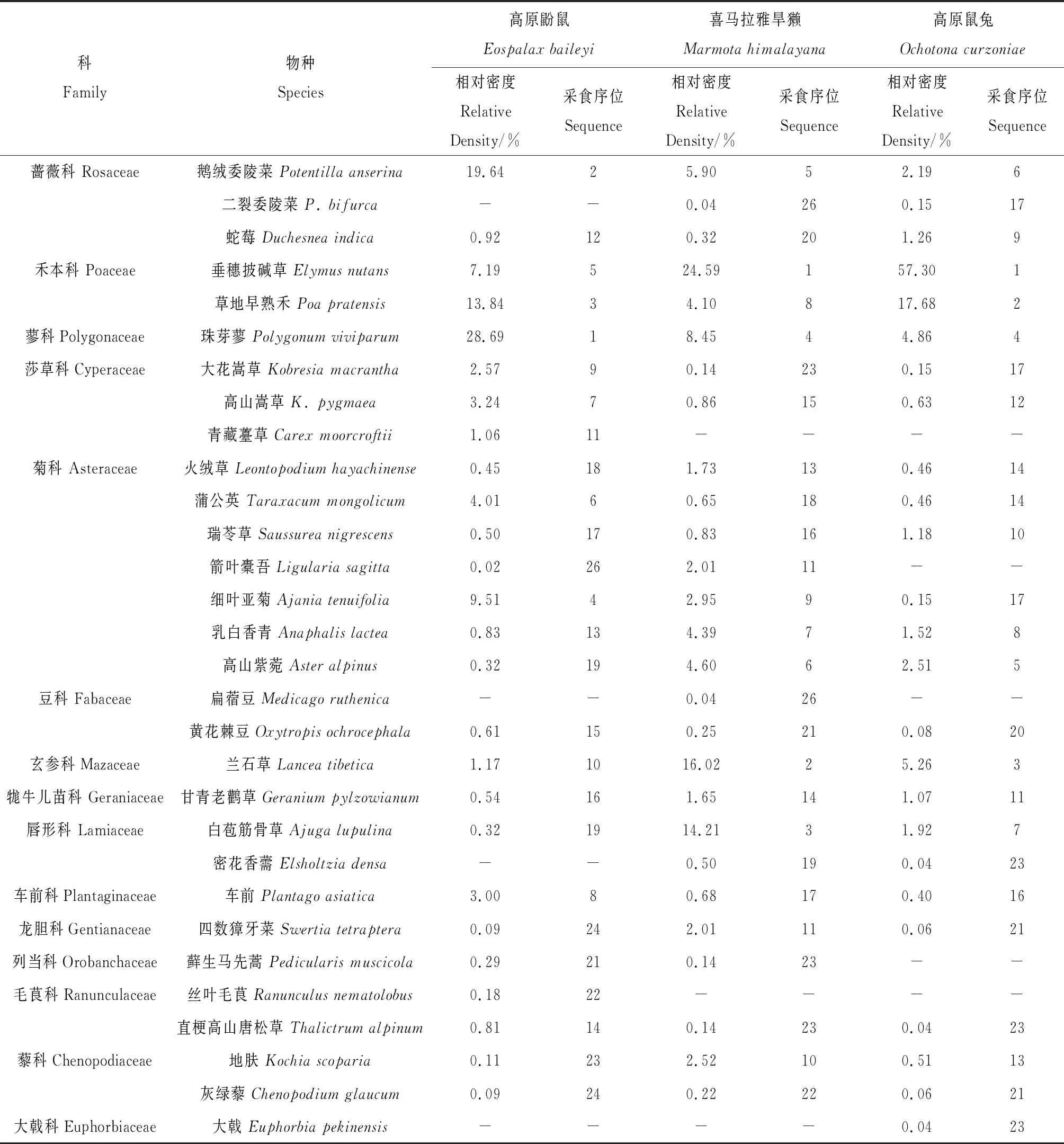
表1 3种啮齿动物的食物组成及比例
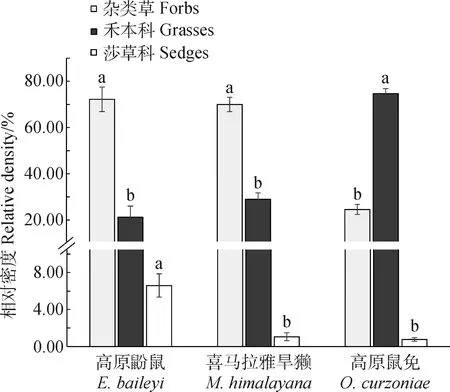
图1 3种啮齿动物食物主要类群
2.2 食物多样性及营养生态位宽度
分析3种啮齿动物食物多样性及营养生态位,结果表明(表2),香农-维纳多样性指数H,皮洛均匀度指数J的值的大小顺序均为:喜马拉雅旱獭>高原鼢鼠>高原鼠兔。喜马拉雅旱獭的食物生态位宽度最高(7.95),高原鼢鼠次之(6.29),高原鼠兔的食物生态位宽度远小于两者,仅为2.73。
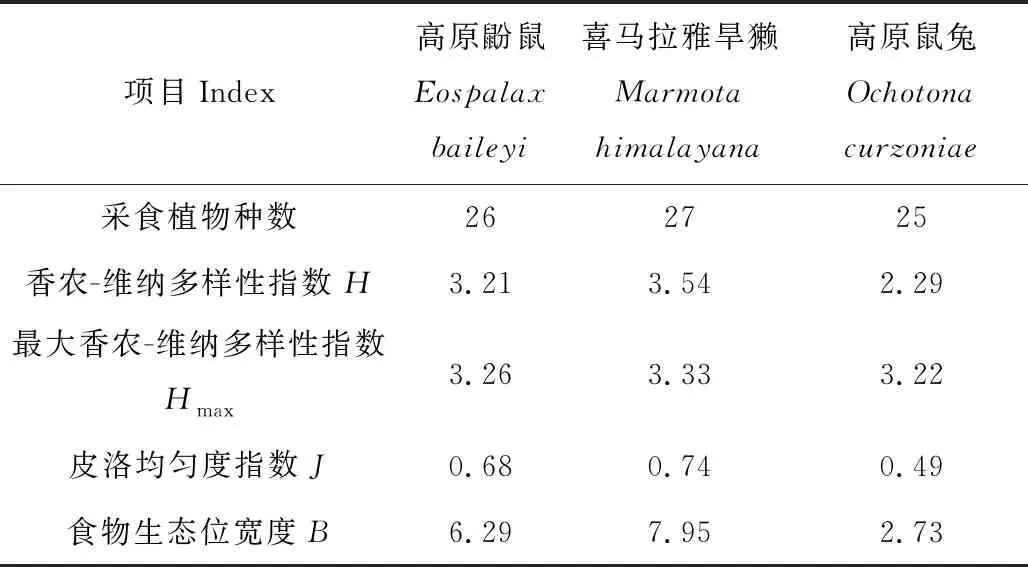
表2 3种啮齿动物的营养生态位参数
2.3 食物重叠度
3种啮齿动物食物重叠指数显示(表3),各物种间均存在营养生态位重叠。其中,高原鼠兔和喜马拉雅旱獭的食物重叠指数最高(0.52)。高原鼢鼠与喜马拉雅旱獭食物重叠指数为0.36,高原鼢鼠与高原鼠兔的食物重叠指数为0.35。

表3 3种啮齿动物的食物重叠指数
3 讨论
3.1 3种啮齿动物的食性差异
本研究中3种啮齿动物所采食植物的种类基本相同,但采食同种植物的比例各异。禾本科、菊科和玄参科植物均在高原鼠兔和喜马拉雅旱獭食谱中占前3位。高原鼠兔和喜马拉雅旱獭日常采食活动在地面上进行,相同的取食方式可能导致二者在食物资源利用方面有着相似之处[23]。热娜古丽·艾合麦提在阿尔金山的相关研究结果表明,高原鼠兔和喜马拉雅旱獭在暖季采食的植物都主要为禾本科、莎草科、豆科(Fabaceae)和菊科[24]。这与我们的研究结果也相似。而对高原鼠兔的研究发现,其食物组成随生境的不同而发生变化。在青藏高原高寒草甸高原鼠兔主要以垂穗披碱草、甘肃棘豆(Oxytropiskansuensis)和铺散亚菊(Ajaniakhartensis)等为食[25],在退化草原上,高原鼠兔主要以紫花针茅(Stipapurpurea)、早熟禾(Poasp.)和甘肃棘豆等为食[26],而在高山蒿草草甸和高山灌丛草甸,高原鼠兔则在秋季贮存细叶亚菊(A.tenuifolia)、甘肃马先蒿(Pediculariskansuensis)等植物[27]。本研究中高原鼠兔采食的是垂穗披碱草、草地早熟禾和兰石草等,与高寒草甸的结果相似。与高原鼠兔和喜马拉雅旱獭的食谱不同,高原鼢鼠所采食的植物以杂类草为主,特别是蓼科和蔷薇科植物,这与已有的研究结果相似[28-29]。有研究表明杂类草生物量与高原鼢鼠的种群密度成正比[30]。高原鼢鼠营地下生活,通过挖掘洞道来获取食物,此过程需要消耗大量能量,因此在觅食过程中需要权衡能量代价和收益[31]。杂类草通常具有膨大根茎,其营养也较为丰富,能为高原鼢鼠提供充足的能量。
3.2 3种啮齿动物营养生态位特征
本研究结果表明,高原鼠兔营养生态位宽度最窄(2.73),仅垂穗披碱草和草地早熟禾就占其食谱的74.98%。主要原因是动物的营养生态位宽度会随环境变化而变化,当食物资源充足时,动物利用最适于自身生存和最方便利用的食物资源,导致营养生态位变窄;当食物资源短缺时,食草动物则会扩大其食谱以满足所需,从而导致营养生态位变宽[32]。对南非隐鼠(Cryptomyshottentotus)在不同环境下的研究表明,其在湿度适宜且食物资源充足地区的生态位宽度显著性低于食物资源贫乏的干旱地区[33]。本研究在9月初进行,此时研究区域内植物资源丰富,不存在缺乏食物资源的情况,且研究发现高原鼠兔喜食垂穗披碱草、棘豆(Oxytropissp.)和早熟禾等植物[34]。由此推断目前此环境对高原鼠兔较为适宜。当喜马拉雅旱獭受到家畜和牧民的干扰时,其用于觅食的时间减少而警戒时间增加[35]。本研究区域临近公路,来往车辆可能会对其产生干扰,从而减少其觅食时间,影响食物的选择。此外,在入蛰前夕,喜马拉雅旱獭体内需储存大量脂肪以备冬眠,但此时其出洞时间却较短,多在10点后才出洞,16点以后回洞[36],这些原因导致喜马拉雅旱獭在食物选择上趋于泛化,因此其食物均匀度指数、营养生态位宽度和香农-维纳指数较大。相比其它2种啮齿动物,其活动范围较广,因此能采食到更多种类的植物。高原鼢鼠的营养生态位宽度介于高原鼠兔和喜马拉雅旱獭之间,虽然高原鼢鼠对食物的选择具有偏好性,但需要考虑采食过程的代价,因此其食物也体现出较高的均匀度指数和香农-维纳指数。
3.3 生态位重叠与种间关系
营养生态位重叠可以有效地反映同域分布物种间对资源利用的竞争情况[34]。本研究结果显示,3个物种间营养生态位均有所重叠,表明它们之间存在食物竞争。高原鼠兔和喜马拉雅旱獭均在地上采食,竞争相对较大,因此营养生态位重叠指数较高(0.52)。由于高原鼢鼠常年营地下生活,与地上活动的啮齿类存在空间上的分化,在食物资源利用上竞争较小,因此高原鼢鼠与高原鼠兔、喜马拉雅旱獭的营养生态位重叠指数较低。生态位竞争理论指出,生态位完全重叠的物种不能共存,共存的物种势必在其他生态位方面分离,同域分布的物种间利用相同的食物资源时,会在其利用上表现出差异,如生境、时间和行为等,这在生态位的格局方面体现出一种相互补偿性[37]。青藏高原东缘同域分布的高原鼠兔和达乌尔鼠兔的生态位重叠程度相当大,二者通过不同的活动节律实现时间生态位上的分离(高原鼠兔白昼活动,达乌尔鼠兔则以晨昏活动为主),从而达到共存的目的。营养生态位重叠大的物种也可以通过栖息地的分化来达到空间生态位分离[26]。王权业等[38]发现,高原鼠兔和高原鼢鼠主要栖息在矮嵩草草甸植物群落,而甘肃鼠兔则主要栖息于金露梅灌丛植物群落,其通过水平空间中栖息生境选择来降低种间竞争。而高原鼠兔和高原鼢鼠则通过对地面和地下垂直生境选择,使之能够共存。对内蒙古典型草原布氏田鼠营养生态位及其种间关系的研究表明,布什田鼠营养生态位窄,对食物的选择性强,当食物资源短缺时会以种群迁移的方式避免与其同域分布的达乌尔黄鼠(Spermophilusdauricus)、黑线毛足鼠(Phodopussungorus)和五趾跳鼠(Allactagasibirica)间的竞争[39]。本研究中高原鼢鼠与高原鼠兔、喜马拉雅旱獭在采食空间上分离,高原鼢鼠的采食活动在地下发生,而高原鼠兔和喜马拉雅旱獭则在地上采食。此外,高原鼢鼠主要采食杂类草,而高原鼠兔主要以禾本科植物为食,喜马拉雅旱獭则对各科植物的采食比较均匀。三者在营养生态位上存在明显的分离,通过营养生态位的分化来实现共存。
4 结论
本研究通过对甘南草原3种啮齿动物粪便和胃容物的显微组织观察,分析其食物组成及其比例,发现3种啮齿动物所采食植物的种类基本相同,但采食同种植物的比例各异。高原鼢鼠所采食的植物以杂类草为主,对珠芽蓼和鹅绒委陵菜有明显偏好。喜马拉雅旱獭主要采食禾本科、菊科和玄参科植物,而高原鼠兔主要采食禾本科植物。从食性角度探讨高原鼠兔、喜马拉雅旱獭和高原鼢鼠的生态位重叠情况及其种间关系,发现3种啮齿间营养生态位均有所重叠,表明它们之间存在食物竞争。高原鼠兔和喜马拉雅旱獭竞争相对较大,营养生态位重叠指数最高。而高原鼢鼠与高原鼠兔、喜马拉雅旱獭的营养生态位重叠指数较低。研究表明同域分布的高原鼢鼠、高原鼠兔和喜马拉雅旱獭之间存在食物竞争,且食物上存在分化,营养生态位有所分离。本研究验证了营养生态位分离是动物共存重要机制。
